A Novel Urease Inhibitor of Ruminal Microbiota Screened through Molecular Docking




Abstract
:1. Introduction
2. Results
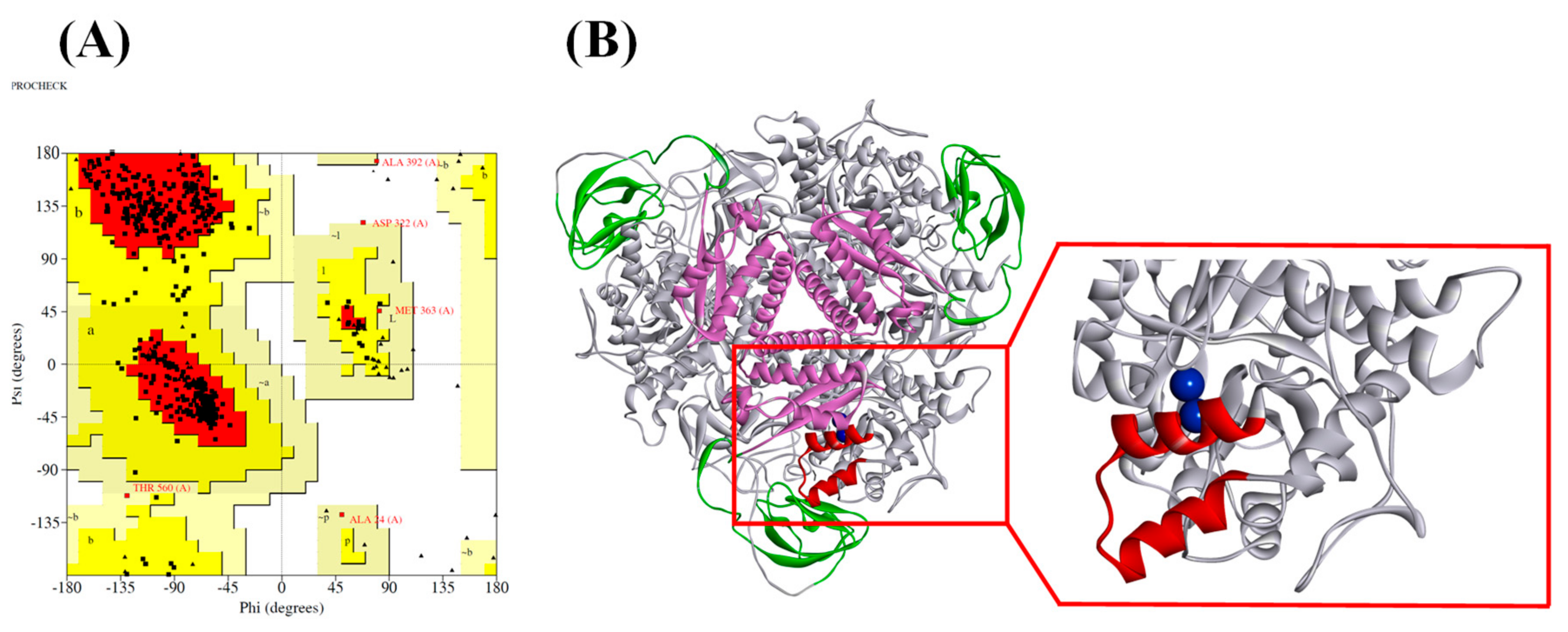
2.1. Ruminal Metagenomic Urease Homology Modeling and Validation
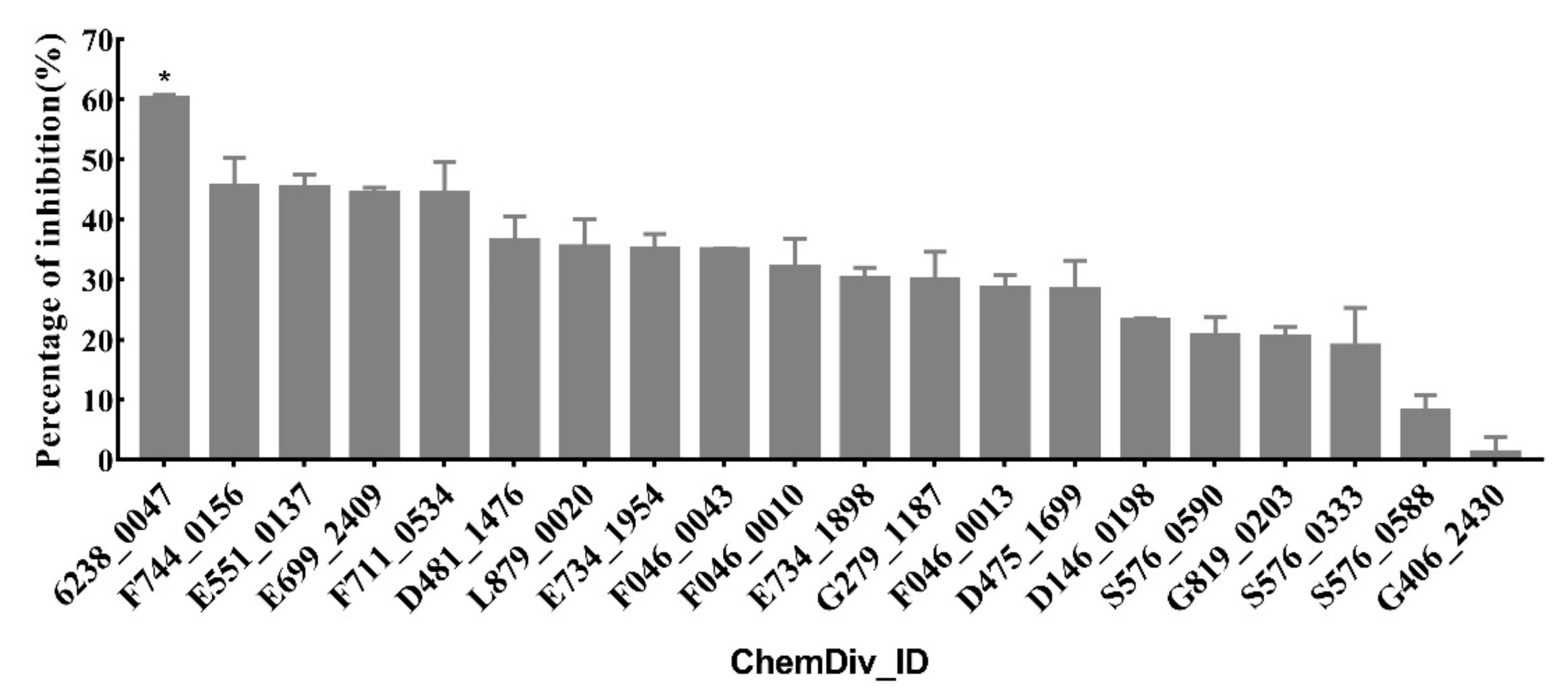
2.2. Virtual Screening for Candidate Compounds
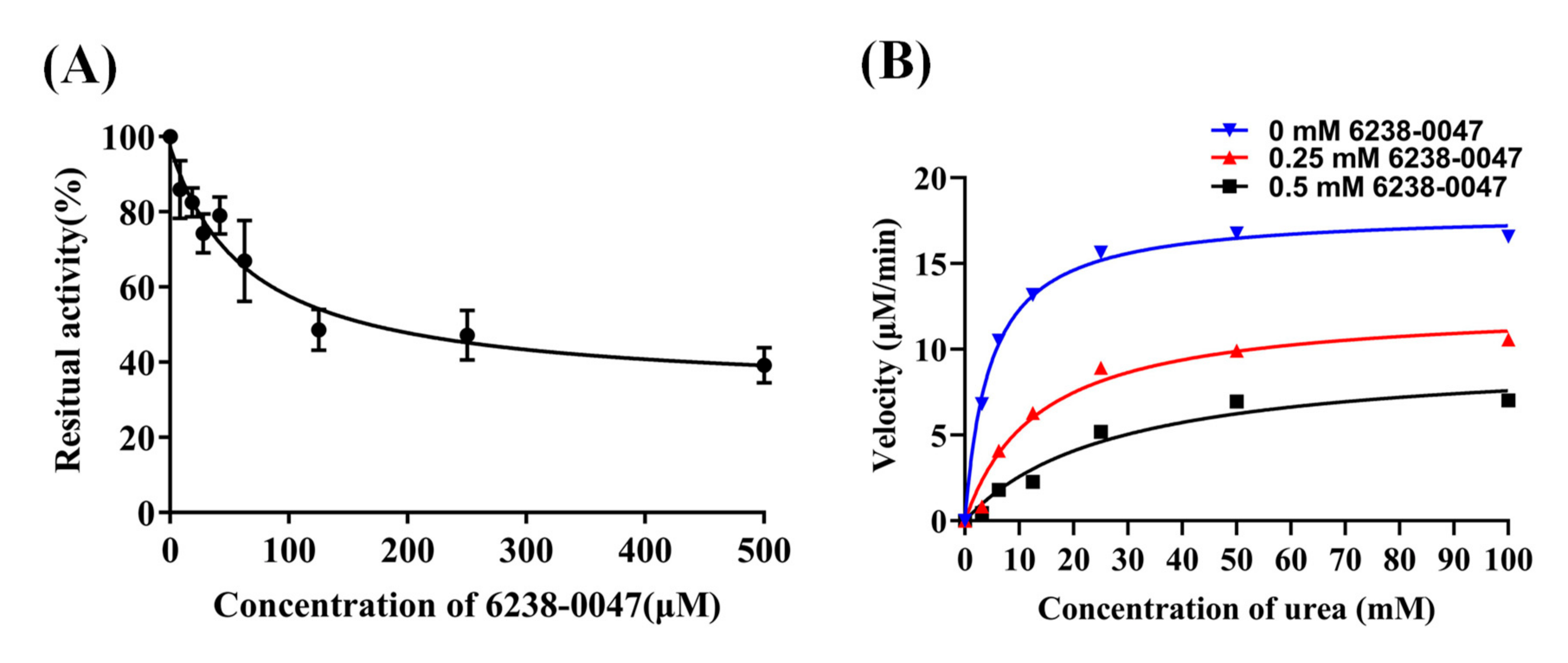
2.3. Urease Enzyme Inhibition Rate, IC50 Value, and Inhibition Mode of Compounds
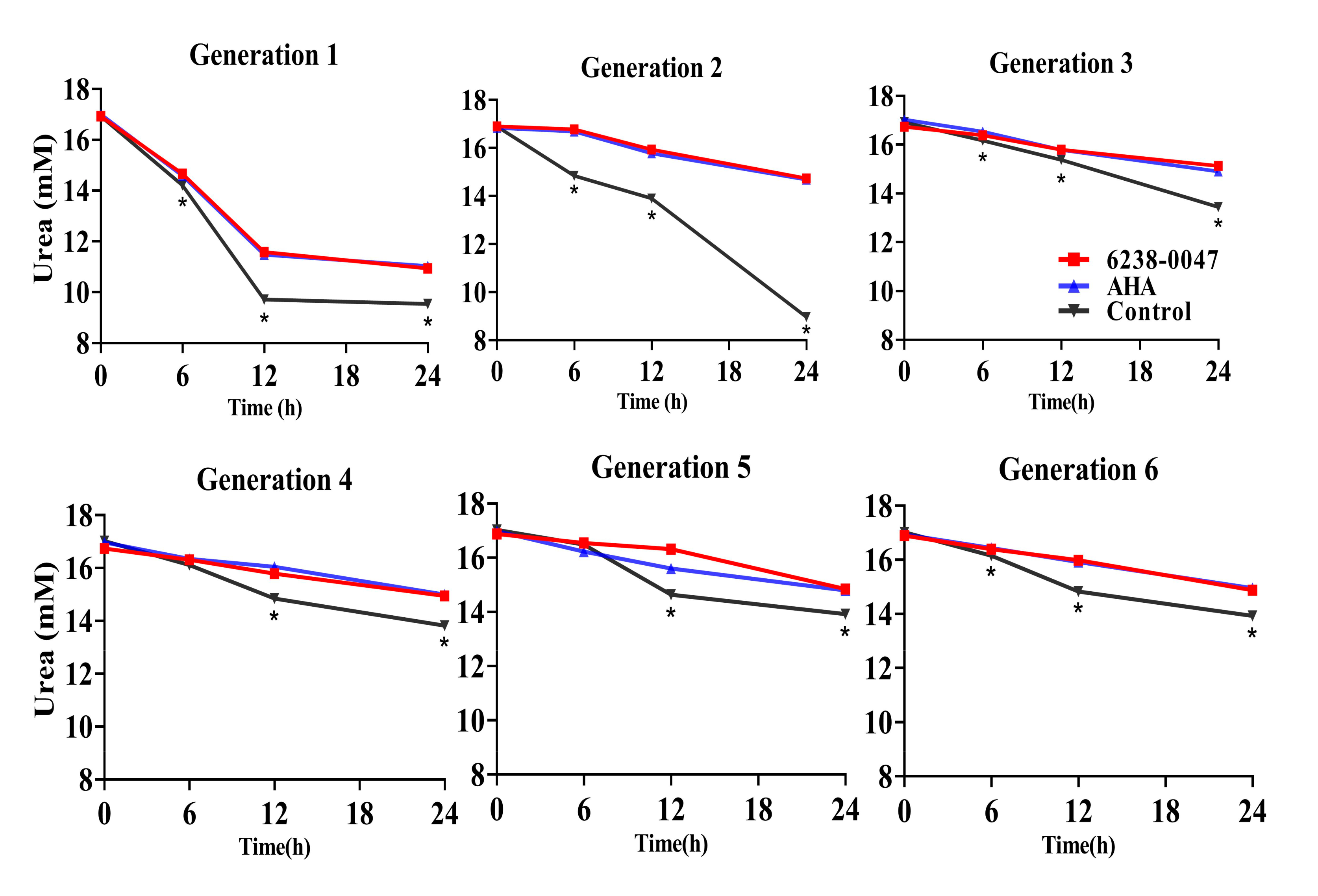
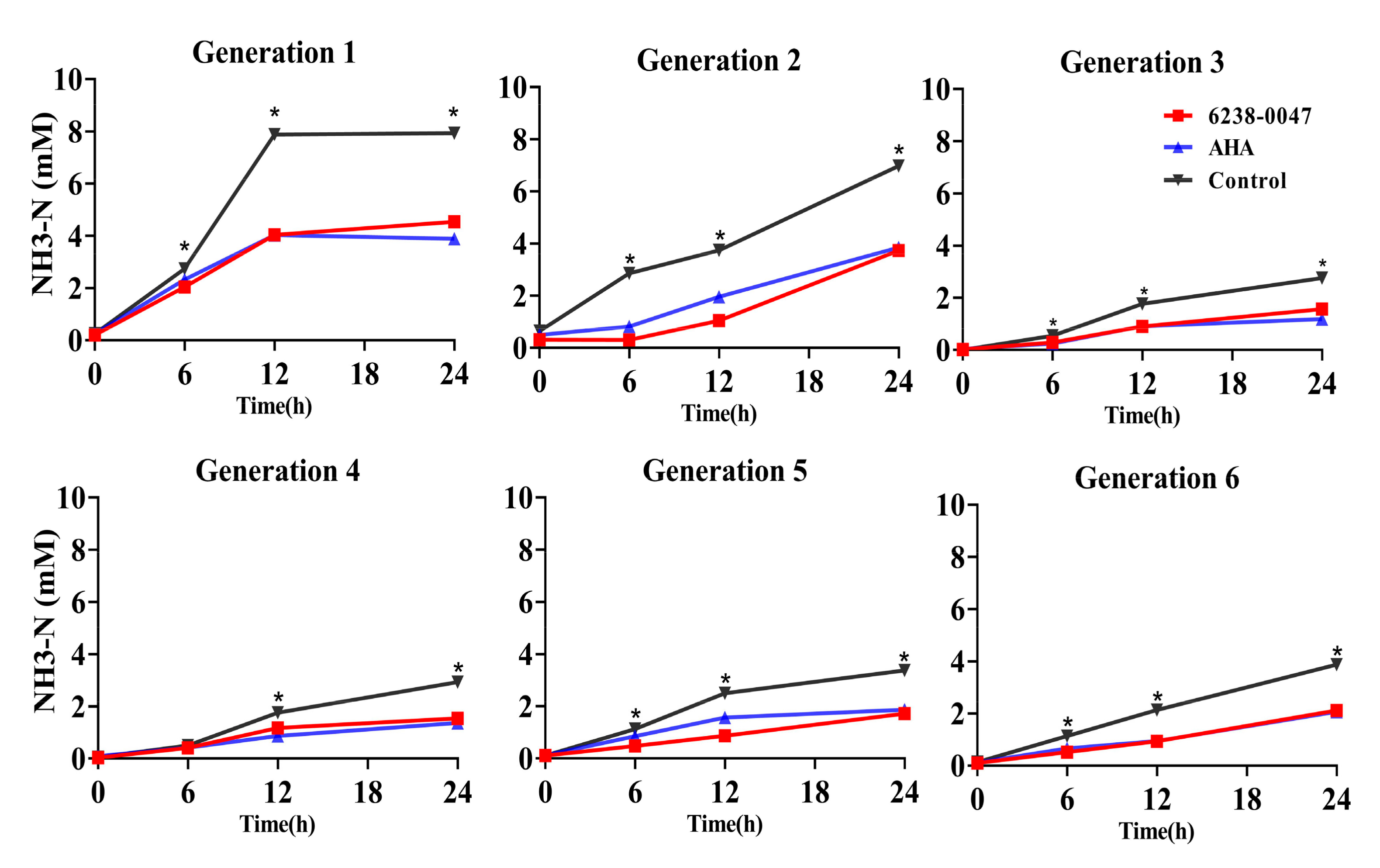
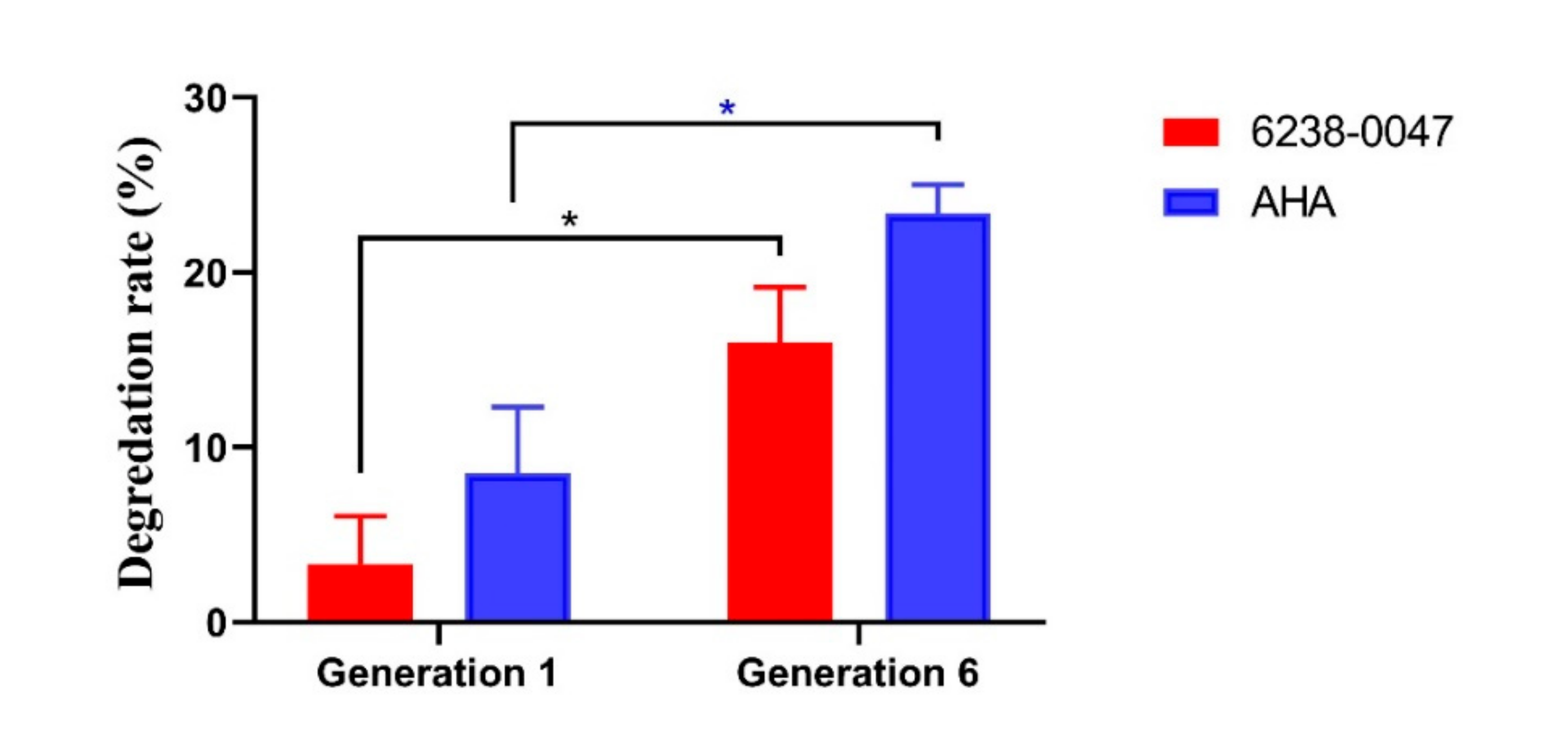
2.4. Inhibition Capacity of the Compound on Ruminal Microbial Community
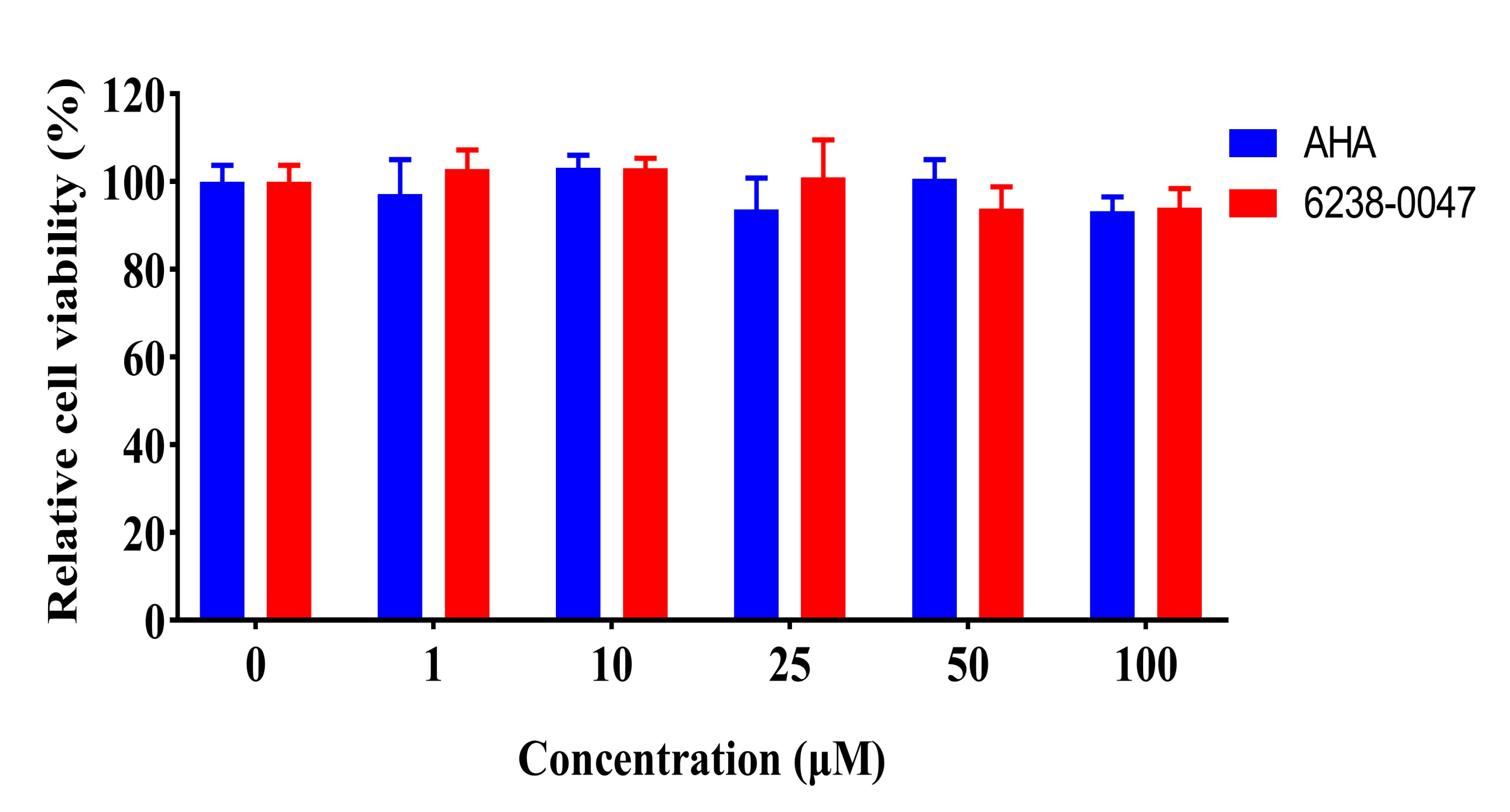
2.5. Toxicity Valuation of Screened Compound
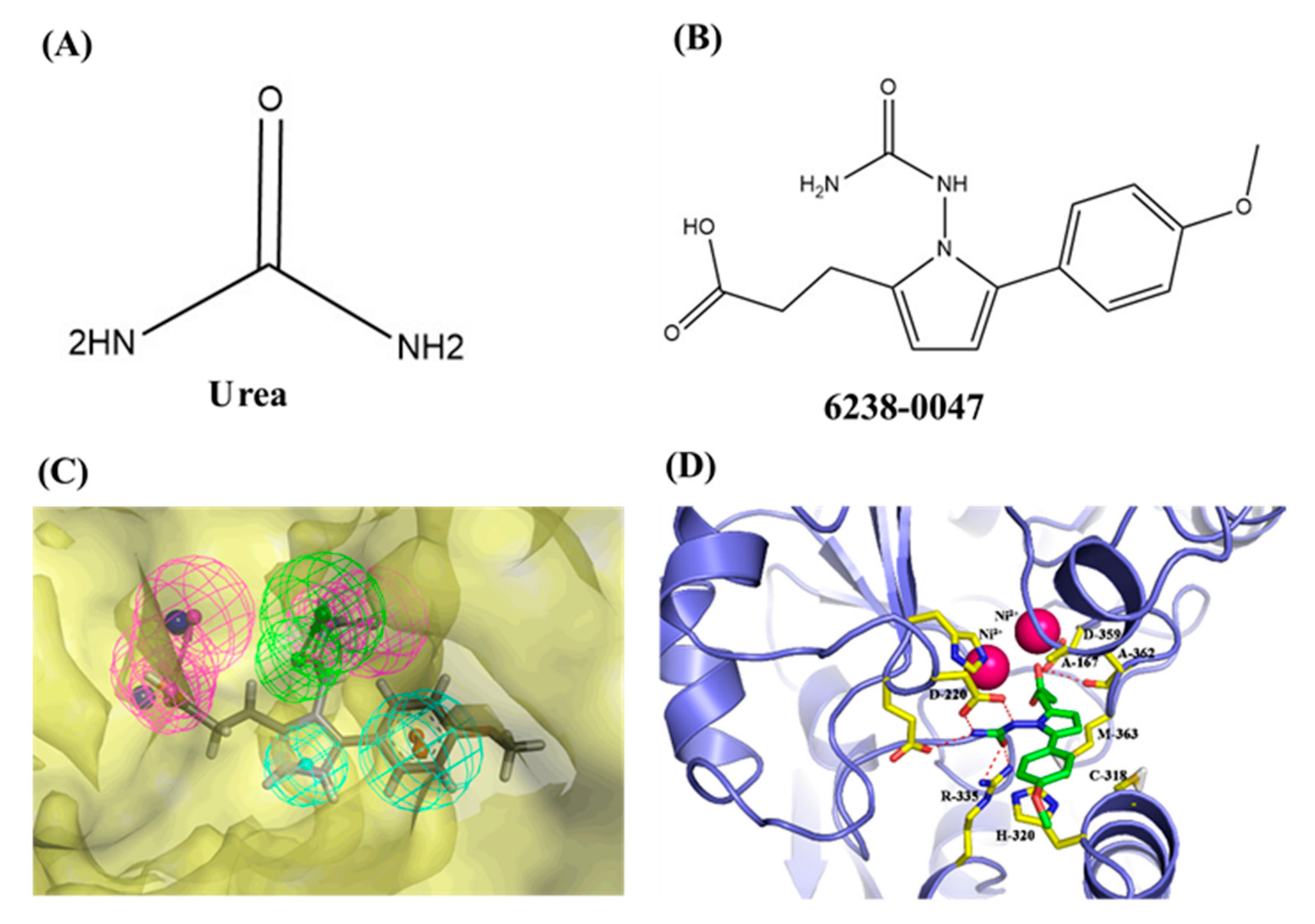
2.6. Chemical Information and Binding Mode
3. Discussion
4. Materials and Methods
4.1. Homology Modeling of Rumen Metagenomic Urease
4.2. Molecular Docking-Based Virtual Screening for Urease Inhibitors
4.3. Ruminal Microbial Urease Preparation
4.4. Urease Activity Inhibition and Kinetic Characterization
4.5. Evaluation of Screened Compound by Ruminal Microbial Fermentation
4.6. Toxicity Evaluation of Screened Compound Using a Caco-2 Cell Culture
4.7. Visulization of the Docking Position of the Screened Compound against Urease
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Alfano, M.; Cavazza, C. Structure, function, and biosynthesis of nickel-dependent enzymes. Protein Sci. 2020, 29, 1071–1089. [Google Scholar] [CrossRef]
- Di, J.; Zhao, S.; Zheng, N.; Bu, D.; Beckers, Y.; Denman, S.E.; McSweeney, C.S.; Wang, J. Differences in Ureolytic Bacterial Composition between the Rumen Digesta and Rumen Wall Based on ureC Gene Classification. Front. Microbiol. 2017, 8, 385. [Google Scholar] [CrossRef] [Green Version]
- Rego, Y.F.; Queiroz, M.P.; Brito, T.O.; Carvalho, P.G.; de Queiroz, V.T.; de Fátima, Â.; Macedo, F. A review on the development of urease inhibitors as antimicrobial agents against pathogenic bacteria. J. Adv. Res. 2018, 13, 69–100. [Google Scholar] [CrossRef]
- Kertz, A.F. Review: Urea Feeding to Dairy Cattle: A Historical Perspective and Review. Prof. Anim. Sci. 2010, 26, 257–272. [Google Scholar] [CrossRef]
- Patra, A.K.; Aschenbach, J.R. Ureases in the gastrointestinal tracts of ruminant and monogastric animals and their implication in urea-N/ammonia metabolism: A review. J. Adv. Res. 2018, 13, 39–50. [Google Scholar] [CrossRef]
- Williams, J.J.; Rodman, J.S.; Peterson, C.M. A Randomized Double-Blind Study of Acetohydroxamic Acid in Struvite Nephrolithiasis. N. Engl. J. Med. 1984, 311, 760–764. [Google Scholar] [CrossRef]
- Thomas, B.; Tolley, D. Concurrent urinary tract infection and stone disease: Pathogenesis, diagnosis and management. Nat. Clin. Pract. Urol. 2008, 5, 668–675. [Google Scholar] [CrossRef]
- Jones, G.A. Influence of acetohydroxamic acid on some activities in vitro of the rumen microbiota. Can. J. Microbiol. 1968, 14, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.C.-L.; Jones, G.A. Effect of acetohydroxamic acid on growth and volatile fatty acid production by rumen bacteria. Can. J. Microbiol. 1973, 19, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Kataria, R.; Khatkar, A. Lead Molecules for Targeted Urease Inhibition: An Updated Review from 2010–2018. Curr. Protein Pept. Sci. 2019, 20, 1158–1188. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, X.; Lin, X. A Review on Applications of Computational Methods in Drug Screening and Design. Molecules 2020, 25, 1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosikowska, P.; Berlicki, L. Urease inhibitors as potential drugs for gastric and urinary tract infections: A patent review. Expert Opin. Ther. Pat. 2011, 21, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Al-Rashida, M.; Uroos, M.; Qazi, S.U.; Naz, S.; Ishtiaq, M.; Khan, K.M. A patent update on therapeutic applications of urease inhibitors (2012–2018). Expert Opin. Ther. Pat. 2019, 29, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B.; Musiani, F.; Benini, S.; Ciurli, S. Chemistry of Ni2+ in urease: Sensing, trafficking, and catalysis. Acc. Chem. Res. 2011, 44, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Macomber, L.; Minkara, M.S.; Hausinger, R.P.; Merz, K.M., Jr. Reduction of urease activity by interaction with the flap covering the active site. J. Chem. Inf. Model. 2015, 55, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, S.; He, Y.; Zheng, N.; Yan, X.; Wang, J. Substitution of residues in UreG to investigate UreE interactions and nickel binding in a predominant urease gene cluster from the ruminal metagenome. Int. J. Boil. Macromol. 2020. [Google Scholar] [CrossRef]
- Rauf, A.; Shahzad, S.A.; Bajda, M.; Yar, M.; Ahmed, F.; Hussain, N.; Akhtar, M.N.; Khan, A.; Jończyk, J. Design and synthesis of new barbituric- and thiobarbituric acid derivatives as potent urease inhibitors: Structure activity relationship and molecular modeling studies. Bioorgan. Med. Chem. 2015, 23, 6049–6058. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.; Ur-Rehman, S.; Channar, P.A.; Larik, F.A.; Abbas, Q.; Hassan, M.; Raza, H.; Seo, S.-Y. Jack Bean Urease Inhibitors, and Antioxidant Activity Based on Palmitic acid Derived 1-acyl-3- Arylthioureas: Synthesis, Kinetic Mechanism and Molecular Docking Studies. Drug Res. 2017, 67, 596–605. [Google Scholar] [CrossRef]
- Shi, W.-K.; Deng, R.-C.; Wang, P.; Yue, Q.-Q.; Liu, Q.; Ding, K.-L.; Yang, M.-H.; Zhang, H.-Y.; Gong, S.-H.; Deng, M.; et al. 3-Arylpropionylhydroxamic acid derivatives as Helicobacter pylori urease inhibitors: Synthesis, molecular docking and biological evaluation. Bioorganic Med. Chem. 2016, 24, 4519–4527. [Google Scholar] [CrossRef]
- Xiao, Z.-P.; Peng, Z.-Y.; Dong, J.-J.; He, J.; Ouyang, H.; Feng, Y.-T.; Lu, C.-L.; Lin, W.-Q.; Wang, J.; Xiang, Y.-P.; et al. Synthesis, structure–activity relationship analysis and kinetics study of reductive derivatives of flavonoids as Helicobacter pylori urease inhibitors. Eur. J. Med. Chem. 2013, 63, 685–695. [Google Scholar] [CrossRef]
- Krajewska, B.; Zaborska, W. Jack bean urease: The effect of active-site binding inhibitors on the reactivity of enzyme thiol groups. Bioorgan. Chem. 2007, 35, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, M.A.; Shams, S.; Choudhary, M.I.; Lodhi, A.; Ul-Haq, Z.; Jalil, S.; Nawaz, S.A.; Khan, K.M.; Iqbal, S.; Rahman, A.-U. Structural Basis of Binding and Rationale for the Potent Urease Inhibitory Activity of Biscoumarins. BioMed Res. Int. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhatt, S.; Bhattacharyya, K. Enzyme kinetics: A note on negative reaction constants in Lineweaver-Burk plots. arXiv 2017, arXiv:1710.08131. [Google Scholar]
- Murugan, R. Solution to Michaelis–Menten enzyme kinetic equation via undetermined gauge functions: Resolving the nonlinearity of Lineweaver–Burk plot. J. Chem. Phys. 2002, 117, 4178–4183. [Google Scholar] [CrossRef]
- Ludden, P.A.; Harmon, D.L.; Huntington, G.B.; Larson, B.T.; Axe, D.E. Influence of the novel urease inhibitor N-(n-butyl) thiophosphoric triamide on ruminant nitrogen metabolism: II. Ruminal nitrogen metabolism, diet digestibility, and nitrogen balance in lambs. J. Anim. Sci. 2000, 78, 188–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabri, E.; Carr, M.; Hausinger, R.P.; Karplus, P. The crystal structure of urease from Klebsiella aerogenes. Science 1995, 268, 998–1004. [Google Scholar] [CrossRef]
- Jabri, E.; Karplus, P. Structures of the Klebsiella aerogenes Urease Apoenzyme and Two Active-Site Mutants. Biochemistry 1996, 35, 10616–10626. [Google Scholar] [CrossRef]
- Hanif, M.; Kanwal, F.; Rafiq, M.; Hassan, M.; Mustaqeem, M.; Seo, S.Y.; Zhang, Y.; Lu, C.; Chen, T.; Saleem, M. Symmetrical Heterocyclic Cage Skeleton: Synthesis, Urease Inhibition Activity, Kinetic Mechanistic Insight, and Molecular Docking Analyses. Molecules 2019, 24, 312. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, M.T.; Khan, K.M.; Arshia; Khan, A.; Arshad, F.; Fatima, B.; Choudhary, M.I.; Syed, N.; Moin, S.T. Syntheses of 4,6-dihydroxypyrimidine diones, their urease inhibition, in vitro, in silico, and kinetic studies. Bioorgan. Chem. 2017, 75, 317–331. [Google Scholar] [CrossRef]
- Saeed, A.; Khan, M.S.; Rafique, H.; Shahid, M.; Iqbal, J. Design, synthesis, molecular docking studies and in vitro screening of ethyl 4-(3-benzoylthioureido) benzoates as urease inhibitors. Bioorgan. Chem. 2014, 52, 1–7. [Google Scholar] [CrossRef]
- Benini, S.; Kosikowska, P.; Cianci, M.; Mazzei, L.; Vara, A.G.; Berlicki, Ł.; Ciurli, S. The crystal structure of Sporosarcina pasteurii urease in a complex with citrate provides new hints for inhibitor design. JBIC J. Boil. Inorg. Chem. 2013, 18, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.P.; Miller, I.B.R.; Roitberg, A.E.; Merz, J.K.M. Wide-Open Flaps Are Key to Urease Activity. J. Am. Chem. Soc. 2012, 134, 9934–9937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; Beer, T.A.P.D.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2012, 64, 4–17. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Tian, H.; Zhou, Z.; Shui, G.; Lam, S.M. Extensive Profiling of Polyphenols from Two Trollius Species Using a Combination of Untargeted and Targeted Approaches. Metabolites 2020, 10, 119. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | ChemDiv ID | Total Score | Structure |
|---|---|---|---|
| 1 | 6238_0047 | 10.0992 |  |
| 2 | G279_1187 | 9.7817 |  |
| 3 | G819_0203 | 9.7376 |  |
| 4 | E734_1898 | 9.703 |  |
| 5 | F046_0043 | 9.2562 |  |
| 6 | E551_0137 | 9.2236 |  |
| 7 | D146_0198 | 9.198 |  |
| 8 | D481_1476 | 9.0675 |  |
| 9 | S576_0588 | 9.0028 |  |
| 10 | F046_0013 | 8.9094 |  |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Li, M.; Zhang, X.; Zheng, N.; Zhao, S.; Wang, J. A Novel Urease Inhibitor of Ruminal Microbiota Screened through Molecular Docking. Int. J. Mol. Sci. 2020, 21, 6006. https://doi.org/10.3390/ijms21176006
Zhang Z, Li M, Zhang X, Zheng N, Zhao S, Wang J. A Novel Urease Inhibitor of Ruminal Microbiota Screened through Molecular Docking. International Journal of Molecular Sciences. 2020; 21(17):6006. https://doi.org/10.3390/ijms21176006
Chicago/Turabian StyleZhang, Zhenyu, Ming Li, Xiaoyin Zhang, Nan Zheng, Shengguo Zhao, and Jiaqi Wang. 2020. "A Novel Urease Inhibitor of Ruminal Microbiota Screened through Molecular Docking" International Journal of Molecular Sciences 21, no. 17: 6006. https://doi.org/10.3390/ijms21176006
APA StyleZhang, Z., Li, M., Zhang, X., Zheng, N., Zhao, S., & Wang, J. (2020). A Novel Urease Inhibitor of Ruminal Microbiota Screened through Molecular Docking. International Journal of Molecular Sciences, 21(17), 6006. https://doi.org/10.3390/ijms21176006