Evaluation of the Individual and Combined Toxicity of Fumonisin Mycotoxins in Human Gastric Epithelial Cells



Abstract
:1. Introduction
2. Results
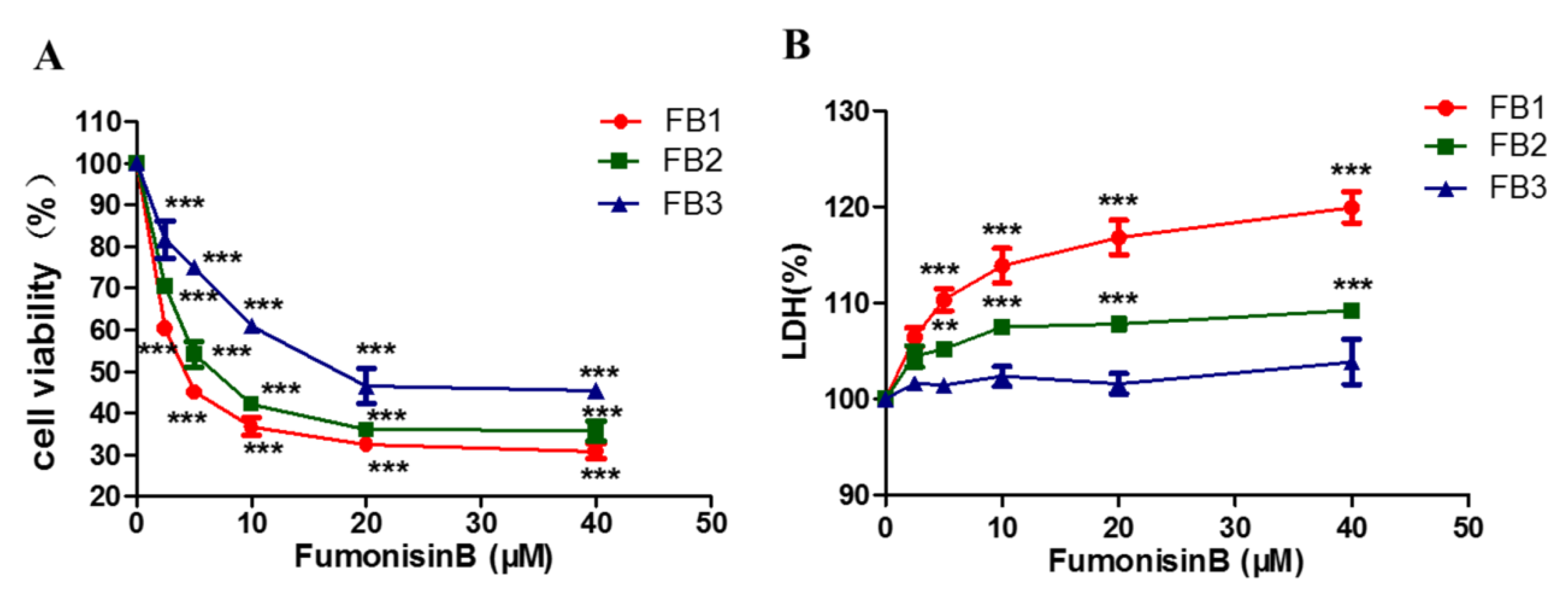
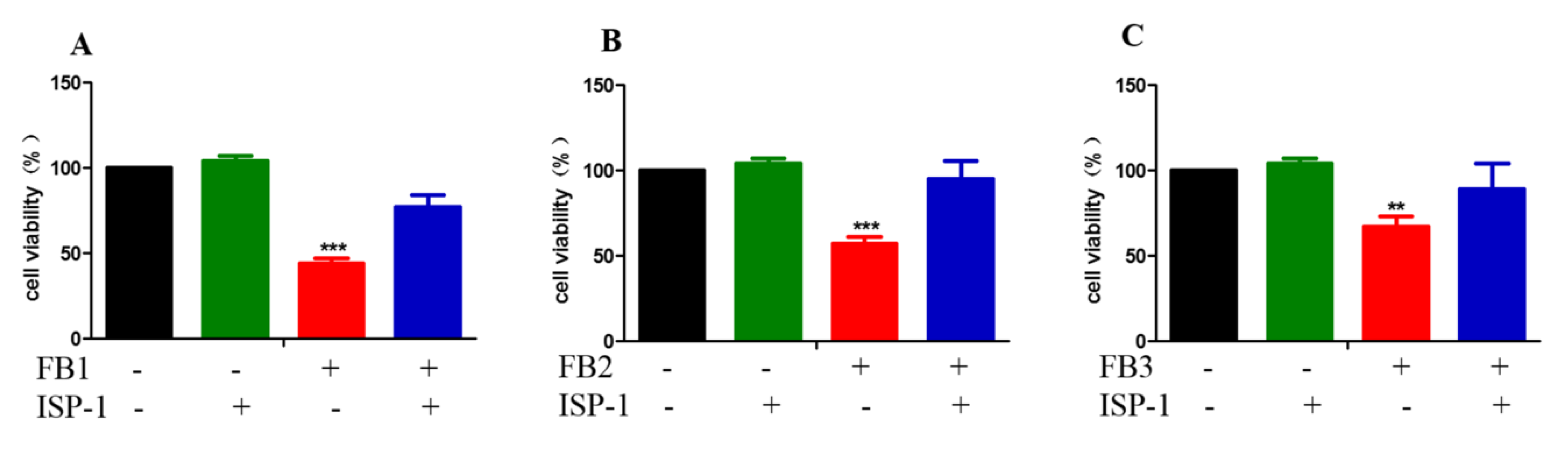
2.1. FBs Inhibited Cell Proliferation and Increased Lactic Dehydrogenase Levels in GES-1 Cells
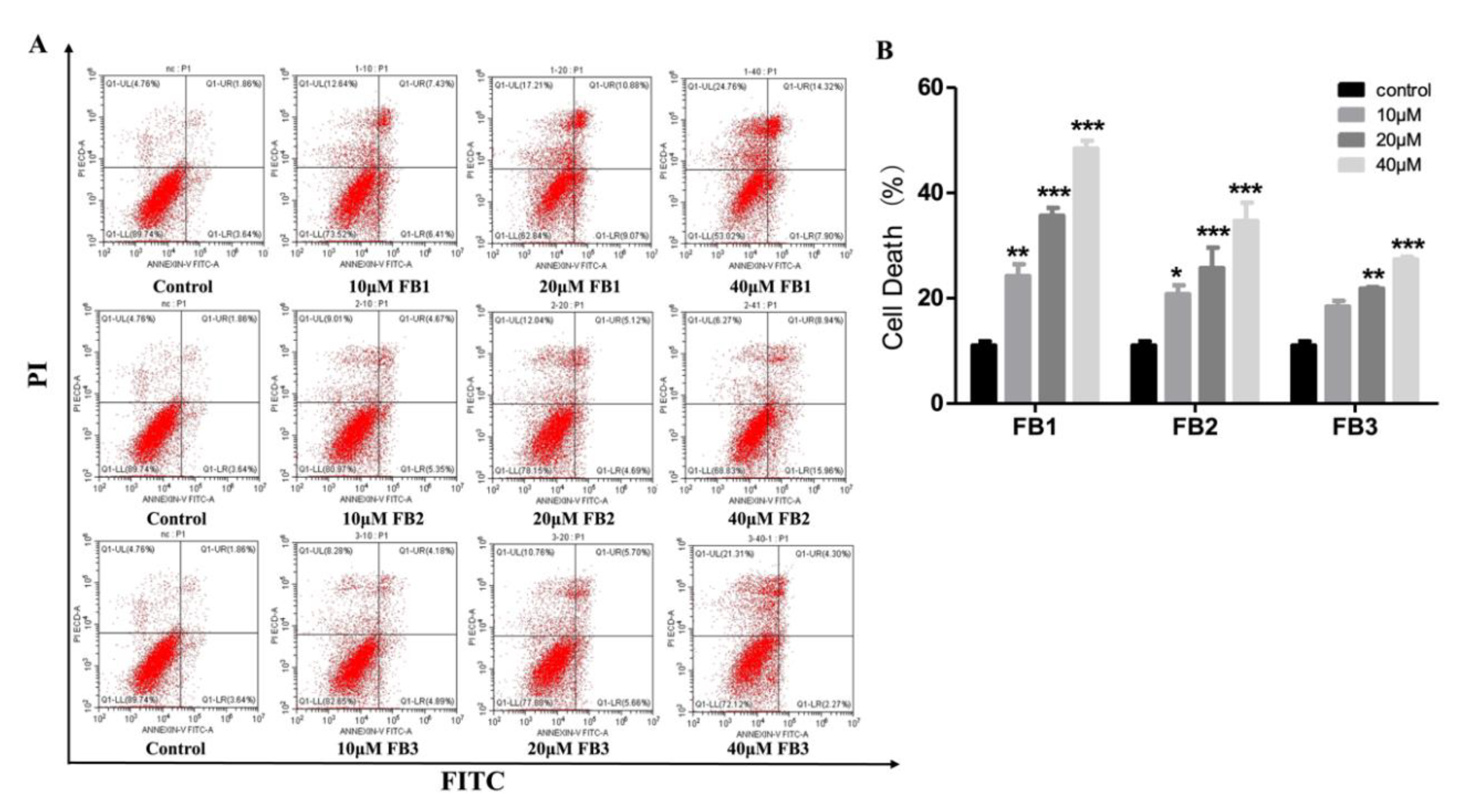
2.2. FBs Induced Cell Death in GES-1 Cells
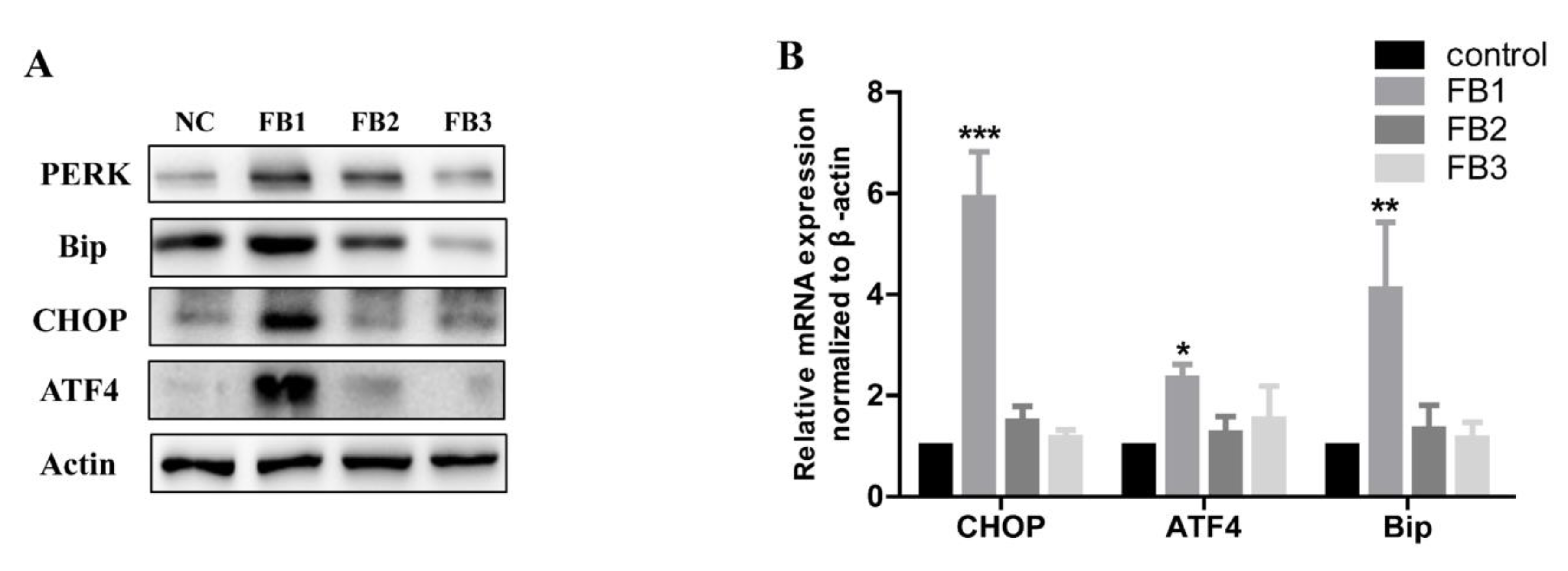
2.3. FBs Induced ER Stress in GES-1 Cells
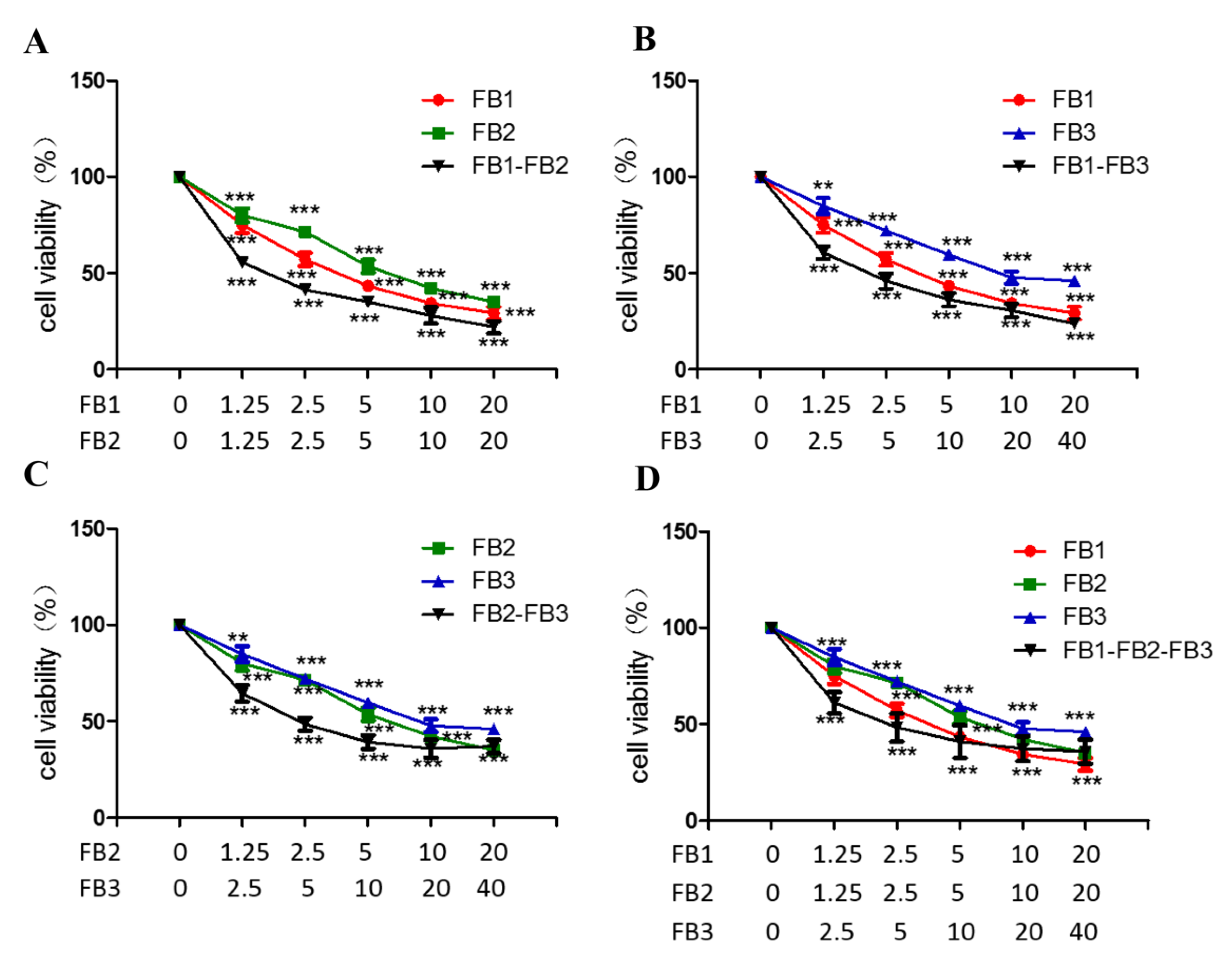
2.4. Combined Toxicity of FB1, FB2 and FB3 in GES-1 Cells
2.5. Disruption of Sphingolipid Metabolism Contributes to FBs Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture and Treatments
4.3. Cell viability Assay and Membrane Leakage Assay
4.4. Cell Death Analysis
4.5. Immunoblotting
4.6. Real-Time PCR
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FBs | Fumonisin Bs |
| FB1 | Fumonisin B1 |
| FB2 | Fumonisin B2 |
| FB3 | Fumonisin B3 |
| CCK-8 | Cell Counting Kit-8 |
| LDH | lactic dehydrogenase |
| Annexin V-FITC/PI | Annexin V-fluoresceine isothiocyanate/propidium iodide |
| ER | Endoplasmic reticulum |
| EDTA | Ethylene diamine tetraacetic acid |
| PERK | Protein kinase R-like ER kinase |
| CHOP | C/EBP homologous protein |
| Bip | Glucose-regulated protein 78 |
| ATF4 | Activating transcription factor 4 |
| GES-1 | Human gastric epithelial cell line |
| H-DMEM | Dulbecco’s modified eagle medium with high glucose |
| GIT | Gastrointestinal tract |
| SPT | Serine palmitoyltransferase |
| PIEC | Pig intestinal epithelial cells |
| ISP-1 | Myriocin |
| IC | Inhibitory concentration |
| CI | Combination index |
| DON | Deoxynivalenol |
References
- Edite Bezerra da Rocha, M.; Freire, F.d.C.O.; Erlan Feitosa Maia, F.; Izabel Florindo Guedes, M.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Mousavi Khaneghah, A.; Fakhri, Y.; Gahruie, H.H.; Niakousari, M.; Sant’Ana, A.S. Mycotoxins in cereal-based products during 24 years (1983–2017): A global systematic review. Trends Food Sci. Technol. 2019, 91, 95–105. [Google Scholar] [CrossRef]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.; Turner, P.C.; El-Nezami, H. Individual and combined cytotoxic effects of Fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins B1) on swine jejunal epithelial cells. Food Chem. Toxicol. 2013, 57, 276–283. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, X.; Chen, G.; Sun, S.; Yang, Y.; Zhu, Z.; Duan, C. The Major Fusarium Species Causing Maize Ear and Kernel Rot and Their Toxigenicity in Chongqing, China. Toxins 2018, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Cao, A.; Santiago, R.; Ramos, A.J.; Souto, X.C.; Aguin, O.; Malvar, R.A.; Butron, A. Critical environmental and genotypic factors for Fusarium verticillioides infection, fungal growth and fumonisin contamination in maize grown in northwestern Spain. Int. J. Food Microbiol. 2014, 177, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Esposito, F.; Fasano, E.; Scognamiglio, G.; Nardone, A.; Triassi, M.; Cirillo, T. Exposure assessment to fumonisins B1, B2 and B3 through consumption of gluten-free foodstuffs intended for people affected by celiac disease. Food Chem. Toxicol. 2016, 97, 395–401. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Beszterda, M.; Goliński, P. Occurrence of fumonisins in food—An interdisciplinary approach to the problem. Food Control 2012, 26, 491–499. [Google Scholar] [CrossRef]
- Gazzotti, T.; Zironi, E.; Lugoboni, B.; Barbarossa, A.; Piva, A.; Pagliuca, G. Analysis of fumonisins B1, B2 and their hydrolysed metabolites in pig liver by LC–MS/MS. Food Chem. 2011, 125, 1379–1384. [Google Scholar] [CrossRef]
- Yu, S.; Jia, B.; Yang, Y.; Liu, N.; Wu, A. Involvement of PERK-CHOP pathway in fumonisin B1- induced cytotoxicity in human gastric epithelial cells. Food Chem. Toxicol. 2020, 136, 111080. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, I.H.; Do, W.H.; Nam, W.S.; Li, H.; Jang, H.S.; Lee, C. Incidence and levels of deoxynivalenol, fumonisins and zearalenone contaminants in animal feeds used in Korea in 2012. Toxins 2013, 6, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Jiang, D.; Zheng, F.; Chen, J.; Li, W. Fumonisins B(1), B(2) and B(3) in corn products, wheat flour and corn oil marketed in Shandong province of China. Food Addit. Contam. Part B Surveill 2015, 8, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, D.; Moll, W. Fumonisin elimination and prospects for detoxification by enzymatic transformation. World Mycotoxin J. 2011, 4, 271–283. [Google Scholar] [CrossRef]
- Maenetje, P.W.; De Villiers, N.; Dutton, M.F. The use of isolated human lymphocytes in mycotoxin cytotoxicity testing. Int. J. Mol. Sci. 2008, 9, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, Y.; Liu, W.; Zhu, X.; Du, L.; Wang, Q. Fumonisin level in corn-based food and feed from Linxian County, a high-risk area for esophageal cancer in china. Food Chem. 2008, 106, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Regnier, M.; Polizzi, A.; Lukowicz, C.; Smati, S.; Lasserre, F.; Lippi, Y.; Naylies, C.; Laffitte, J.; Betoulieres, C.; Montagner, A.; et al. The protective role of liver X receptor (LXR) during fumonisin B1-induced hepatotoxicity. Arch. Toxicol. 2019, 93, 505–517. [Google Scholar] [CrossRef]
- Szabo, A.; Szabo-Fodor, J.; Kachlek, M.; Mezes, M.; Balogh, K.; Glavits, R.; Ali, O.; Zeebone, Y.Y.; Kovacs, M. Dose and Exposure Time-Dependent Renal and Hepatic Effects of Intraperitoneally Administered Fumonisin B(1) in Rats. Toxins 2018, 10, 465. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Jiang, Y.; Fan, Y.; Ma, Y.; Lei, H.; Su, J. Fumonisin B1 Induces Oxidative Stress and Breaks Barrier Functions in Pig Iliac Endothelium Cells. Toxins 2019, 11, 387. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.; Dekant, W.; Mally, A. Fumonisin B1 and the kidney: Modes of action for renal tumor formation by fumonisin B1 in rodents. Food Chem. Toxicol. 2012, 50, 3833–3846. [Google Scholar] [CrossRef]
- Liu, X.; Fan, L.; Yin, S.; Chen, H.; Hu, H. Molecular mechanisms of fumonisin B1-induced toxicities and its applications in the mechanism-based interventions. Toxicon 2019, 167, 1–5. [Google Scholar] [CrossRef]
- Van der Westhuizen, L.; Shephard, G.S.; Rheeder, J.P.; Burger, H.M. Individual fumonisin exposure and sphingoid base levels in rural populations consuming maize in South Africa. Food Chem. Toxicol. 2010, 48, 1698–1703. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Guo, X.; Li, J.; Fan, L.; Hu, H. Fumonisin B1 induces autophagic cell death via activation of ERN1-MAPK8/9/10 pathway in monkey kidney MARC-145 cells. Arch. Toxicol. 2016, 90, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Direito, G.M.; Almeida, A.P.; Aquino, S.; Dos Reis, T.A.; Pozzi, C.R.; Correa, B. Evaluation of sphingolipids in Wistar rats treated to prolonged and single oral doses of fumonisin b(1). Int. J. Mol. Sci. 2009, 10, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, S.; Tan, Y.; Liu, N.; Wu, A. Individual and Combined Cytotoxic Effects of Co-Occurring Deoxynivalenol Family Mycotoxins on Human Gastric Epithelial Cells. Toxins 2017, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, S.; Zhang, M.; Yang, L.; Cheng, B.; Li, J.; Shan, A. Individual and combined effects of Fusarium toxins on apoptosis in PK15 cells and the protective role of N-acetylcysteine. Food Chem. Toxicol. 2018, 111, 27–43. [Google Scholar] [CrossRef]
- Xue, K.S.; Qian, G.; Lin, S.; Su, J.; Tang, L.; Gelderblom, W.C.A.; Riley, R.T.; Phillips, T.D.; Wang, J.S. Modulation of pre-neoplastic biomarkers induced by sequential aflatoxin B1 and fumonisin B1 exposure in F344 rats treated with UPSN clay. Food Chem. Toxicol. 2018, 114, 316–324. [Google Scholar] [CrossRef]
- Li, R.; Tao, B.; Pang, M.; Liu, Y.; Dong, J. Natural occurrence of fumonisins B1 and B2 in maize from three main maize-producing provinces in China. Food Control 2015, 50, 838–842. [Google Scholar] [CrossRef]
- Bryła, M.; Roszko, M.; Szymczyk, K.; Jędrzejczak, R.; Obiedziński, M.W. Fumonisins and their masked forms in maize products. Food Control 2016, 59, 619–627. [Google Scholar] [CrossRef]
- Hahn, I.; Nagl, V.; Schwartz-Zimmermann, H.E.; Varga, E.; Schwarz, C.; Slavik, V.; Reisinger, N.; Malachova, A.; Cirlini, M.; Generotti, S.; et al. Effects of orally administered fumonisin B(1) (FB(1)), partially hydrolysed FB(1), hydrolysed FB(1) and N-(1-deoxy-D-fructos-1-yl) FB(1) on the sphingolipid metabolism in rats. Food Chem. Toxicol. 2015, 76, 11–18. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, S.; Liu, N.; Xu, H.; Gong, Y.; Wu, Y.; Wang, P.; Su, X.; Liao, Y.; De Saeger, S.; et al. Transcription Factor FOXO3a Is a Negative Regulator of Cytotoxicity of Fusarium mycotoxin in GES-1 Cells. Toxicol. Sci. 2018, 166, 370–381. [Google Scholar] [CrossRef]
- Gajecka, M.; Waskiewicz, A.; Zielonka, L.; Golinski, P.; Rykaczewska, A.; Lisieska-Zolnierczyk, S.; Gajecki, M.T. Mycotoxin levels in the digestive tissues of immature gilts exposed to zearalenone and deoxynivalenol. Toxicon 2018, 153, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Devreese, M.; De Baere, S.; Martel, A.; Van Immerseel, F.; Croubels, S. Impact of Fusarium mycotoxins on hepatic and intestinal mRNA expression of cytochrome P450 enzymes and drug transporters, and on the pharmacokinetics of oral enrofloxacin in broiler chickens. Food Chem. Toxicol. 2017, 101, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Debellis, L.; Garbetta, A.; De Girolamo, A.; Schena, R.; Portincasa, P.; Visconti, A. Influence on functional parameters of intestinal tract induced by short-term exposure to fumonisins contaminated corn chyme samples. Food Chem. Toxicol. 2014, 66, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, Q.; Zhang, Z.; Lin, P.; Lei, L.; Wang, A.; Jin, Y. Endoplasmic Reticulum Stress Cooperates in Zearalenone-Induced Cell Death of RAW 264.7 Macrophages. Int. J. Mol. Sci. 2015, 16, 19780–19795. [Google Scholar] [CrossRef] [Green Version]
- Ashton, J.C. Drug combination studies and their synergy quantification using the Chou-Talalay method--letter. Cancer Res. 2015, 75, 2400. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Hojjati, M.R.; Li, Z.; Zhou, H.; Tang, S.; Huan, C.; Ooi, E.; Lu, S.; Jiang, X.C. Effect of myriocin on plasma sphingolipid metabolism and atherosclerosis in apoE-deficient mice. J. Biol. Chem. 2005, 280, 10284–10289. [Google Scholar] [CrossRef] [Green Version]
- Missmer, S.A.; Suarez, L.; Felkner, M.; Wang, E.; Merrill, A.H., Jr.; Rothman, K.J.; Hendricks, K.A. Exposure to fumonisins and the occurrence of neural tube defects along the Texas-Mexico border. Environ. Health Perspect. 2006, 114, 237–241. [Google Scholar] [CrossRef]
- Assuncao, R.; Pinhao, M.; Loureiro, S.; Alvito, P.; Silva, M.J. A multi-endpoint approach to the combined toxic effects of patulin and ochratoxin a in human intestinal cells. Toxicol. Lett. 2019, 313, 120–129. [Google Scholar] [CrossRef]
- Sobral, M.M.C.; Faria, M.A.; Cunha, S.C.; Miladinovic, B.; Ferreira, I.M. Transport of mycotoxins across human gastric NCI-N87 and intestinal Caco-2cell models. Food Chem. Toxicol. 2019, 131, 110595. [Google Scholar] [CrossRef]
- Bouhet, S.; Le Dorze, E.; Peres, S.; Fairbrother, J.M.; Oswald, I.P. Mycotoxin fumonisin B1 selectively down-regulates the basal IL-8 expression in pig intestine: In vivo and in vitro studies. Food Chem. Toxicol. 2006, 44, 1768–1773. [Google Scholar] [CrossRef]
- Gao, Y.N.; Wang, J.Q.; Li, S.L.; Zhang, Y.D.; Zheng, N. Aflatoxin M1 cytotoxicity against human intestinal Caco-2 cells is enhanced in the presence of other mycotoxins. Food Chem. Toxicol. 2016, 96, 79–89. [Google Scholar] [CrossRef]
- Gu, M.J.; Han, S.E.; Hwang, K.; Mayer, E.; Reisinger, N.; Schatzmayr, D.; Park, B.C.; Han, S.H.; Yun, C.H. Hydrolyzed fumonisin B1 induces less inflammatory responses than fumonisin B1 in the co-culture model of porcine intestinal epithelial and immune cells. Toxicol. Lett. 2019, 305, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, A.; Shi, Z.; He, C.; Ding, J.; Wang, X.; Ma, J.; Zhang, H. A mitochondria-mediated apoptotic pathway induced by deoxynivalenol in human colon cancer cells. Toxicol. In Vitro 2012, 26, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Dietrich, D.R.; O’Brien, E. In vitro investigation of individual and combined cytotoxic effects of ochratoxin A and other selected mycotoxins on renal cells. Toxicol. In Vitro 2006, 20, 332–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, R.B.; Phulukdaree, A.; Chuturgoon, A.A. Fumonisin B1 induces oxidative stress in oesophageal (SNO) cancer cells. Toxicon 2018, 141, 104–111. [Google Scholar] [CrossRef]
- Follmann, W.; Behm, C.; Degen, G.H. Toxicity of the mycotoxin citrinin and its metabolite dihydrocitrinone and of mixtures of citrinin and ochratoxin A in vitro. Arch. Toxicol. 2014, 88, 1097–1107. [Google Scholar] [CrossRef]
- Nagata, S.; Zhou, X.; Okamura, H. Antagonistic and Synergistic Effects of Antifouling Chemicals in Mixture. Encycl. Ecol. 2008, 194–203. [Google Scholar] [CrossRef]
- Fernández-Alba, A.R.; Hernando, M.D.; Piedra, L.; Chisti, Y. Toxicity evaluation of single and mixed antifouling biocides measured with acute toxicity bioassays. Anal. Chim. Acta 2002, 456, 303–312. [Google Scholar]
- Smith, M.C.; Gheux, A.; Coton, M.; Madec, S.; Hymery, N.; Coton, E. In vitro co-culture models to evaluate acute cytotoxicity of individual and combined mycotoxin exposures on Caco-2, THP-1 and HepaRG human cell lines. Chem. Biol. Interact. 2018, 281, 51–59. [Google Scholar] [CrossRef]
- Meneely, J.P.; Hajslova, J.; Krska, R.; Elliott, C.T. Assessing the combined toxicity of the natural toxins, aflatoxin B1, fumonisin B1 and microcystin-LR by high content analysis. Food Chem. Toxicol. 2018, 121, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Soriano, J.M.; Gonzalez, L.; Catala, A.I. Mechanism of action of sphingolipids and their metabolites in the toxicity of fumonisin B1. Prog. Lipid. Res. 2005, 44, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Howard, P.C.; Couch, L.H.; Patton, R.E.; Eppley, R.M.; Doerge, D.R.; Churchwell, M.I.; Marques, M.M.; Okerberg, C.V. Comparison of the toxicity of several fumonisin derivatives in a 28-day feeding study with female B6C3F(1) mice. Toxicol. Appl. Pharmacol. 2002, 185, 153–165. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dm | M | r | |
|---|---|---|---|
| FB1 | 4.43919 | −0.70722 | 0.97471 |
| FB2 | 7.45614 | −0.75675 | 0.98928 |
| FB3 | 22.43032 | −0.69375 | 0.96569 |
| FB1 + FB2 | 1.59058 | −0.51736 | 0.99049 |
| FB1 + FB3 | 4.39135 | −0.566 | 0.98716 |
| FB2 + FB3 | 5.972 | −0.40263 | 0.95451 |
| FB1 + FB2 + FB3 | 5.46792 | −0.36484 | 0.94273 |
| Mycotoxin | Combination Ratio | IC10 | IC25 | IC50 | IC75 |
|---|---|---|---|---|---|
| Combination Index | |||||
| FB1:FB2 | 1:1 | 0.17 | 0.31 | 0.57 | 1.05 |
| FB1:FB3 | 1:2 | 0.30 | 0.46 | 0.69 | 1.04 |
| FB2:FB3 | 1:2 | 0.06 | 0.20 | 0.66 | 2.27 |
| FB1:FB2:FB3 | 1:1:2 | 0.06 | 0.28 | 1.23 | 5.41 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Jia, B.; Liu, N.; Yu, D.; Wu, A. Evaluation of the Individual and Combined Toxicity of Fumonisin Mycotoxins in Human Gastric Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 5917. https://doi.org/10.3390/ijms21165917
Yu S, Jia B, Liu N, Yu D, Wu A. Evaluation of the Individual and Combined Toxicity of Fumonisin Mycotoxins in Human Gastric Epithelial Cells. International Journal of Molecular Sciences. 2020; 21(16):5917. https://doi.org/10.3390/ijms21165917
Chicago/Turabian StyleYu, Song, Bingxuan Jia, Na Liu, Dianzhen Yu, and Aibo Wu. 2020. "Evaluation of the Individual and Combined Toxicity of Fumonisin Mycotoxins in Human Gastric Epithelial Cells" International Journal of Molecular Sciences 21, no. 16: 5917. https://doi.org/10.3390/ijms21165917
APA StyleYu, S., Jia, B., Liu, N., Yu, D., & Wu, A. (2020). Evaluation of the Individual and Combined Toxicity of Fumonisin Mycotoxins in Human Gastric Epithelial Cells. International Journal of Molecular Sciences, 21(16), 5917. https://doi.org/10.3390/ijms21165917