Contribution of KCNQ and TREK Channels to the Resting Membrane Potential in Sympathetic Neurons at Physiological Temperature

 , , and
, , and 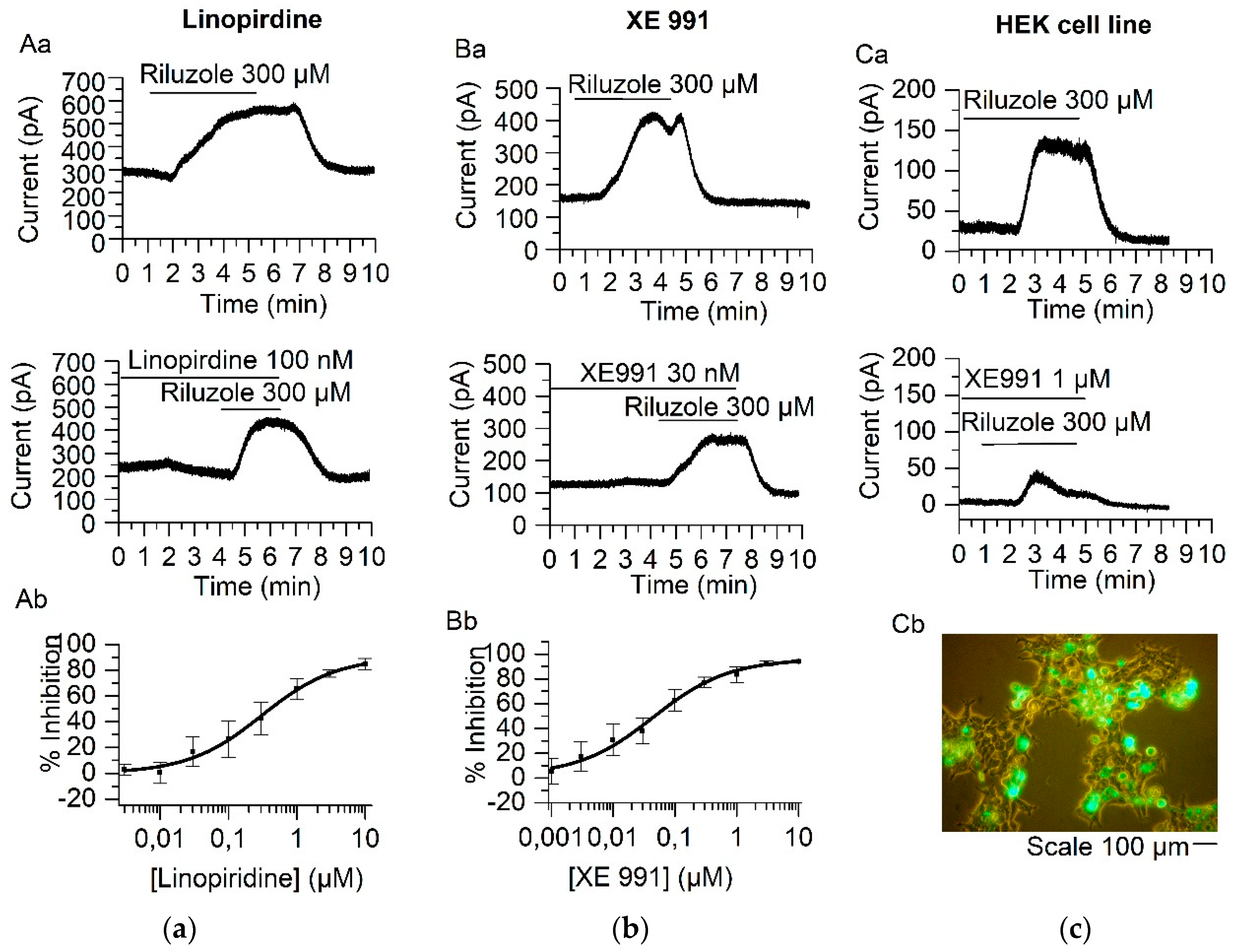{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
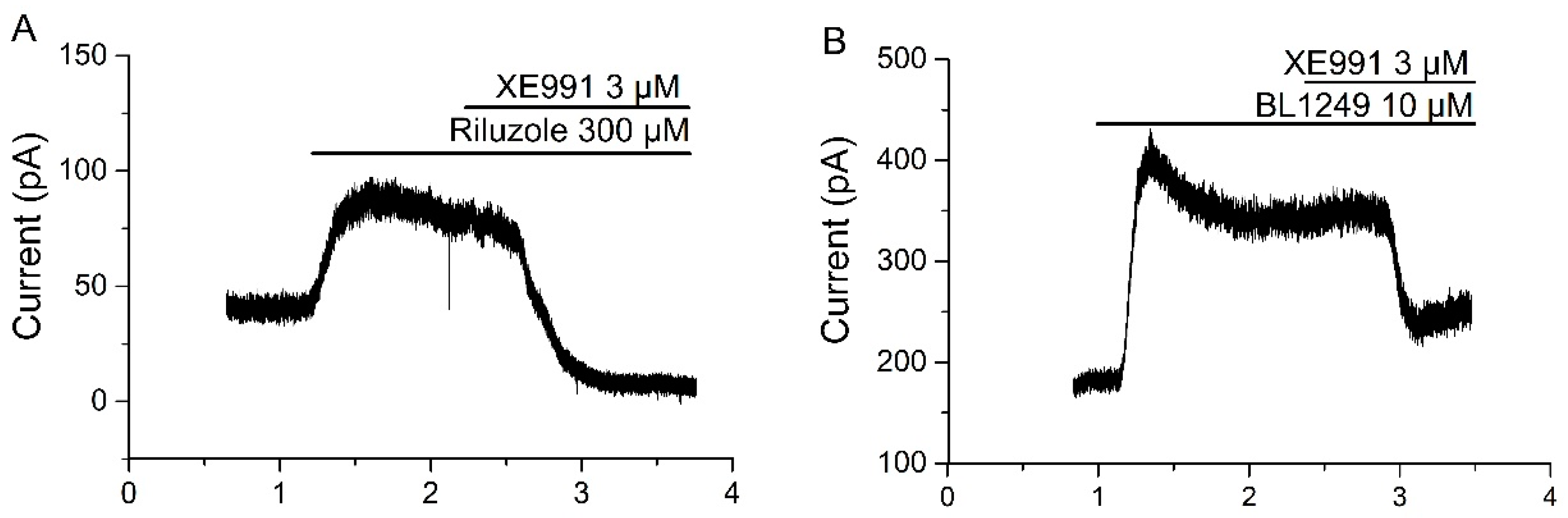
2.1. Linopirdine and Its Analog XE991 Inhibit TREK-2 Current
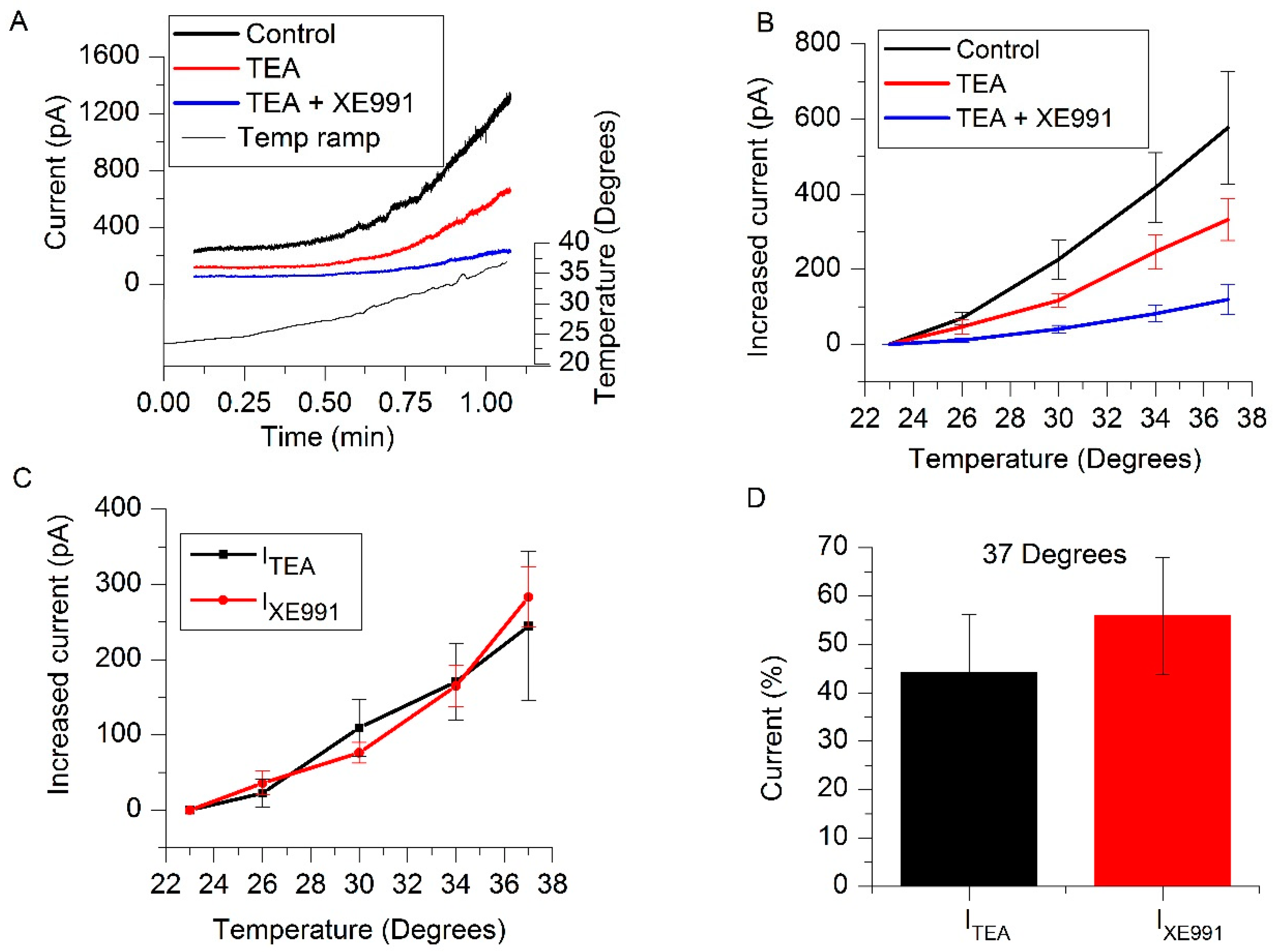
2.2. IM and TREK-2 Currents Are Enhanced by Temperature
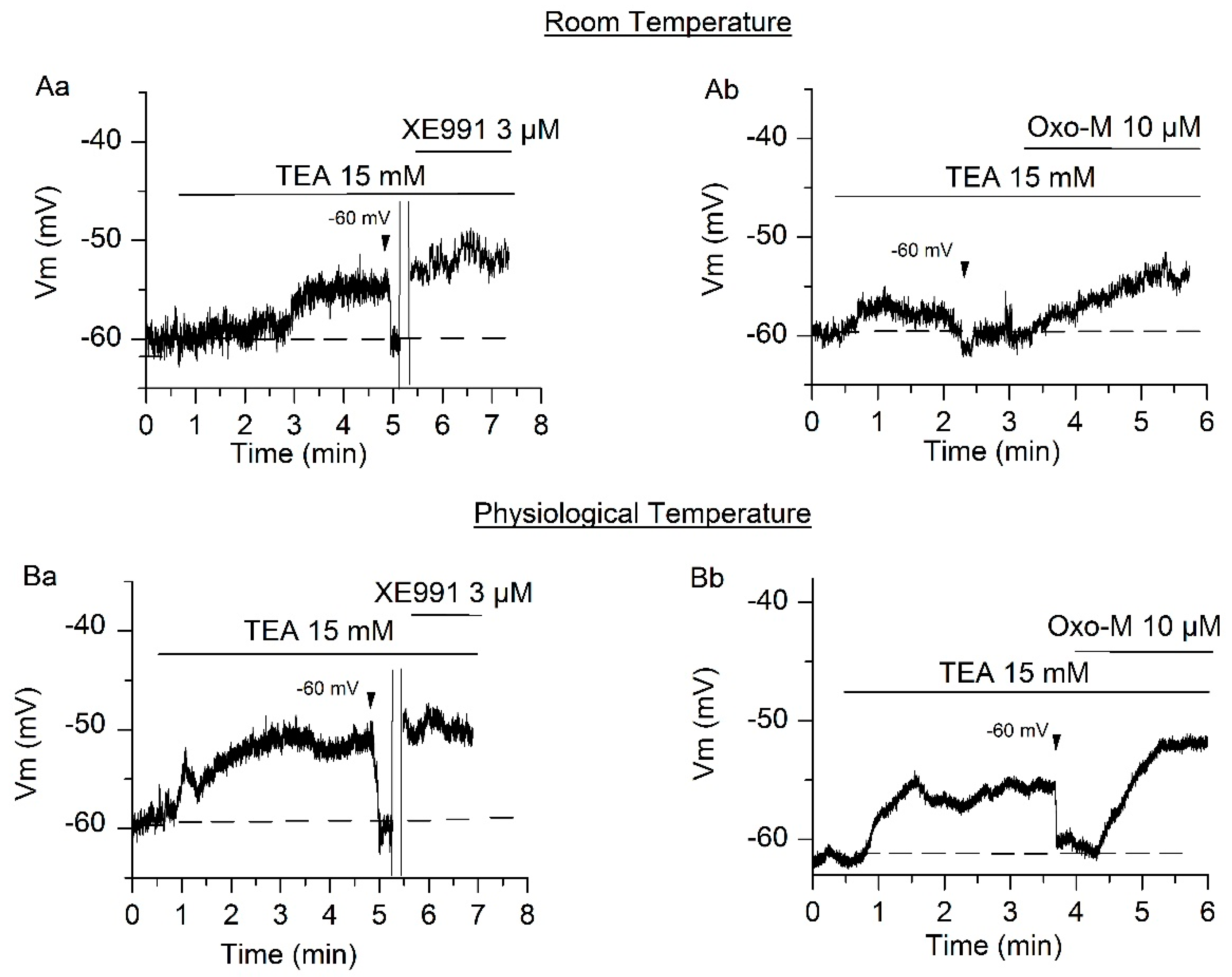
2.3. Contribution of IM and ITREK to the RMP at Physiological Temperature
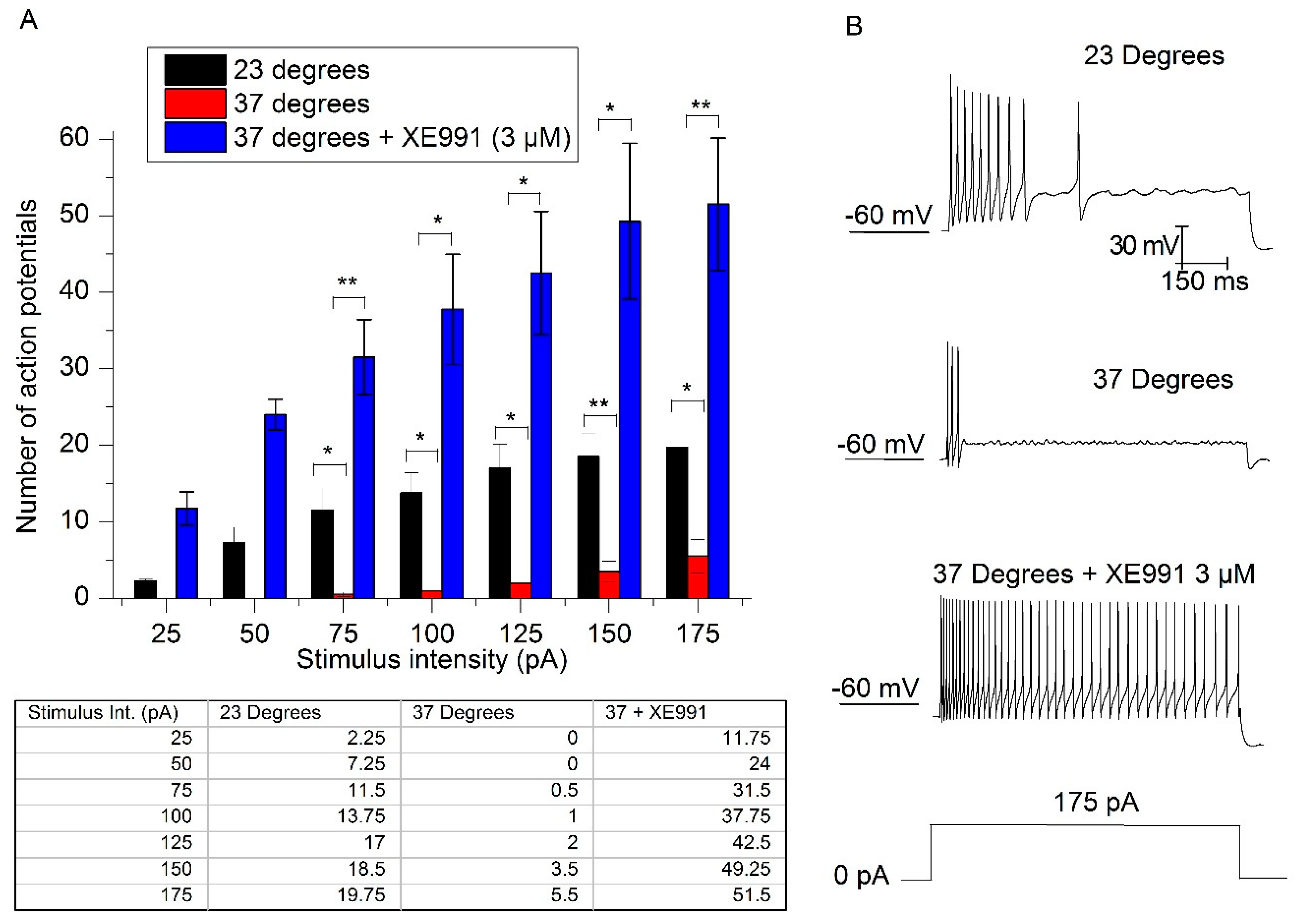
2.4. The Increase in Temperature Enhances Adaptation
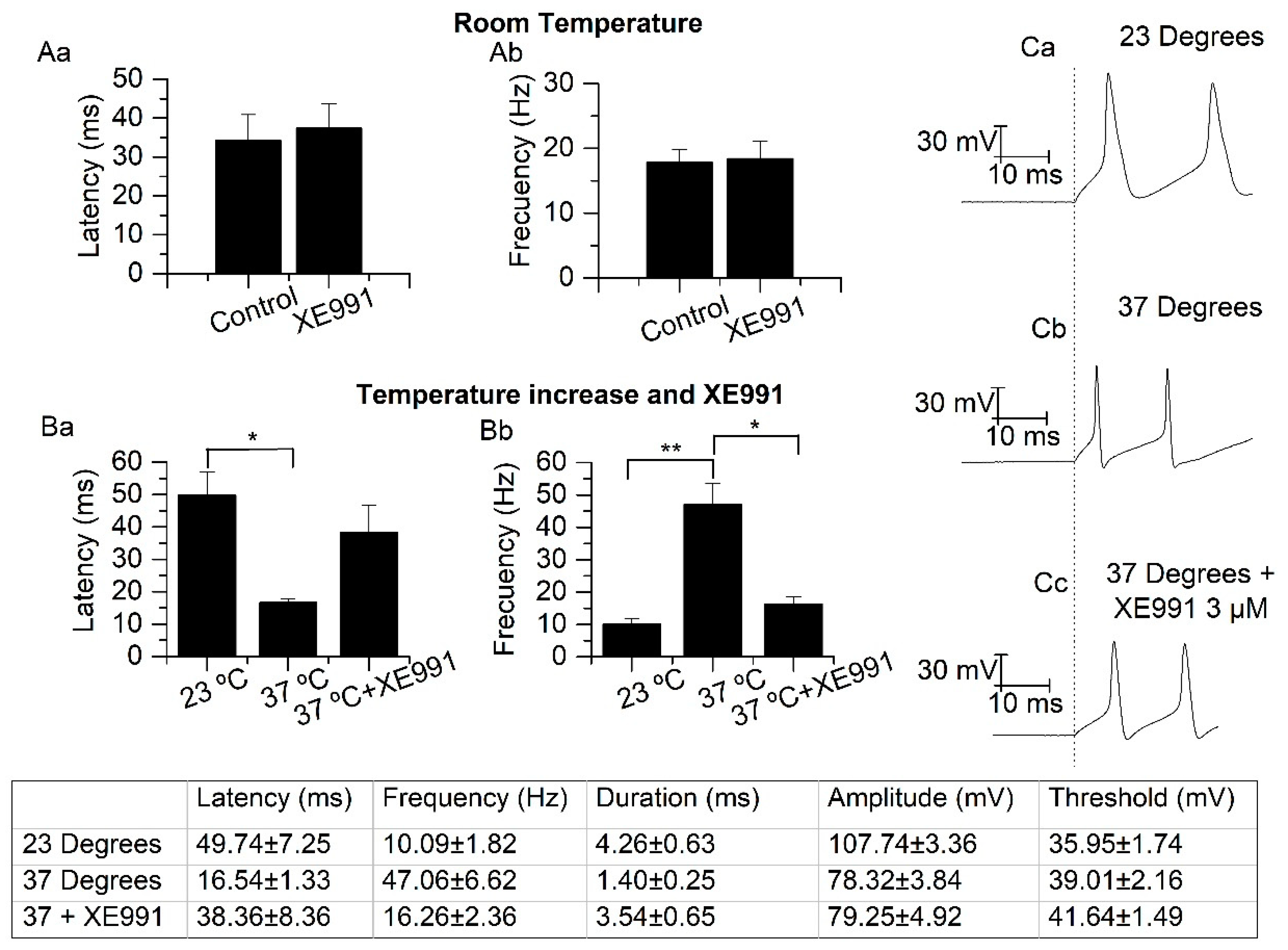
2.5. Temperature Modifies the Action Potential Frequency and Latency
3. Discussion
3.1. Isolation of IM and TREK-2 Current
3.2. Resting Membrane Potential and Excitability
3.3. Physiological Relevance and Conclusion
4. Materials and Methods
4.1. Superior Cervical Ganglion (SCG) Culture
4.2. HEK293 Cell Culture and Transfection
4.3. Perforated Patch Electrophysiology
4.4. Solutions and Drugs
4.5. Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GPCRs | G-protein coupled receptors |
| Ih | h-current |
| INaP | sodium persistent current |
| IM | M-current |
| IRIL | riluzole-activated current |
| ITEA | TEA-inhibited current |
| ITREK | potassium current through TREK channels |
| IXE991 | XE991-inhibited current |
| K2P | two pore domain potassium channels |
| SCG | superior cervical ganglion |
| Oxo-M | oxotremorine-M |
| RMP | resting membrane potential |
| SFA | spike frequency adaptation |
| TALK | TWIK-related alkaline pH-activated K+ channel |
| TASK | TWIK-related acid-sensitive K+ channel |
| TEA | tetraethylammonium |
| THIK | Tandem pore-domain Halothane Inhibited K+ channel |
| TRAAK | TWIK-related arachidonic acid-stimulated K+ channel |
| TREK | TWIK-related K+ channel |
| TRESK | TWIK-related spinal cord K+ channel |
| TTX | tetrodotoxin |
| TWIK | Tandem of pore-domains in a Weakly Inward rectifying K+ channel |
References
- Brown, D.A.; Constanti, A. Intracellular observations on the effects of muscarinic agonists on rat sympathetic neurones. Br. J. Pharmacol. 1980, 70, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.A.; Marrion, N.V.; Smart, T.G. On the transduction mechanism for muscarine-induced inhibition of M-current in cultured rat sympathetic neurones. J. Physiol.-Lond. 1989, 413, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Lamas, J.A. The development of the concept of neuronal resting potential. Fundamental and clinical aspects. Rev. Neurol. 2005, 41, 538–549. [Google Scholar]
- Lamas, J.A.; Reboreda, A.; Codesido, V. Ionic basis of the resting membrane potential in cultured rat sympathetic neurons. Neuroreport 2002, 13, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Lamas, J.A.; Romero, M.; Reboreda, A.; Sanchez, E.; Ribeiro, S.J. A riluzole- and valproate-sensitive persistent sodium current contributes to the resting membrane potential and increases the excitability of sympathetic neurones. Pflugers. Arch. Eur. J. Physiol. 2009, 458, 589–599. [Google Scholar] [CrossRef]
- Cadaveira-Mosquera, A.; Ribeiro, S.J.; Reboreda, A.; Perez, M.; Lamas, J.A. Activation of TREK currents by the neuroprotective agent riluzole in mouse sympathetic neurons. J. Neurosci. 2011, 31, 1375–1385. [Google Scholar] [CrossRef]
- Lesage, F.; Guillemare, E.; Fink, M.; Duprat, F.; Lazdunski, M.; Romey, G.; Barhanin, J. TWIK-1, a ubiquitous human weakly inward rectifying K+ channel with a novel structure. EMBO J. 1996, 15, 1004–1011. [Google Scholar] [CrossRef]
- Cadaveira-Mosquera, A.; Perez, M.; Reboreda, A.; Rivas-Ramirez, P.; Fernandez-Fernandez, D.; Lamas, J.A. Expression of K2P channels in sensory and motor neurons of the autonomic nervous system. J. Mol. Neurosci. 2012, 48, 86–96. [Google Scholar] [CrossRef]
- Lotshaw, D.P. Biophysical, pharmacological, and functional characteristics of cloned and native mammalian two-pore domain K+ channels. Cell Biochem. Biophys. 2007, 47, 209–256. [Google Scholar] [CrossRef]
- Romero, M.; Reboreda, A.; Sanchez, E.; Lamas, J.A. Newly developed blockers of the M-current do not reduce spike frequency adaptation in cultured mouse sympathetic neurons. Eur. J. Neurosci. 2004, 19, 2693–2702. [Google Scholar] [CrossRef]
- Brown, D.A.; Selyanko, A.A. Membrane currents underlying the cholinergic slow excitatory post-synaptic potential in the rat sympathetic ganglion. J. Physiol.-Lond. 1985, 365, 365–387. [Google Scholar] [CrossRef]
- Aiken, S.P.; Lampe, B.J.; Murphy, P.A.; Brown, B.S. Reduction of spike frequency adaptation and blockade of M-current in rat CA1 pyramidal neurones by linopirdine (DuP 996), a neurotransmitter release enhancer. Br. J. Pharmacol. 1995, 115, 1163–1168. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Ramirez, P.; Cadaveira-Mosquera, A.; Lamas, J.A.; Reboreda, A. Muscarinic modulation of TREK currents in mouse sympathetic superior cervical ganglion neurons. Eur. J. Neurosci. 2015, 42, 1797–1807. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Ramírez, P.; Reboreda, A.; Rueda-Ruzafa, L.; Herrera-Pérez, S.; Lamas, J.A. PIP2 Mediated Inhibition of TREK Potassium Currents by Bradykinin in Mouse Sympathetic Neurons. Int. J. Mol. Sci. 2020, 21, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.I.; Hwang, S.W. Depolarizing Effectors of Bradykinin Signaling in Nociceptor Excitation in Pain Perception. Biomol. Ther. (Seoul) 2018, 26, 255–267. [Google Scholar] [CrossRef]
- Kennard, L.E.; Chumbley, J.R.; Ranatunga, K.M.; Armstrong, S.J.; Veale, E.L.; Mathie, A. Inhibition of the human two-pore domain potassium channel, TREK-1, by fluoxetine and its metabolite norfluoxetine. Br. J. Pharmacol. 2005, 144, 821–829. [Google Scholar] [CrossRef] [Green Version]
- de la Pena, E.; Malkia, A.; Vara, H.; Caires, R.; Ballesta, J.J.; Belmonte, C.; Viana, F. The Influence of Cold Temperature on Cellular Excitability of Hippocampal Networks. PLoS ONE 2012, 7, e52475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, J.A. Mechanosensitive K2P channels, TREKking through the autonomic nervous system. In Mechanically Gated Channels and Their Regulation; Kamkin, A., Lozinsky, I., Eds.; Springer: Dordrecht, Germany, 2012; pp. 35–68. [Google Scholar]
- Maingret, F. TREK-1 is a heat-activated background K+ channel. EMBO J. 2000, 19, 2483–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.; Choe, C.; Kim, D. Thermosensitivity of the two-pore domain K+ channels TREK-2 and TRAAK. J. Physiol.-Lond. 2005, 564, 103–116. [Google Scholar] [CrossRef]
- Brown, D.A. M Currents. In Ion Channels: Volume 1; Narahashi, T., Ed.; Springer: Boston, MA, USA, 1988; pp. 55–94. [Google Scholar]
- Duprat, F.; Lesage, F.; Patel, A.J.; Fink, M.; Romey, G.; Lazdunski, M. The neuroprotective agent riluzole activates the two P domain K(+) channels TREK-1 and TRAAK. Mol. Pharmacol. 2000, 57, 906–912. [Google Scholar]
- Lesage, F.; Terrenoire, C.; Romey, G.; Lazdunski, M. Human TREK2, a 2P domain mechano-sensitive K+ channel with multiple regulations by polyunsaturated fatty acids, lysophospholipids, and Gs, Gi, and Gq protein-coupled receptors. J. Biol. Chem. 2000, 275, 28398–28405. [Google Scholar] [CrossRef] [Green Version]
- Pope, L.; Arrigoni, C.; Lou, H.; Bryant, C.; Gallardo-Godoy, A.; Renslo, A.R.; Minor, D.L., Jr. Protein and Chemical Determinants of BL-1249 Action and Selectivity for K2P Channels. ACS Chem. Neurosci. 2018, 9, 3153–3165. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Au, J.D.; Zou, H.L.; Cotten, J.F.; Yost, C.S. Potent activation of the human tandem pore domain K channel TRESK with clinical concentrations of volatile anesthetics. Anesth. Analg. 2004, 99, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Lesage, F. Pharmacology of neuronal background potassium channels. Neuropharmacology 2003, 44, 1–7. [Google Scholar] [CrossRef]
- Hadley, J.K.; Noda, M.; Selyanko, A.A.; Wood, I.C.; Abogadie, F.C.; Brown, D.A. Differential tetraethylammonium sensitivity of KCNQ1-4 potassium channels. Br. J. Pharmacol. 2000, 129, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Sandoz, G.; Douguet, D.; Chatelain, F.; Lazdunski, M.; Lesage, F. Extracellular acidification exerts opposite actions on TREK1 and TREK2 potassium channels via a single conserved histidine residue. Proc. Natl. Acad. Sci. USA 2009, 106, 14628–14633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, J.A.; Selyanko, A.A.; Brown, D.A. Effects of a cognition-enhancer, linopirdine (DuP 996), on M-type potassium currents (I-K(M)) and some other voltage- and ligand-gated membrane currents in rat sympathetic neurons. Eur. J. Neurosci. 1997, 9, 605–616. [Google Scholar] [CrossRef]
- Adams, P.R.; Brown, D.A.; Constanti, A. M-currents and other potassium currents in bullfrog sympathetic neurones. J. Physiol. 1982, 330, 537–572. [Google Scholar] [CrossRef]
- Goldstein, S.A.; Price, L.A.; Rosenthal, D.N.; Pausch, M.H. ORK1, a potassium-selective leak channel with two pore domains cloned from Drosophila melanogaster by expression in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1996, 93, 13256–13261. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.; Brown, D.A.; Milligan, G.; Willer, E.; Buckley, N.J.; Caulfield, M.P. Bradykinin excites rat sympathetic neurons by inhibition of M current through a mechanism involving B2 Receptors and Gaq/11. Neuron 1995, 14, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, C.C.; Zaika, O.; Tolstykh, G.P.; Shapiro, M.S. Regulation of neural KCNQ channels: Signalling pathways, structural motifs and functional implications. J Physiol.-Lond. 2008, 586, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.A.; Bockenhauer, D.; O’Kelly, I.; Zilberberg, N. Potassium leak channels and the KCNK family of two-P-domain subunits. Nat. Rev. Neurosci. 2001, 2, 175–184. [Google Scholar] [CrossRef]
- Lamas, J.A.; Rueda-Ruzafa, L.; Herrera-Perez, S. Ion Channels and Thermosensitivity: TRP, TREK, or Both? Int. J. Mol. Sci. 2019, 20, 2371. [Google Scholar] [CrossRef] [Green Version]
- Cruzblanca, H. An M2-like muscarinic receptor enhances a delayed rectifier K+ current in rat sympathetic neurones. Br. J. Pharmacol. 2006, 149, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruzblanca, H.; Koh, D.S.; Hille, B. Bradykinin inhibits M current via phospholipase C and Ca2+ release from IP3-sensitive Ca2+ stores in rat sympathetic neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 7151–7156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babbedge, R.; Dray, A.; Urban, L. Bradykinin depolarises the rat isolated superior cervical ganglion via B2 receptor activation. Neurosci. Lett. 1995, 193, 161–164. [Google Scholar] [CrossRef]
- Noel, J.; Sandoz, G.; Lesage, F. Molecular regulations governing TREK and TRAAK channel functions. Channels (Austin) 2011, 5, 402–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadley, J.K.; Passmore, G.M.; Tatulian, L.; Al-Qatari, M.; Ye, F.; Wickenden, A.D.; Brown, D.A. Stoichiometry of expressed KCNQ2/KCNQ3 potassium channels and subunit composition of native ganglionic M channels deduced from block by tetraethylammonium. J. Neurosci. 2003, 23, 5012–5019. [Google Scholar] [CrossRef]
- Wang, H.S.; Pan, Z.; Shi, W.; Brown, B.S.; Wymore, R.S.; Cohen, I.S.; Dixon, J.E.; McKinnon, D. KCNQ2 and KCNQ3 potassium channel subunits: Molecular correlates of the M-channel. Science 1998, 282, 1890–1893. [Google Scholar] [CrossRef] [Green Version]
- DeCoursey, T.E.; Dempster, J.; Hutter, O.F. Inward rectifier current noise in frog skeletal muscle. J. Physiol.-Lond. 1984, 349, 299–327. [Google Scholar] [CrossRef]
- Cobbett, P.; Legendre, P.; Mason, W.T. Characterization of three types of potassium current in cultured neurones of rat supraoptic nucleus area. J. Physiol.-Lond. 1989, 410, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Lipton, S.A.; Tauck, D.L. Voltage-dependent conductances of solitary ganglion cells dissociated from the rat retina. J. Physiol.-Lond. 1987, 385, 361–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.M.; Brown, B.S. Inhibition of M-current in cultured rat superior cervical ganglia by linopirdine: Mechanism of action studies. Neuropharmacology 1997, 36, 1747–1753. [Google Scholar] [CrossRef]
- Zhang, H.; Shepherd, N.; Creazzo, T.L. Temperature-sensitive TREK currents contribute to setting the resting membrane potential in embryonic atrial myocytes. J. Physiol.-Lond. 2008, 586, 3645–3656. [Google Scholar] [CrossRef]
- Springer, M.G.; Kullmann, P.H.; Horn, J.P. Virtual leak channels modulate firing dynamics and synaptic integration in rat sympathetic neurons: Implications for ganglionic transmission in vivo. J. Physiol.-Lond. 2015, 593, 803–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Pinna, J.; Lamas, J.A.; Gallego, R. Calcium current components in intact and dissociated adult mouse sympathetic neurons. Brain Res. 2002, 951, 227–236. [Google Scholar] [CrossRef]
- Reboreda, A.; Sanchez, E.; Romero, M.; Lamas, J.A. Intrinsic spontaneous activity and subthreshold oscillations in neurones of the rat dorsal column nuclei in culture. J. Physiol.-Lond. 2003, 551, 191–205. [Google Scholar] [CrossRef]
- Rae, J.; Cooper, K.; Gates, P.; Watsky, M. Low access resistance perforated patch recordings using amphotericin B. J Neurosci. Methods 1991, 37, 15–26. [Google Scholar] [CrossRef]
- Lamas, J.A. A hyperpolarization-activated cation current (Ih) contributes to resting membrane potential in rat superior cervical sympathetic neurones. Pflugers. Arch. Eur. J. Physiol. 1998, 436, 429–435. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivas-Ramírez, P.; Reboreda, A.; Rueda-Ruzafa, L.; Herrera-Pérez, S.; Lamas, J.A. Contribution of KCNQ and TREK Channels to the Resting Membrane Potential in Sympathetic Neurons at Physiological Temperature. Int. J. Mol. Sci. 2020, 21, 5796. https://doi.org/10.3390/ijms21165796
Rivas-Ramírez P, Reboreda A, Rueda-Ruzafa L, Herrera-Pérez S, Lamas JA. Contribution of KCNQ and TREK Channels to the Resting Membrane Potential in Sympathetic Neurons at Physiological Temperature. International Journal of Molecular Sciences. 2020; 21(16):5796. https://doi.org/10.3390/ijms21165796
Chicago/Turabian StyleRivas-Ramírez, Paula, Antonio Reboreda, Lola Rueda-Ruzafa, Salvador Herrera-Pérez, and Jose Antonio Lamas. 2020. "Contribution of KCNQ and TREK Channels to the Resting Membrane Potential in Sympathetic Neurons at Physiological Temperature" International Journal of Molecular Sciences 21, no. 16: 5796. https://doi.org/10.3390/ijms21165796
APA StyleRivas-Ramírez, P., Reboreda, A., Rueda-Ruzafa, L., Herrera-Pérez, S., & Lamas, J. A. (2020). Contribution of KCNQ and TREK Channels to the Resting Membrane Potential in Sympathetic Neurons at Physiological Temperature. International Journal of Molecular Sciences, 21(16), 5796. https://doi.org/10.3390/ijms21165796