Carbon Fibers as a New Type of Scaffold for Midbrain Organoid Development

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Scaffolds Characterization
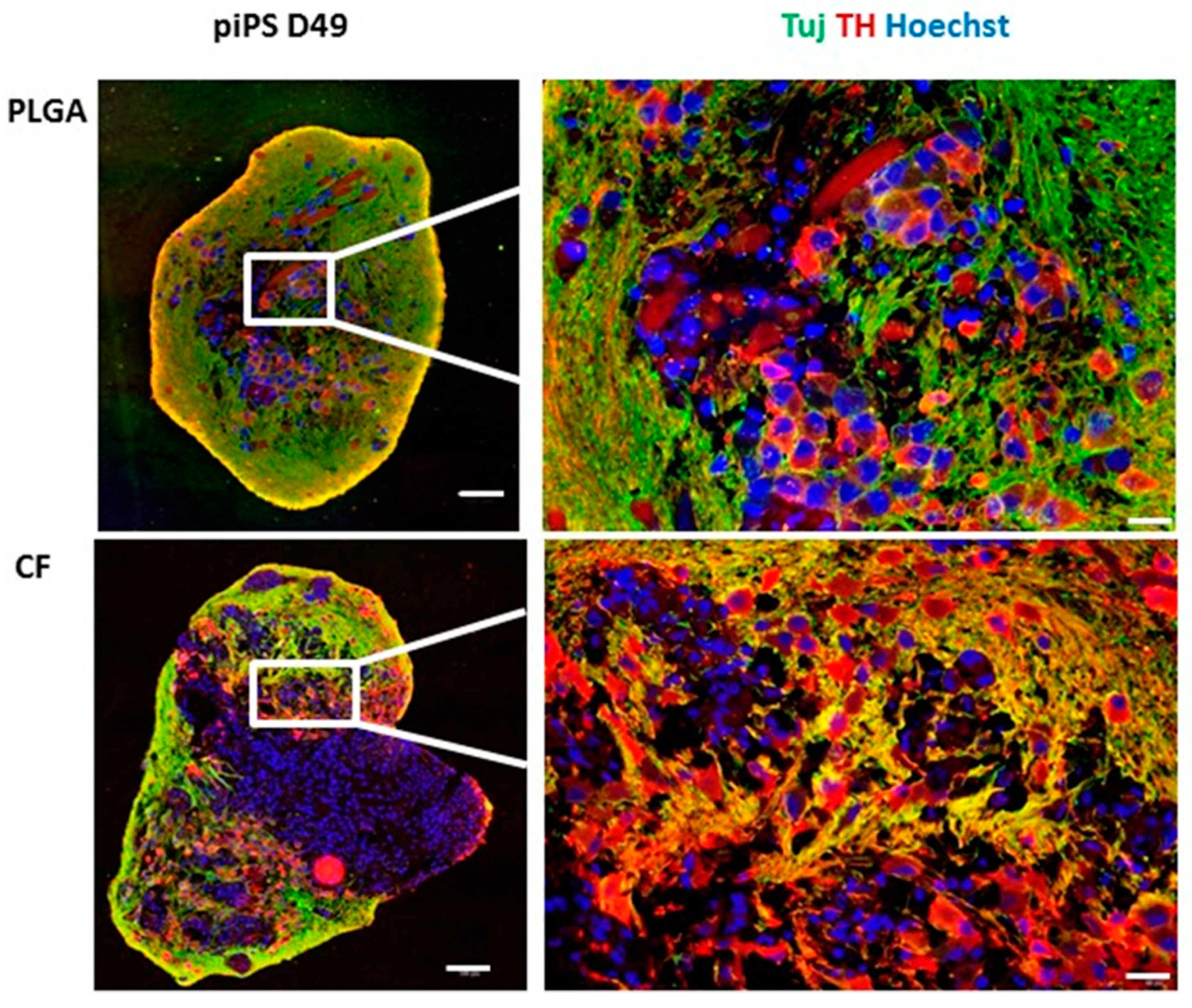
2.2. Generation and Characterization of Organoids from iPSCs
3. Discussion
4. Materials and Methods
4.1. Culture of Human Induced Pluripotent Stem Cells and the Generation of Organoids
4.2. Immunohistochemistry
4.3. RNA Extraction, Reverse Transcription, Real-Time RT-PCR
4.4. Carbon Fiber Synthesis
4.5. Obtaining PLGA Fibers
4.6. Fiber Morphology and Microstructure Characterization
4.7. Fiber Degradation Test
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BP | basal plate |
| CF | carbon fiber |
| EB | embryoid body |
| FL | fiber-less |
| iPSC | induced pluripotent stem cell |
| Lmx1a | LIM homeobox transcription factor |
| mDA | midbrain dopaminergic neuron |
| mFP | midbrain floor plate (mFP) |
| Msx1 | homologue 1 muscle segment homeobox |
| Nurr1 | Nr4a2, nuclear receptor 4a2 |
| PD | Parkinson’s disease |
| piPS | protein-induced pluripotent stem cells |
| Pitx3 | pituitary homeobox 3 or paired-like homeodomain transcription factor 3 |
| PLGA | copolymer poly-(lactic-co-glycolic acid) |
| SEM | scanning electron microscopy |
| TH | tyrosine hydroxylase |
| Tuj1 | neuron-specific class III β-tubulin |
References
- Shi, Y.; Inoue, H.; Wu, J.C.; Yamanaka, S. Induced pluripotent stem cell technology: A decade of progress. Nat. Rev. Drug Discov. 2017, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Sułkowski, M.; Konieczny, P.; Chlebanowska, P.; Majka, M. Introduction of Exogenous HSV-TK Suicide Gene Increases Safety of Keratinocyte-Derived Induced Pluripotent Stem Cells by Providing Genetic “Emergency Exit” Switch. Int. J. Mol. Sci. 2018, 19, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, J.; Xiao, Y.; Sun, A.X.; Cukuroglu, E.; Tran, H.D.; Göke, J.; Tan, Z.Y.; Saw, T.Y.; Tan, C.P.; Lokman, H.; et al. Midbrain-like Organoids from Human Pluripotent Stem Cells Contain Functional Dopaminergic and Neuromelanin-Producing Neurons. Cell Stem Cell 2016, 19, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.; Tryggvason, U.; Deng, Q.; Friling, S.; Alekseenko, Z.; Robert, B.; Perlmann, T.; Ericson, J. Identification of intrinsic determinants of midbrain dopamine neurons. Cell 2006, 124, 393–405. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Andersson, E.; Hedlund, E.; Alekseenko, Z.; Coppola, E.; Panman, L.; Millonig, J.H.; Brunet, J.F.; Ericson, J.; Perlmann, T. Specific and integrated roles of Lmx1a, Lmx1b and Phox2a in ventral midbrain development. Development 2011, 138, 3399–3408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, E.; Jensen, J.B.; Parmar, M.; Guillemot, F.; Björklund, A. Development of the mesencephalic dopaminergic neuron system is compromised in the absence of neurogenin 2. Development 2006, 133, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Arenas, E.; Denham, M.; Villaescusa, J.C. How to make a midbrain dopaminergic neuron. Development 2015, 142, 1918–1936. [Google Scholar] [CrossRef] [Green Version]
- Smidt, M.P.; Asbreuk, C.H.J.; Cox, J.J.; Chen, H.; Johnson, R.L.; Burbach, J.P.H. A second independent pathway for development of mesencephalic dopaminergic neurons requires Lmx1b. Nat. Neurosci. 2000, 16, 337–341. [Google Scholar] [CrossRef]
- Zetterström, R.H.; Williams, R.; Perlmann, T.; Olson, L. Cellular expression of the immediate early transcription factors Nurr1 and NGFI-B suggests a gene regulatory role in several brain regions including the nigrostriatal dopamine system. Mol. Brain Res. 1996, 41, 111–120. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Ho, H.Y.; Kuehner, E.; Zhao, S.; Li, M. Pitx3 regulates tyrosine hydroxylase expression in the substantia nigra and identifies a subgroup of mesencephalic dopaminergic progenitor neurons during mouse development. Dev. Biol. 2005, 282, 467–479. [Google Scholar] [CrossRef] [Green Version]
- van den Munckhof, P.; Luk, K.C.; Ste-Marie, L.; Montgomery, J.; Blanchet, P.J.; Sadikot, A.F.; Drouin, J. Pitx3 is required for motor activity and for survival of a subset of midbrain dopaminergic neurons. Development 2003, 130, 2535–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasca, S.P. Assembling human brain organoids. Science 2019, 363, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Kratochvil, M.J.; Seymour, A.J.; Li, T.L.; Paşca, S.P.; Kuo, C.J.; Heilshorn, S.C. Engineered materials for organoid systems. Nat. Rev. Mater. 2019, 4, 606–622. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Corsini, N.S.; Wolfinger, S.; Gustafson, E.H.; Phillips, A.W.; Burkard, T.R.; Otani, T.; Livesey, F.J.; Knoblich, J.A. Guided self-organization and cortical plate formation in human brain organoids. Nat. Biotechnol. 2017, 35, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranga, A.; Girgin, M.; Meinhardt, A.; Eberle, D.; Caiazzo, M.; Tanaka, E.M.; Lutolf, M.P. Neural tube morphogenesis in synthetic 3D microenvironments. Proc. Natl. Acad. Sci. USA 2016, 113, E6831–E6839. [Google Scholar] [CrossRef] [Green Version]
- Debnath, J.; Muthuswamy, S.K.; Brugge, J.S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods 2003, 30, 256–268. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular Matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef]
- Chlebanowska, P.; Tejchman, A.; Su, M.; Skrzypek, K.; Majka, M. Use of 3D Organoids as a Model to Study Idiopathic Form of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 694. [Google Scholar] [CrossRef] [Green Version]
- Fraczek-Szczypta, A.; Bogun, M.; Blazewicz, S. Carbon fibers modified with carbon nanotubes. J. Mater. Sci. 2009, 44, 4721–4727. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, W.; Conneely, O.M.; He, Y.; Jankovic, J.; Appel, S.H. Reduced Nurrl Expression Increases the Vulnerability of Mesencephalic Dopamine Neurons to MPTP-Induced Injury. J. Neurochem. 2002, 73, 2218–2221. [Google Scholar]
- Saucedo-Cardenas, O.; Quintana-Hau, J.D.; Le, W.D.; Smidt, M.P.; Cox, J.J.; De Mayo, F.; Burbach, J.P.H.; Conneely, O.M. Nurr1 is essential for the induction of the dopaminergic phenotype and the survival of ventral mesencephalic late dopaminergic precursor neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 4013–4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, F.M.J.; Smits, S.M.; Noorlander, C.W.; von Oerthel, L.; van der Linden, A.J.A.; Burbach, J.P.H.; Smidt, M.P. Retinoic acid counteracts developmental defects in the substantia nigra caused by Pitx3 deficiency. Development 2007, 134, 2673–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadkhodaei, B.; Ito, T.; Joodmardi, E.; Mattsson, B.; Rouillard, C.; Carta, M.; Muramatsu, S.I.; Sumi-Ichinose, C.; Nomura, T.; Metzger, D.; et al. Nurr1 is required for maintenance of maturing and adult midbrain dopamine neurons. J. Neurosci. 2009, 29, 15923–15932. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, F.M.J.; van Erp, S.; van der Linden, A.J.A.; von Oerthel, L.; Burbach, P.H.; Smidt, M.P. Pitx3 potentiates Nurr1 in dopamine neuron terminal differentiation through release of SMRT-mediated repression. Development 2009, 136, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Volpicelli, F.; de Gregorio, R.; Pulcrano, S.; Perrone-Capano, C.; Porzio, U.; Bellenchi, G.C. Direct regulation of Pitx3 expression by Nurr1 in culture and in developing mouse midbrain. PLoS ONE 2012, 7, e30661. [Google Scholar] [CrossRef] [Green Version]
- Rajzer, I.; Menaszek, E.; Bacakova, L.; Rom, M.; Blazewicz, M. In vitro and in vivo studies on biocompatibility of carbon fibres. J. Mater. Sci. Mater. Med. 2010, 21, 2611–2622. [Google Scholar] [CrossRef] [Green Version]
- Blazewicz, M.; Piekarczyk, I.; Menaszek, E.; Haberko, K. Polymer and carbon fibers with HAp nanopowder: Properties and biocompatibility of degradation products. Eur. Cells Mater. 2004, 7, 47. [Google Scholar]
- Lü, J.M.; Wang, X.; Marin-Muller, C.; Wang, H.; Lin, P.H.; Yao, Q.; Chen, C. Current advances in research and clinical applications of PLGA-based nanotechnology. Expert Rev. Mol. Diagn. 2009, 9, 325–341. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fibers | Fraction of Small Pores, 4–15 nm (%) | Fraction of Large Pores, 150–750 nm (%) |
|---|---|---|
| PLGA | 0.67 | 2.46 |
| CF | 31.24 | 9.15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejchman, A.; Znój, A.; Chlebanowska, P.; Frączek-Szczypta, A.; Majka, M. Carbon Fibers as a New Type of Scaffold for Midbrain Organoid Development. Int. J. Mol. Sci. 2020, 21, 5959. https://doi.org/10.3390/ijms21175959
Tejchman A, Znój A, Chlebanowska P, Frączek-Szczypta A, Majka M. Carbon Fibers as a New Type of Scaffold for Midbrain Organoid Development. International Journal of Molecular Sciences. 2020; 21(17):5959. https://doi.org/10.3390/ijms21175959
Chicago/Turabian StyleTejchman, Anna, Agnieszka Znój, Paula Chlebanowska, Aneta Frączek-Szczypta, and Marcin Majka. 2020. "Carbon Fibers as a New Type of Scaffold for Midbrain Organoid Development" International Journal of Molecular Sciences 21, no. 17: 5959. https://doi.org/10.3390/ijms21175959
APA StyleTejchman, A., Znój, A., Chlebanowska, P., Frączek-Szczypta, A., & Majka, M. (2020). Carbon Fibers as a New Type of Scaffold for Midbrain Organoid Development. International Journal of Molecular Sciences, 21(17), 5959. https://doi.org/10.3390/ijms21175959