Adipose Tissue Fibrosis: Mechanisms, Models, and Importance


Abstract
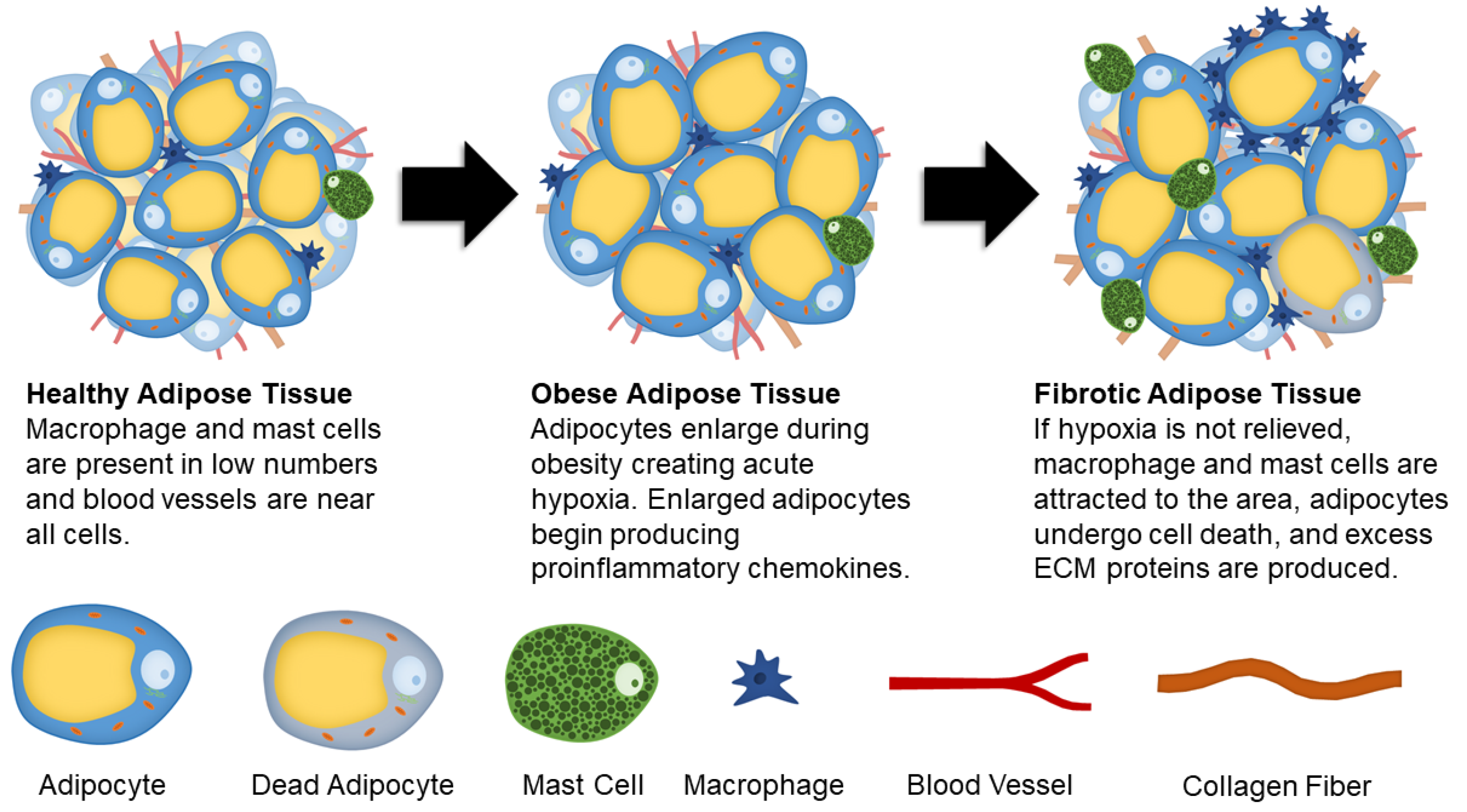
:1. Introduction
2. Mechanisms
3. In Vitro Adipose Tissue Fibrosis Modeling
3.1. 2D Models
3.2. Three-Dimensional Models
Biomaterials Used for 3D Adipose Tissue Culture
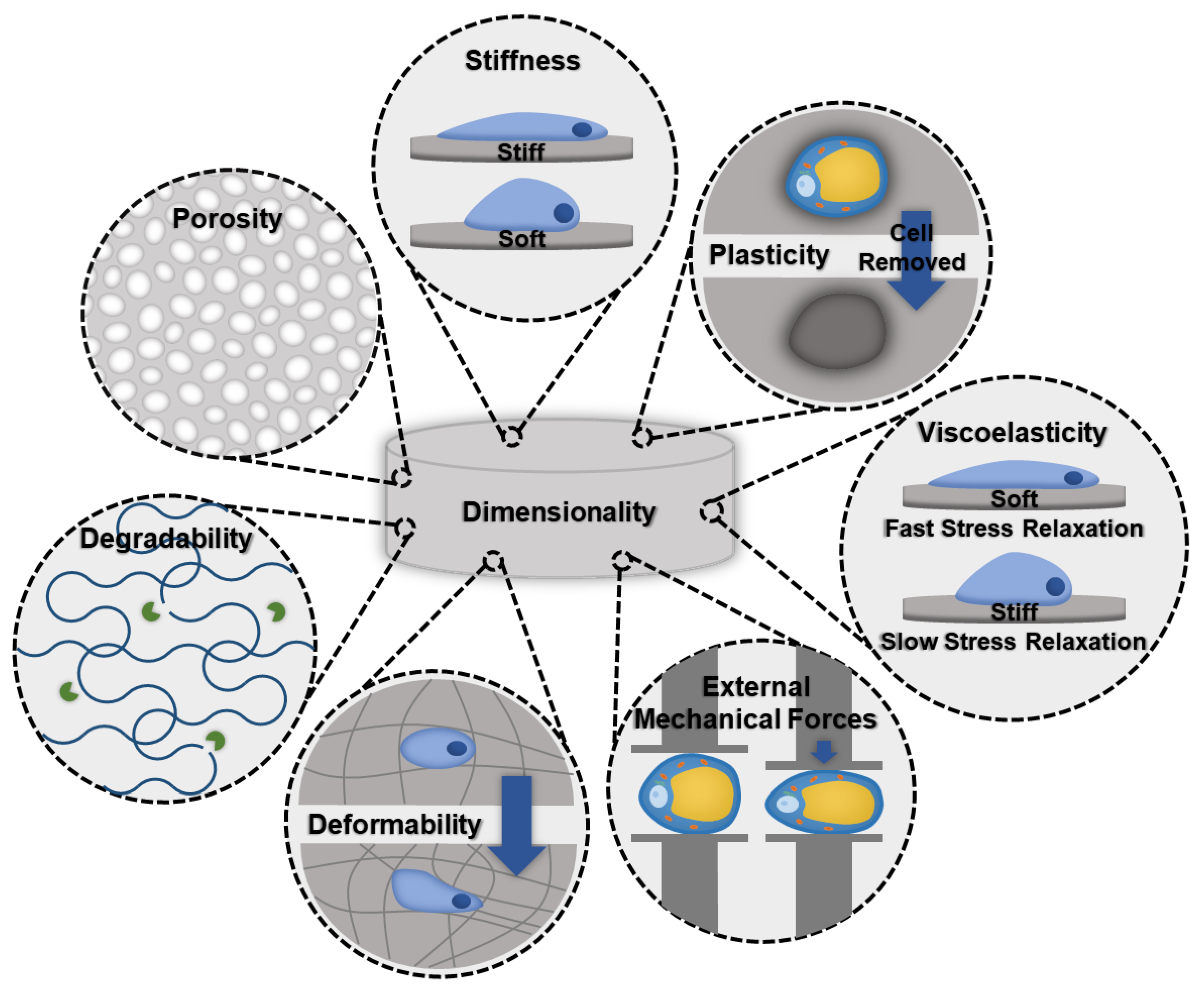
3.3. Biomaterial Considerations for In Vitro Modeling
- Stiffness: Stiffness is an intrinsic property of a material and is defined as the resistance of an elastic material to deformation by an applied force. Adipocytes and ASCs are mechanosensitive and mechanoresponsive [100,101]. In the body, mechanical forces are balanced internally by the cytoskeleton and externally through the ECM. Scaffolding materials act as the ECM in in vitro models, therefore, the stiffness should be carefully considered as it will influence the cellular cytoskeleton and phenotype [100]. Scaffold stiffness can be controlled by adding other polymers or additives [81]. A recent study found that stiffer biomaterials triggered fibrotic traits in adipocytes, such as increased profibrotic gene expression and ECM deposition [81]. Additionally, stiffer matrices were found to promote osteogenic differentiation of ASCs, while softer matrices promoted adipogenic commitment [101].
- Viscoelasticity: Under deformation, a viscoelastic biomaterial will exhibit both elastic and viscous behavior making it a time-dependent response. Similar to stiffness, viscoelasticity is an intrinsic material property. The stress response of the material will vary based on strain and history of deformation and allows viscoelastic materials to exhibit stress relaxation, hysteresis, and creep. Soft tissues, like adipose tissue, are composed of solids and liquids and naturally behave like viscoelastic materials. An increase in collagen content results in a greater elastic component to the adipose tissue, and therefore fibrotic tissues would exhibit a decrease in the time-dependent viscous behavior.
- Degradability: A biomaterials degradability is also an intrinsic trait. It is dependent on the molecular properties of the biomaterial. Factors, such as cross-linking, can decrease the degradability. As fibrosis involves significant deposition of ECM proteins, degradability should be considered to ensure that cells have enough time and the ability to remodel their environment before the scaffolding degrades.
- Dimensionality: Dimensionality is an extrinsic material property that defines the number of dimensions an object occupies. To make the most accurate model for adipose tissue fibrosis a 3-dimensional model is necessary. Typically, growing cells in 3D constructs is more difficult than 2D, but due to adipocytes’ unique buoyancy issues, and the fact that 2D cultures results in different morphological (multilocular lipid droplets as opposed to the single unilocular lipid droplet observed in vivo) and functional adipocytes, 3D culture is often pursued as a more physiologically relevant option. In particular, the largest difference is in cell volume; 2D culture results in adipocytes with less than 20% the total cell volume of lean subcutaneous adipocytes and less than 3% the total cell volume of obese subcutaneous adipocytes [102,103,104,105,106].
- Deformability: Deformability is an intrinsic material property that is defined as the ability for the material to change shape. The goal of designing a biomaterial to model adipose tissue fibrosis is to encapsulate adipocytes and the stromal vascular fraction (SVF) in a scaffold with properties similar to the ECM found in adipose tissues. One method is to begin with a highly deformable biomaterial that has similar properties to healthy adipose tissue and allow cells to remodel their environment by triggering fibrosis. Alternatively, by beginning with a biomaterial scaffold that has properties of fibrotic adipose tissue, and a low degree of deformability, fibrotic adipose tissue can be modeled. Depending on the experiment and the end goal, each method offers unique advantages.
- Plasticity: Plasticity is an intrinsic material property that is related to stiffness and viscoelasticity. Several studies have found that cells are able to plastically remodel certain biomaterial environments, such as collagen and fibrin gels [107,108,109]. Cells align and compact the fibers around them to the point where, when the cells are removed, voids remain [110]. Healthy adipose tissue is a dynamic organ that is remodeled constantly to allow fluctuations in adipocyte size to meet energy storage and demand needs. However, adipose tissue fibrosis limits the capacity of adipose tissue to remodel [111]. The addition of crosslinks can lower the accessibility of cells to remodel the biomaterial [112].
- Porosity: Porosity is an extrinsic property that is defined as the percentage of void space in a material. Generally, biomaterial scaffolds used to culture adipose tissue have high porosities (>90%) [83,84] to accommodate the large size of lipid-laden adipocytes. However, pore sizes vary considerably based on the method of formation (135–633 µm) [83,93]. For example, gas foaming results in a larger range of pore sizes and is more difficult to control compared to salt leaching [113]. Researchers that have used salt leaching to generate their scaffolds have chosen pore sizes ranging from 500–600 µm [93]. Electrospun scaffolds had slightly lower porosity (~88%) and considerably smaller pore sizes (6–70 µm); however, these scaffolds were used with murine ASC cell lines that are smaller and do not contain the large unilocular lipid droplet [89]. Three-dimensional printing can be used to create specific pore sizes but depends heavily on the print resolution.
- Processing: There are many methods of processing biomaterials. Some researchers have used electrospinning as a method of making scaffolds for in vitro adipose tissue modeling [89,90]. Electrospinning was chosen because the resulting scaffold has similar structure to collagen fibers found in the ECM matrix of adipose tissue. However, electrospinning resulted in lower porosity and smaller pore sizes. Three-dimensional bioprinting has been used to model other fibrotic diseases [114]. Printing with cells allows for more complex tissues to be created. Currently, printing vasculature is difficult, which limits the size of the print, as necrotic cores develop if cells do not have access to nutrients in large tissue constructs without vasculature. Three-dimensional printing also is not an option for all biomaterials. Decellularized tissue matrices have been used to study adipose tissue fibrosis [80], with conduits for vasculature to help sustain long term culture. However, decellularized tissue matrices can limit the cell-cell interactions and only represent the final stages of the disease [115]. Hydrogels are useful for tissue culture due to ease of fabrication and can be made using ECM proteins [81]. However, the mechanical properties of hydrogels have lower tunability.
- External Mechanical Forces: Including mechanical forces, such as tensile or compressive strains, is independent of material properties. Applying tensile strains of about 12% to adipocyte cultures resulted in faster accumulation of lipids and larger lipid droplets compared to adipocytes cultured under no external stresses [116]. This could be a useful technique to expedite the formation of a lipid-rich model.
4. Current Methods of Modeling Fibrotic Adipose Tissue In Vivo
Rodent
5. Human Epidemiological Studies
6. Adipose Tissue’s Role in Treating Other Fibrotic Diseases
7. Methods for Evaluating Degree of Fibrosis
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Saely:, C.H.; Geiger, K.; Drexel, H. Brown versus white adipose tissue: A mini-review. Gerontology 2012, 58, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Zimmermann, R.; Oberer, M.; Zechner, R. Lipolysis—A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Prog. Lipid Res. 2011, 50, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Lelliott, C.; Vidal-Puig, A. Lipotoxicity, an imbalance between lipogenesis de novo and fatty acid oxidation. Int. J. Obes. 2004, 28, S22–S28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verboven, K.; Wouters, K.; Gaens, K.; Hansen, D.; Bijnen, M.; Wetzels, S.; Stehouwer, C.D.; Goossens, G.H.; Schalkwijk, C.G.; Blaak, E.E.; et al. Abdominal subcutaneous and visceral adipocyte size, lipolysis and inflammation relate to insulin resistance in male obese humans. Sci. Rep. 2018, 8, 4677. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, B.; Gogg, S.; Hedjazifar, S.; Jenndahl, L.; Hammarstedt, A.; Smith, U. Inflammation and impaired adipogenesis in hypertrophic obesity in man. Am. J. Physiol. Metab. 2009, 297, E999–E1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buechler, C.; Krautbauer, S.; Eisinger, K. Adipose tissue fibrosis. World J. Diabetes 2015, 6, 548–553. [Google Scholar] [CrossRef]
- Mariman, E.C.M.; Wang, P. Adipocyte extracellular matrix composition, dynamics and role in obesity. Cell. Mol. Life Sci. 2010, 67, 1277–1292. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.J.; Barnard, R.J. Effect of diet on fat cell size and hormone-sensitive lipase activity. J. Appl. Physiol. 1999, 87, 227–232. [Google Scholar] [CrossRef]
- Wueest, S.; Rapold, R.A.; Rytka, J.M.; Schoenle, E.J.; Konrad, D. Basal lipolysis, not the degree of insulin resistance, differentiates large from small isolated adipocytes in high-fat fed mice. Diabetologia 2008, 52, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Jocken, J.W.E.; Goossens, G.H.; Van Hees, A.M.J.; Frayn, K.N.; Van Baak, M.; Stegen, J.; Pakbiers, M.T.W.; Saris, W.H.M.; Blaak, E.E. Effect of beta-adrenergic stimulation on whole-body and abdominal subcutaneous adipose tissue lipolysis in lean and obese men. Diabetologia 2007, 51, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Reynisdottir, S.; Ellerfeldt, K.; Wahrenberg, H.; Lithell, H.; Arner, P. Multiple lipolysis defects in the insulin resistance (metabolic) syndrome. J. Clin. Investig. 1994, 93, 2590–2599. [Google Scholar] [CrossRef] [Green Version]
- Campbell, P.J.; Carlson, M.G.; Nurjhan, N. Fat metabolism in human obesity. Am. J. Physiol. Metab. 1994, 266, E600–E605. [Google Scholar] [CrossRef] [PubMed]
- Renes, J.; Bouwman, F.; Noben, J.-P.; Evelo, C.; Robben, J.; Mariman, E. Protein profiling of 3T3-L1 adipocyte differentiation and (tumor necrosis factor alpha-mediated) starvation. Cell. Mol. Life Sci. 2005, 62, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Tordjman, J.; Clément, K.; Scherer, P.E. Fibrosis and adipose tissue dysfunction. Cell Metab. 2013, 18, 470–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawler, H.M.; Underkofler, C.M.; Kern, P.A.; Erickson, C.; Bredbeck, B.; Rasouli, N. Adipose tissue hypoxia, inflammation, and fibrosis in obese insulin-sensitive and obese insulin-resistant subjects. J. Clin. Endocrinol. Metab. 2016, 101, 1422–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchwald, H.; Avidor, Y.; Braunwald, E. Bariatric surgery. A systematic review and meta-analysis. ACC Curr. J. Rev. 2005, 14, 1724–1737. [Google Scholar] [CrossRef]
- Dandona, P.; Aljada, A.; Chaudhuri, A.; Mohanty, P.; Garg, R. Metabolic syndrome: A comprehensive perspective based on interactions between obesity, diabetes, and inflammation. Circulation 2005, 111, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Dankel, S.N.; Svard, J.; Mattha, S.; Claussnitzer, M.; Kloting, N.; Glunk, V.; Fandalyuk, Z.; Grytten, E.; Solsvik, M.H.; Nielsen, H.J.; et al. COL6A3 expression in adipocytes associates with insulin resistance and depends on PPARgamma and adipocyte size. Obesity 2014, 22, 1807–1813. [Google Scholar] [CrossRef]
- Divoux, A.; Tordjman, J.; Lacasa, D.; Veyrie, N.; Hugol, D.; Aissat, A.; Basdevant, A.; Guerre-Millo, M.; Poitou, C.; Zucker, J.D.; et al. Fibrosis in human adipose tissue: Composition, distribution, and link with lipid metabolism and fat mass loss. Diabetes 2010, 59, 2817–2825. [Google Scholar] [CrossRef] [Green Version]
- Lackey, D.E.; Burk, D.H.; Ali, M.R.; Mostaedi, R.; Smith, W.H.; Park, J.; Scherer, P.E.; Seay, S.A.; McCoin, C.S.; Bonaldo, P.; et al. Contributions of adipose tissue architectural and tensile properties toward defining healthy and unhealthy obesity. Am. J. Physiol. Metab. 2014, 306, E233–E246. [Google Scholar] [CrossRef] [Green Version]
- Muir, L.A.; Neeley, C.K.; Meyer, K.A.; Baker, N.A.; Brosius, A.M.; Washabaugh, A.R.; Varban, O.A.; Finks, J.F.; Zamarron, B.F.; Flesher, C.G.; et al. Adipose tissue fibrosis, hypertrophy, and hyperplasia: Correlations with diabetes in human obesity. Obesity 2016, 24, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Park, J.; Gupta, O.T.; Holland, W.L.; Auerbach, P.; Zhang, N.; Marangoni, R.G.; Nicoloro, S.M.; Czech, M.P.; Varga, J.; et al. Endotrophin triggers adipose tissue fibrosis and metabolic dysfunction. Nat. Commun. 2014, 5, 3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, M.; Yao-Borengasser, A.; Unal, R.; Rasouli, N.; Gurley, C.M.; Zhu, B.; Peterson, C.A.; Kern, P.A. Adipose tissue macrophages in insulin-resistant subjects are associated with collagen VI and fibrosis and demonstrate alternative activation. Am. J. Physiol. Metab. 2010, 299, E1016–E1027. [Google Scholar] [CrossRef]
- Divoux, A.; Clement, K. Architecture and the extracellular matrix: The still unappreciated components of the adipose tissue. Obes. Rev. 2011, 12, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Datta, R.; Podolsky, M.J.; Atabai, K. Fat fibrosis: Friend or foe? JCI Insight 2018, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.; Löffler, M.; Bilban, M.; Reimers, M.; Kadl, A.; Todoric, J.; Zeyda, M.; Geyeregger, R.; Schreiner, M.; Weichhart, T.; et al. Prevention of high-fat diet-induced adipose tissue remodeling in obese diabetic mice by n-3 polyunsaturated fatty acids. Int. J. Obes. 2006, 31, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Spencer, M.; Unal, R.; Zhu, B.; Rasouli, N.; McGehee, R.E.; Peterson, C.A.; Kern, P.A. Adipose tissue extracellular matrix and vascular abnormalities in obesity and insulin resistance. J. Clin. Endocrinol. Metab. 2011, 96, E1990–E1998. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.V.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic dysregulation and adipose tissue fibrosis: Role of collagen VI. Mol. Cell. Biol. 2008, 29, 1575–1591. [Google Scholar] [CrossRef] [Green Version]
- Marcelin, G.; Silveira, A.L.M.; Martins, L.B.; Ferreira, A.V.; Clément, K. Deciphering the cellular interplays underlying obesity-induced adipose tissue fibrosis. J. Clin. Investig. 2019, 129, 4032–4040. [Google Scholar] [CrossRef]
- Giordano, A.; Murano, I.; Mondini, E.; Perugini, J.; Smorlesi, A.; Severi, I.; Barazzoni, R.; Scherer, P.E.; Cinti, S. Obese adipocytes show ultrastructural features of stressed cells and die of pyroptosis. J. Lipid Res. 2013, 54, 2423–2436. [Google Scholar] [CrossRef] [Green Version]
- Jernås, M.; Palming, J.; Sjöholm, K.; Jennische, E.; Svensson, P.A.; Gabrielsson, B.G.; Levin, M.; Sjögren, A.; Rudemo, M.; Lystig, T.; et al. Separation of human adipocytes by size: Hypertrophic fat cells display distinct gene expression. FASEB J. 2006, 20, 1540–1542. [Google Scholar] [CrossRef] [PubMed]
- Sartipy, P.; Loskutoff, D.J. Monocyte chemoattractant protein 1 in obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 7265–7270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.E.; Ko, M.S.; Yun, J.Y.; Kim, M.O.; Kim, J.H.; Park, H.S.; Kim, A.R.; Kim, H.J.; Kim, B.J.; Ahn, Y.E.; et al. Nitric oxide produced by macrophages inhibits adipocyte differentiation and promotes profibrogenic responses in preadipocytes to induce adipose tissue fibrosis. Diabetes 2016, 65, 2516–2528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trayhurn, P.; Wang, B.; Wood, I.S. Hypoxia in adipose tissue: A basis for the dysregulation of tissue function in obesity? Br. J. Nutr. 2008, 100, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415. [Google Scholar] [CrossRef]
- Hotamisligil, G.; Shargill, N.; Spiegelman, B. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- Fried, S.K.; Bunkin, D.A.; Greenberg, A.S. Omental and subcutaneous adipose tissues of obese subjects release interleukin-6: Depot difference and regulation by glucocorticoid. J. Clin. Endocrinol. Metab. 1998, 83, 847–850. [Google Scholar] [CrossRef]
- Perreault, M.; Marette, A. Targeted disruption of inducible nitric oxide synthase protects against obesity-linked insulin resistance in muscle. Nat. Med. 2001, 7, 1138–1143. [Google Scholar] [CrossRef]
- Visser, M.; Bouter, L.M.; McQuillan, G.M.; Wener, M.H.; Harris, T.B. Elevated C-reactive protein levels in overweight and obese adults. JAMA 1999, 282, 2131–2135. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.E.C.; Rabhi, N.; Orofino, J.; Gamini, R.; Perissi, V.; Vernochet, C.; Farmer, S.R. The adipocyte acquires a fibroblast-like transcriptional signature in response to a high fat diet. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Marcelin, G.; Ferreira, A.; Liu, Y.; Atlan, M.; Aron-Wisnewsky, J.; Pelloux, V.; Botbol, Y.; Ambrosini, M.; Fradet, M.; Rouault, C.; et al. A PDGFRalpha-mediated switch toward CD9(high) adipocyte progenitors controls obesity-induced adipose tissue fibrosis. Cell Metab. 2017, 25, 673–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwayama, T.; Steele, C.; Yao, L.; Dozmorov, M.G.; Karamichos, D.; Wren, J.D.; Olson, L.E. PDGFRalpha signaling drives adipose tissue fibrosis by targeting progenitor cell plasticity. Genes Dev. 2015, 29, 1106–1119. [Google Scholar] [CrossRef] [Green Version]
- Engin, A. Adipose tissue hypoxia in obesity and its impact on preadipocytes and macrophages: Hypoxia hypothesis. Adv. Exp. Med. Biol. 2017, 960, 305–326. [Google Scholar] [CrossRef]
- Cao, Y. Angiogenesis modulates adipogenesis and obesity. J. Clin. Investig. 2007, 117, 2362–2368. [Google Scholar] [CrossRef] [Green Version]
- Pellegrinelli, V.; Rouault, C.; Veyrie, N.; Clement, K.; Lacasa, D. Endothelial cells from visceral adipose tissue disrupt adipocyte functions in a three-dimensional setting: Partial rescue by angiopoietin-1. Diabetes 2013, 63, 535–549. [Google Scholar] [CrossRef] [Green Version]
- Crewe, C.; An, Y.A.; Scherer, P.E. The ominous triad of adipose tissue dysfunction: Inflammation, fibrosis, and impaired angiogenesis. J. Clin. Investig. 2017, 127, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Tanaka, M.; Ikeda, K.; Suganami, T.; Komiya, C.; Ochi, K.; Shirakawa, I.; Hamaguchi, M.; Nishimura, S.; Manabe, I.; Matsuda, T.; et al. Macrophage-inducible C-type lectin underlies obesity-induced adipose tissue fibrosis. Nat. Commun. 2014, 5, 4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murano, I.; Barbatelli, G.; Parisani, V.; Latini, C.; Muzzonigro, G.; Castellucci, M.; Cinti, S. Dead adipocytes, detected as crown-like structures, are prevalent in visceral fat depots of genetically obese mice. J. Lipid Res. 2008, 49, 1562–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keophiphath, M.; Achard, V.; Henegar, C.; Rouault, C.; Clément, K.; Lacasa, D. Macrophage-secreted factors promote a profibrotic phenotype in human preadipocytes. Mol. Endocrinol. 2009, 23, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Chun, T.H.; Kang, L. Adipose extracellular matrix remodelling in obesity and insulin resistance. Biochem. Pharmacol. 2016, 119, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopps, E.; Canino, B.; Montana, M.; Calandrino, V.; Urso, C.; Presti, R.L.; Caimi, G. Gelatinases and their tissue inhibitors in a group of subjects with obstructive sleep apnea syndrome. Clin. Hemorheol. Microcirc. 2016, 62, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, G.; Barchuk, M.; Miksztowicz, V. Behavior of metalloproteinases in adipose tissue, liver and arterial wall: An update of extracellular matrix remodeling. Cells 2019, 8, 158. [Google Scholar] [CrossRef] [Green Version]
- Gurung, P.; Moussa, K.; Adams-Huet, B.; Devaraj, S.; Jialal, I. Increased mast cell abundance in adipose tissue of metabolic syndrome: Relevance to the proinflammatory state and increased adipose tissue fibrosis. Am. J. Physiol. Metab. 2019, 316, E504–E509. [Google Scholar] [CrossRef]
- Hirai, S.; Ohyane, C.; Kim, Y.I.; Lin, S.; Goto, T.; Takahashi, N.; Kim, C.S.; Kang, J.; Yu, R.; Kawada, T. Involvement of mast cells in adipose tissue fibrosis. Am. J. Physiol. Metab. 2014, 306, E247–E255. [Google Scholar] [CrossRef] [Green Version]
- Frungieri, M.B.; Weidinger, S.; Meineke, V.; Köhn, F.M.; Mayerhofer, A. Proliferative action of mast-cell tryptase is mediated by PAR2, COX2, prostaglandins, and PPAR: Possible relevance to human fibrotic disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 15072–15077. [Google Scholar] [CrossRef] [Green Version]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Divoux, A.; Moutel, S.; Poitou, C.; Lacasa, D.; Veyrie, N.; Aissat, A.; Arock, M.; Guerre-Millo, M.; Clément, K. Mast cells in human adipose tissue: Link with morbid obesity, inflammatory status, and Diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E1677–E1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajbhandari, P.; Arneson, D.; Hart, S.K.; Ahn, I.S.; Diamante, G.; Santos, L.C.; Zaghari, N.; Feng, A.C.; Thomas, B.J.; Vergnes, L.; et al. Single cell analysis reveals immune cell–adipocyte crosstalk regulating the transcription of thermogenic adipocytes. eLife 2019, 8, 8. [Google Scholar] [CrossRef]
- Merrick, D.; Sakers, A.; Irgebay, Z.; Okada, C.; Calvert, C.; Morley, M.; Percec, I.; Seale, P. Identification of a mesenchymal progenitor cell hierarchy in adipose tissue. Science 2019, 364, eaav2501. [Google Scholar] [CrossRef] [PubMed]
- Schwalie, P.C.; Dong, H.; Zachara, M.; Russeil, J.; Alpern, D.; Akchiche, N.; Caprara, C.; Sun, W.; Schlaudraff, K.U.; Soldati, G.; et al. A stromal cell population that inhibits adipogenesis in mammalian fat depots. Nature 2018, 559, 103–108. [Google Scholar] [CrossRef]
- Hepler, C.; Shan, B.; Zhang, Q.; Henry, G.H.; Shao, M.; Vishvanath, L.; Ghaben, A.L.; Mobley, A.B.; Strand, D.W.; Hon, G.C.; et al. Identification of functionally distinct fibro-inflammatory and adipogenic stromal subpopulations in visceral adipose tissue of adult mice. eLife 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Burl, R.B.; Ramseyer, V.D.; Rondini, E.A.; Pique-Regi, R.; Lee, Y.H.; Granneman, J.G. Deconstructing adipogenesis induced by beta3-adrenergic receptor activation with single-cell expression profiling. Cell Metab. 2018, 28, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Vijay, J.; Gauthier, M.F.; Biswell, R.L.; Louiselle, D.A.; Johnston, J.; Cheung, W.A.; Belden, B.; Pramatarova, A.; Biertho, L.; Gibson, M.; et al. Single-cell analysis of human adipose tissue identifies depot- and disease-specific cell types. Nat. Metab. 2019, 2, 97–109. [Google Scholar] [CrossRef]
- Cho, D.S.; Lee, B.; Doles, J.D. Refining the adipose progenitor cell landscape in healthy and obese visceral adipose tissue using single-cell gene expression profiling. Life Sci. Alliance 2019, 2, e201900561. [Google Scholar] [CrossRef]
- Zhang, H.; Kumar, S.; Barnett, A.H.; Eggo, M.C. Ceiling culture of mature human adipocytes: Use in studies of adipocyte functions. J. Endocrinol. 2000, 164, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sugihara, H.; Yonemitsu, N.; Miyabara, S.; Toda, S. Proliferation of unilocular fat cells in the primary culture. J. Lipid Res. 1987, 28, 1038–1045. [Google Scholar]
- Sugihara, H.; Yonemitsu, N.; Miyabara, S.; Yun, K. Primary cultures of unilocular fat cells: Characteristics of growth in vitro and changes in differentiation properties. Differentiation 1986, 31, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.S.; Loftus, T.M.; Mandrup, S.; Lane, M.D. Adipocyte differentiation and leptin expression. Annu. Rev. Cell Dev. Biol. 1997, 13, 231–259. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Song, K.; Ahn, K.S. Alantolactone improves palmitate-induced glucose intolerance and inflammation in both lean and obese states in vitro: Adipocyte and adipocyte-macrophage co-culture system. Int. Immunopharmacol. 2017, 49, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.F.; Craig, C.M.; Tolentino, L.L.; Choi, O.; Morton, J.; Rivas, H.; Cushman, S.W.; Engleman, E.G.; McLaughlin, T. Adipose tissue macrophages impair preadipocyte differentiation in humans. PLoS ONE 2017, 12, e0170728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suganami, T.; Nishida, J.; Ogawa, Y. A paracrine loop between adipocytes and macrophages aggravates inflammatory changes. Arter. Thromb. Vasc. Biol. 2005, 25, 2062–2068. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Soga, Y.; Iwamoto, Y.; Yoshizawa, S.; Iwata, H.; Kokeguchi, S.; Takashiba, S.; Nishimura, F. Macrophage-adipocyte interaction: Marked interleukin-6 production by Lipopolysaccharide**. Obesity 2007, 15, 2549–2552. [Google Scholar] [CrossRef]
- Lacasa, D.; Taleb, S.; Keophiphath, M.; Miranville, A.; Clément, K. Macrophage-secreted factors impair human adipogenesis: Involvement of proinflammatory state in preadipocytes. Endocrinology 2007, 148, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Isakson, P.; Hammarstedt, A.; Gustafson, B.; Smith, U. Impaired preadipocyte differentiation in human abdominal obesity. Diabetes 2009, 58, 1550–1557. [Google Scholar] [CrossRef] [Green Version]
- Chabot, K.; Gauthier, M.S.; Garneau, P.; Rabasa-Lhoret, R. Evolution of subcutaneous adipose tissue fibrosis after bariatric surgery. Diabetes Metab. 2017, 43, 125–133. [Google Scholar] [CrossRef]
- Baker, N.A.; Muir, L.A.; Washabaugh, A.R.; Neeley, C.K.; Chen, Y.-P.; Flesher, C.G.; Vorwald, J.; Finks, J.F.; Ghaferi, A.A.; Mulholland, M.W.; et al. Diabetes-specific regulation of adipocyte metabolism by the adipose tissue extracellular matrix. J. Clin. Endocrinol. Metab. 2017, 102, 1032–1043. [Google Scholar] [CrossRef] [Green Version]
- Di Caprio, N.; Bellas, E. Collagen stiffness and architecture regulate fibrotic gene expression in engineered adipose tissue. Adv. Biosyst. 2020, 4, e1900286. [Google Scholar] [CrossRef] [PubMed]
- Rajangam, T.; Park, M.H.; Kim, S.H. 3D Human adipose-derived stem cell clusters as a model for in vitro fibrosis. Tissue Eng. Part. C Methods 2016, 22, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Patrick, C.W.; Chauvin, P.; Hobley, J.; Reece, G. Preadipocyte seeded PLGA scaffolds for adipose tissue engineering. Tissue Eng. 1999, 5, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Patrick, C.W.; Zheng, B.; Johnston, C.; Reece, G.P. Long-term implantation of preadipocyte-seeded PLGA scaffolds. Tissue Eng. 2002, 8, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Cha, S.M.; Lee, Y.Y.; Kwon, S.W.; Park, C.; Kim, M. Adipogenic differentiation of adipose tissue derived adult stem cells in nude mouse. Biochem. Biophys. Res. Commun. 2006, 345, 631–637. [Google Scholar] [CrossRef]
- Neubauer, M.; Hacker, M.; Bauer-Kreisel, P.; Weiser, B.; Fischbach, C.; Schulz, M.B.; Goepferich, A.; Blunk, T. Adipose tissue engineering based on mesenchymal stem cells and basic fibroblast growth factor in vitro. Tissue Eng. 2005, 11, 1840–1851. [Google Scholar] [CrossRef]
- Halbleib, M.; Skurk, T.; De Luca, C.; Von Heimburg, D.; Hauner, H. Tissue engineering of white adipose tissue using hyaluronic acid-based scaffolds. I: In vitro differentiation of human adipocyte precursor cells on scaffolds. Biomaterials 2003, 24, 3125–3132. [Google Scholar] [CrossRef]
- Kessler, L.; Gehrke, S.; Winnefeld, M.; Huber, B.; Hoch, E.; Walter, T.; Wyrwa, R.; Schnabelrauch, M.; Schmidt, M.; Kückelhaus, M.; et al. Methacrylated gelatin/hyaluronan-based hydrogels for soft tissue engineering. J. Tissue Eng. 2017, 8, 2041731417744157. [Google Scholar] [CrossRef]
- Kang, X.; Xie, Y.; Powell, H.M.; Lee, L.J.; Belury, M.A.; Lannutti, J.J.; Kniss, D.A. Adipogenesis of murine embryonic stem cells in a three-dimensional culture system using electrospun polymer scaffolds. Biomaterials 2007, 28, 450–458. [Google Scholar] [CrossRef]
- Brännmark, C.; Paúl, A.; Ribeiro, D.; Magnusson, B.; Brolén, G.; Enejder, A.; Forslöw, A. Increased adipogenesis of human adipose-derived stem cells on polycaprolactone fiber matrices. PLoS ONE 2014, 9, e113620. [Google Scholar] [CrossRef] [Green Version]
- Krontiras, P.; Gatenholm, P.; Hägg, D.A. Adipogenic differentiation of stem cells in three-dimensional porous bacterial nanocellulose scaffolds. J. Biomed. Mater. Res. Part. B Appl. Biomater. 2014, 103, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Volz, A.C.; Hack, L.; Kluger, P.J. A cellulose-based material for vascularized adipose tissue engineering. J. Biomed. Mater. Res. Part. B Appl. Biomater. 2018, 107, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Mauney, J.R.; Nguyen, T.; Gillen, K.M.; Kirker-Head, C.; Gimble, J.M.; Kaplan, D.L. Engineering adipose-like tissue in vitro and in vivo utilizing human bone marrow and adipose-derived mesenchymal stem cells with silk fibroin 3D scaffolds. Biomaterials 2007, 28, 5280–5290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, R.D.; Wang, R.Y.; Reagan, M.R.; Chen, Y.; Borowsky, F.E.; Zieba, A.; Marra, K.G.; Rubin, J.P.; Ghobrial, I.M.; Kaplan, D.L. The use of silk as a scaffold for mature, sustainable unilocular adipose 3D tissue engineered systems. Adv. Health Mater. 2016, 5, 1667–1677. [Google Scholar] [CrossRef] [Green Version]
- Abbott, R.D.; Borowsky, F.E.; Alonzo, C.A.; Zieba, A.; Georgakoudi, I.; Kaplan, D.L. Variability in responses observed in human white adipose tissue models. J. Tissue Eng. Regen. Med. 2017, 12, 840–847. [Google Scholar] [CrossRef]
- Tytgat, L.; Kollert, M.R.; Van Damme, L.; Thienpont, H.; Ottevaere, H.; Duda, G.N.; Geissler, S.; Dubruel, P.; Van Vlierberghe, S.; Qazi, T.H. Evaluation of 3D printed gelatin-based scaffolds with varying pore size for MSC-based adipose tissue engineering. Macromol. Biosci. 2020, 20, e1900364. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Liu, Y.; Hui, L. Preparation and characterization of acellular adipose tissue matrix using a combination of physical and chemical treatments. Mol. Med. Rep. 2017, 17, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Flynn, L.E. The use of decellularized adipose tissue to provide an inductive microenvironment for the adipogenic differentiation of human adipose-derived stem cells. Biomaterials 2010, 31, 4715–4724. [Google Scholar] [CrossRef]
- Choi, J.S.; Yang, H.-J.; Kim, B.G.; Kim, J.D.; Lee, S.H.; Lee, E.K.; Park, K.; Cho, Y.W.; Lee, H.Y. Fabrication of porous extracellular matrix scaffolds from human adipose tissue. Tissue Eng. Part. C Methods 2010, 16, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Gao, J.; Ogawa, R. Mechanobiology and mechanotherapy of adipose tissue-effect of mechanical force on fat tissue engineering. Plast. Reconstr. Surg. Glob. Open 2015, 3, e578. [Google Scholar] [CrossRef]
- Zhang, T.; Lin, S.; Shao, X.; Shi, S.; Zhang, Q.; Xue, C.; Lin, Y.; Zhu, B.; Cai, X. Regulating osteogenesis and adipogenesis in adipose-derived stem cells by controlling underlying substrate stiffness. J. Cell. Physiol. 2017, 233, 3418–3428. [Google Scholar] [CrossRef] [PubMed]
- Pope, B.D.; Warren, C.R.; Parker, K.K.; Cowan, C.A. Microenvironmental control of adipocyte fate and function. Trends Cell Biol. 2016, 26, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Van, R.L.A.; Roncari, D. Complete differentiation of adipocyte precursors. A culture system for studying the cellular nature of adipose tissue. Cell Tissue Res. 1978, 195, 317–329. [Google Scholar]
- Or-Tzadikario, S.; Sopher, R.; Gefen, A. Quantitative monitoring of lipid accumulation over time in cultured adipocytes as function of culture conditions: Toward controlled adipose tissue engineering. Tissue Eng. Part. C Methods 2010, 16, 1167–1181. [Google Scholar] [CrossRef]
- Veilleux, A.; Caron-Jobin, M.; Noël, S.; Laberge, P.Y.; Tchernof, A. Visceral Adipocyte hypertrophy is associated with dyslipidemia independent of body composition and fat distribution in women. Diabetes 2011, 60, 1504–1511. [Google Scholar] [CrossRef] [Green Version]
- Anand, S.S.; Tarnopolsky, M.A.; Rashid, S.; Schulze, K.M.; Desai, D.; Mente, A.; Rao, S.; Yusuf, S.; Gerstein, H.; Sharma, A.M. Adipocyte hypertrophy, fatty liver and metabolic risk factors in South Asians: The molecular study of health and risk in ethnic groups (mol-SHARE). PLoS ONE 2011, 6, e22112. [Google Scholar] [CrossRef] [Green Version]
- Notbohm, J.; Lesman, A.; Tirrell, D.A.; Ravichandran, G. Quantifying cell-induced matrix deformation in three dimensions based on imaging matrix fibers. Integr. Biol. 2015, 7, 1186–1195. [Google Scholar] [CrossRef] [Green Version]
- Petroll, W.M.; Cavanagh, H.D.; Jester, J.V. Dynamic three-dimensional visualization of collagen matrix remodeling and cytoskeletal organization in living corneal fibroblasts. Scanning 2004, 26, 1–10. [Google Scholar] [CrossRef]
- Mohammadi, H.; Arora, P.D.; Simmons, C.A.; Janmey, P.A.; McCulloch, C.A. Inelastic behaviour of collagen networks in cell–matrix interactions and mechanosensation. J. R. Soc. Interface 2015, 12, 20141074. [Google Scholar] [CrossRef] [Green Version]
- Ban, E.; Franklin, J.M.; Nam, S.; Smith, L.R.; Wang, H.; Wells, R.G.; Chaudhuri, O.; Liphardt, J.T.; Shenoy, V.B. Mechanisms of plastic deformation in collagen networks induced by cellular forces. Biophys. J. 2018, 114, 450–461. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, C.; Bremer, A.; Alba, D.; Apovian, C.; Koethe, J.R.; Koliwad, S.; Lewis, D.; Lo, J.A.; McComsey, G.; Eckard, A.; et al. Obesity and fat metabolism in human immunodeficiency virus–infected individuals: Immunopathogenic mechanisms and clinical implications. J. Infect. Dis. 2019, 220, 420–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, M.D.; Burdick, J.A.; Wells, R.G. Engineered biomaterial platforms to study fibrosis. Adv. Health Mater. 2020, 9, e1901682. [Google Scholar] [CrossRef]
- Loh, Q.L.; Choong, C. Three-dimensional scaffolds for tissue engineering applications: Role of porosity and pore size. Tissue Eng. Part. B Rev. 2013, 19, 485–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Grunsven, L.A. 3D in vitro models of liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, M.; Bansal, R.; Rouwkema, J. Bioengineered 3D models to recapitulate tissue fibrosis. Trends Biotechnol. 2020, 38, 623–636. [Google Scholar] [CrossRef]
- Shoham, N.; Gottlieb, R.; Sharabani-Yosef, O.; Zaretsky, U.; Benayahu, D.; Gefen, A. Static mechanical stretching accelerates lipid production in 3T3-L1 adipocytes by activating the MEK signaling pathway. Am. J. Physiol. Physiol. 2012, 302, C429–C441. [Google Scholar] [CrossRef] [Green Version]
- Lutz, T.; Woods, S.C. Overview of animal models of obesity. Curr. Protoc. Pharmacol. 2012, 58, 5–61. [Google Scholar] [CrossRef] [Green Version]
- Kanasaki, K.; Koya, D. Biology of obesity: Lessons from animal models of obesity. J. Biomed. Biotechnol. 2011, 2011, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; De Angelis, M.H.; Schurmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Vickers, S.P.; Jackson, H.C.; Cheetham, S.C. The utility of animal models to evaluate novel anti-obesity agents. Br. J. Pharmacol. 2011, 164, 1248–1262. [Google Scholar] [CrossRef] [Green Version]
- Chusyd, D.E.; Wang, D.; Huffman, D.M.; Nagy, T.R. Relationships between rodent white adipose fat pads and human white adipose fat depots. Front. Nutr. 2016, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyer, C.; Tataranni, P.A.; Snitker, S.; Danforth, E.; Ravussin, E. Increase in insulin action and fat oxidation after treatment with CL 316,243, a highly selective beta3-adrenoceptor agonist in humans. Diabetes 1998, 47, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Arch, J.R.; Wilson, S. Prospects for beta 3-adrenoceptor agonists in the treatment of obesity and diabetes. Int. J. Obes. Relat. Metab. Disord. 1996, 20, 191–199. [Google Scholar] [PubMed]
- Tavernier, G.; Barbe, P.; Galitzky, J.; Berlan, M.; Caput, D.; Lafontan, M.; Langin, D. Expression of beta3-adrenoceptors with low lipolytic action in human subcutaneous white adipocytes. J. Lipid Res. 1996, 37, 87–97. [Google Scholar]
- Coleman, D.L. Obese and diabetes: Two mutant genes causing diabetes-obesity syndromes in mice. Diabetologia 1978, 14, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Mayer, J.; Bates, M.W.; Dickie, M.M. Hereditary diabetes in genetically obese mice. Science 1951, 113, 746–747. [Google Scholar] [CrossRef]
- Zhang, Y.; Proença, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Chua, S.C.; Chung, W.K.; Wu-Peng, X.S.; Zhang, Y.; Liu, S.-M.; Tartaglia, L.; Leibel, R.L. Phenotypes of mouse diabetes and rat fatty due to mutations in the OB (leptin) receptor. Science 1996, 271, 994–996. [Google Scholar] [CrossRef]
- Bates, S.H.; Kulkarni, R.N.; Seifert, M.; Myers, M.G. Roles for leptin receptor/STAT3-dependent and -independent signals in the regulation of glucose homeostasis. Cell Metab. 2005, 1, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.H.; Stearns, W.H.; Dundon, T.A.; Schubert, M.; Tso, A.W.K.; Wang, Y.; Banks, A.S.; Lavery, H.J.; Haq, A.K.; Maratos-Flier, E.; et al. STAT3 signalling is required for leptin regulation of energy balance but not reproduction. Nature 2003, 421, 856–859. [Google Scholar] [CrossRef]
- Bray, G.A. The Zucker-fatty rat: A review. Fed. Proc. 1977, 36, 148–153. [Google Scholar] [PubMed]
- Bray, G.A.; York, D.A. Hypothalamic and genetic obesity in experimental animals: An autonomic and endocrine hypothesis. Physiol. Rev. 1979, 59, 719–809. [Google Scholar] [CrossRef] [PubMed]
- Challis, B.G.; Coll, A.P.; Yeo, G.S.H.; Pinnock, S.B.; Dickson, S.L.; Thresher, R.R.; Dixon, J.; Zahn, D.; Rochford, J.J.; White, A.; et al. Mice lacking pro-opiomelanocortin are sensitive to high-fat feeding but respond normally to the acute anorectic effects of peptide-YY3-36. Proc. Natl. Acad. Sci. USA 2004, 101, 4695–4700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaswen, L.; Diehl, N.; Brennan, M.B.; Hochgeschwender, U. Obesity in the mouse model of pro-opiomelanocortin deficiency responds to peripheral melanocortin. Nat. Med. 1999, 5, 1066–1070. [Google Scholar] [CrossRef]
- Corander, M.P.; Rimmington, D.; Challis, B.G.; O’Rahilly, S.; Coll, A.P. Loss of agouti-related peptide does not significantly impact the phenotype of murine POMC deficiency. Endocrinology 2011, 152, 1819–1828. [Google Scholar] [CrossRef]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [Green Version]
- Becerril, S.; Rodríguez, A.; Catalán, V.; Méndez-Giménez, L.; Ramírez, B.; Sainz, N.; Llorente, M.; Unamuno, X.; Gomez-Ambrosi, J.; Frühbeck, G. Targeted disruption of the iNOS gene improves adipose tissue inflammation and fibrosis in leptin-deficient ob/ob mice: Role of tenascin C. Int. J. Obes. 2018, 42, 1458–1470. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, A.; Makino, N.; Tozawa, T.; Shirahata, N.; Honda, T.; Ikeda, Y.; Sato, H.; Ito, M.; Kakizaki, Y.; Akamatsu, M.; et al. Pancreatic fat accumulation, fibrosis, and acinar cell injury in the zucker diabetic fatty rat fed a chronic high-fat diet. Pancreas 2014, 43, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Lasker, S.; Rahman, M.; Parvez, F.; Zamila, M.; Miah, P.; Nahar, K.; Kabir, F.; Sharmin, S.B.; Subhan, N.; Ahsan, G.U.; et al. High-fat diet-induced metabolic syndrome and oxidative stress in obese rats are ameliorated by yogurt supplementation. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Nakazeki, F.; Nishiga, M.; Horie, T.; Nishi, H.; Nakashima, Y.; Baba, O.; Kuwabara, Y.; Nishino, T.; Nakao, T.; Ide, Y.; et al. Loss of periostin ameliorates adipose tissue inflammation and fibrosis in vivo. Sci. Rep. 2018, 8, 8553. [Google Scholar] [CrossRef] [Green Version]
- Vila, I.K.; Badin, P.M.; Marques, M.A.; Monbrun, L.; Lefort, C.; Mir, L.; Louche, K.; Bourlier, V.; Roussel, B.; Gui, P.; et al. Immune cell toll-like receptor 4 mediates the development of obesity- and endotoxemia-associated adipose tissue fibrosis. Cell Rep. 2014, 7, 1116–1129. [Google Scholar] [CrossRef] [PubMed]
- Payette, C.; Blackburn, P.; Lamarche, B.; Tremblay, A.; Bergeron, J.; Lemieux, I.; Després, J.-P.; Couillard, C. Sex differences in postprandial plasma tumor necrosis factor–α, interleukin-6, and C-reactive protein concentrations. Metabolism 2009, 58, 1593–1601. [Google Scholar] [CrossRef]
- Grove, K.L.; Fried, S.K.; Greenberg, A.S.; Xiao, X.; Clegg, D.J. A microarray analysis of sexual dimorphism of adipose tissues in high-fat-diet-induced obese mice. Int. J. Obes. 2010, 34, 989–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macotela, Y.; Boucher, J.; Tran, T.T.; Kahn, C.R. Sex and depot differences in adipocyte insulin sensitivity and glucose metabolism. Diabetes 2009, 58, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, K.E.; Neinast, M.D.; Sun, K.; Skiles, W.M.; Bills, J.D.; Zehr, J.A.; Zeve, D.; Hahner, L.D.; Cox, D.W.; Gent, L.M.; et al. The sexually dimorphic role of adipose and adipocyte estrogen receptors in modulating adipose tissue expansion, inflammation, and fibrosis. Mol. Metab. 2013, 2, 227–242. [Google Scholar] [CrossRef]
- Hilton, C.; Neville, M.J.; Karpe, F. MicroRNAs in adipose tissue: Their role in adipogenesis and obesity. Int. J. Obes. 2012, 37, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Abente, E.; Subramanian, M.; Ramachandran, V.; Najafi-Shoushtari, H.; Najafi-Shoushtari, H. MicroRNAs in obesity-associated disorders. Arch. Biochem. Biophys. 2016, 589, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yang, Y.; Wu, J. TNFalpha-induced up-regulation of miR-155 inhibits adipogenesis by down-regulating early adipogenic transcription factors. Biochem. Biophys. Res. Commun. 2011, 414, 618–624. [Google Scholar] [CrossRef]
- Chen, Y.; Siegel, F.; Kipschull, S.; Haas, B.; Fröhlich, H.; Meister, G.; Pfeifer, A. miR-155 regulates differentiation of brown and beige adipocytes via a bistable circuit. Nat. Commun. 2013, 4, 1769. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, K.T.; Enos, R.T.; Carson, M.S.; Cranford, T.L.; Bader, J.E.; Sougiannis, A.T.; Pritchett, C.; Fan, D.; Carson, J.A.; Murphy, E. miR155 deficiency aggravates high-fat diet-induced adipose tissue fibrosis in male mice. Physiol. Rep. 2017, 5, e13412. [Google Scholar] [CrossRef]
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.C. MiRNA-mediated macrophage polarization and its potential role in the regulation of inflammatory response. Shock 2016, 46, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Y.; Zhong, M.; Suo, Q.; Lv, K. Expression profiles of miRNAs in polarized macrophages. Int. J. Mol. Med. 2013, 31, 797–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonski, K.A.; Gaudet, A.D.; Amici, S.A.; Popovich, P.G.; Guerau-De-Arellano, M. Control of the inflammatory macrophage transcriptional signature by miR-155. PLoS ONE 2016, 11, e0159724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudet, A.D.; Fonken, L.K.; Gushchina, L.V.; Aubrecht, T.G.; Maurya, S.K.; Periasamy, M.; Nelson, R.J.; Popovich, P.G. miR-155 deletion in female mice prevents diet-induced obesity. Sci. Rep. 2016, 6, 22862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Nagai, Y.; Honda, H.; Okamoto, N.; Yamamoto, S.; Hamashima, T.; Ishii, Y.; Tanaka, M.; Suganami, T.; Sasahara, M.; et al. Isoliquiritigenin attenuates adipose tissue inflammation in vitro and adipose tissue fibrosis through inhibition of innate immune responses in mice. Sci. Rep. 2016, 6, 23097. [Google Scholar] [CrossRef] [Green Version]
- Henninger, A.M.J.; Eliasson, B.; Jenndahl, L.E.; Hammarstedt, A. Adipocyte hypertrophy, inflammation and fibrosis characterize subcutaneous adipose tissue of healthy, non-obese subjects predisposed to type 2 diabetes. PLoS ONE 2014, 9, e105262. [Google Scholar] [CrossRef]
- Alba, D.; Farooq, J.A.; Lin, M.Y.C.; Schafer, A.L.; Shepherd, J.; Koliwad, S.K. Subcutaneous fat fibrosis links obesity to insulin resistance in chinese americans. J. Clin. Endocrinol. Metab. 2018, 103, 3194–3204. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.; He, J.; Duan, X.; Reynolds, K.; Wu, X.; Chen, J.; Huang, G.; Whelton, P.K. Body weight and mortality among men and women in China. JAMA 2006, 295, 776–783. [Google Scholar] [CrossRef] [Green Version]
- Jan, V.; Cervera, P.; Maachi, M.; Baudrimont, M.; Kim, M.; Vidal, H.; Girard, P.-M.; LeVan, P.; Rozenbaum, W.; Lombès, A.; et al. Altered fat differentiation and adipocytokine expression are inter-related and linked to morphological changes and insulin resistance in HIV-1-infected lipodystrophic patients. Antivir. Ther. 2004, 9, 555–564. [Google Scholar]
- Lassen, P.B.; Liu, Y.; Tordjman, J.; Poitou, C.; Bouillot, J.-L.; Genser, L.; Zucker, J.-D.; Sokolovska, N.; Aron-Wisnewsky, J.; Charlotte, F.; et al. The FAT score, a fibrosis score of adipose tissue: Predicting weight-loss outcome after gastric bypass. J. Clin. Endocrinol. Metab. 2017, 102, 2443–2453. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawitan, J.A. Prospect of stem cell conditioned medium in regenerative medicine. Biomed. Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, A.T.J.; Toupet, K.; Maumus, M.; Fonteneau, G.; Le Quellec, A.; Jorgensen, C.; Guilpain, P.; Noel, D. Human adipose mesenchymal stem cells as potent anti-fibrosis therapy for systemic sclerosis. J. Autoimmun. 2016, 70, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, A.A.; Ziegler, M.E.; Banyard, D.A.; Wirth, G.A.; Paydar, K.Z.; Evans, G.R.D.; Widgerow, A.D. Adipose-derived tissue in the treatment of dermal fibrosis. Ann. Plast. Surg. 2018, 80, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.; Yang, Y.; Liu, F.; Ye, B.; Chen, Z.; Zheng, M.; Liu, Y. MiR-122 modification enhances the therapeutic efficacy of adipose tissue-derived mesenchymal stem cells against liver fibrosis. J. Cell. Mol. Med. 2017, 21, 2963–2973. [Google Scholar] [CrossRef]
- Paik, K.Y.; Kim, K.H.; Park, J.H.; Lee, J.I.; Kim, O.H.; Hong, H.E.; Seo, H.; Choi, H.J.; Ahn, J.; Lee, T.Y.; et al. A novel antifibrotic strategy utilizing conditioned media obtained from miR-150-transfected adipose-derived stem cells: Validation of an animal model of liver fibrosis. Exp. Mol. Med. 2020, 52, 438–449. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.M.; Harn, H.-J.; Chiou, T.W.; Chuang, M.H.; Chen, C.H.; Chuang, C.-H.; Lin, P.C.; Lin, S.Z. Preconditioned adipose-derived stem cells ameliorate cardiac fibrosis by regulating macrophage polarization in infarcted rat hearts through the PI3K/STAT3 pathway. Lab. Investig. 2019, 99, 634–647. [Google Scholar] [CrossRef]
- Song, Y.; Peng, C.; Lv, S.; Cheng, J.; Liu, S.; Wen, Q.; Guan, G.; Liu, G. Adipose-derived stem cells ameliorate renal interstitial fibrosis through inhibition of EMT and inflammatory response via TGF-beta1 signaling pathway. Int. Immunopharmacol. 2017, 44, 115–122. [Google Scholar] [CrossRef]
- Rivera-Valdés, J.J.; García-Bañuelos, J.; Salazar-Montes, A.; García-Benavides, L.; Rosales-Dominguez, A.; Armendáriz-Borunda, J.; Sandoval-Rodríguez, A. Human adipose derived stem cells regress fibrosis in a chronic renal fibrotic model induced by adenine. PLoS ONE 2017, 12, e0187907. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Sun, N. Adipose-derived mesenchymal stem cells: A new tool for the treatment of renal fibrosis. Stem Cells Dev. 2018, 27, 1406–1411. [Google Scholar] [CrossRef]
- Sun, W.; Ni, X.; Sun, S.; Cai, L.; Yu, J.; Wang, J.; Nie, B.; Sun, Z.; Ni, X.; Cao, X. Adipose-derived stem cells alleviate radiation-induced muscular fibrosis by suppressing the expression of TGF-beta1. Stem Cells Int. 2016, 2016, 5638204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathinasabapathy, A.; Bruce, E.; Espejo, A.; Horowitz, A.; Sudhan, D.; Nair, A.; Guzzo, D.; Francis, J.; Raizada, M.K.; Shenoy, V.; et al. Therapeutic potential of adipose stem cell-derived conditioned medium against pulmonary hypertension and lung fibrosis. Br. J. Pharmacol. 2016, 173, 2859–2879. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, E.J.; Lee, S.Y.; Kim, J.H.; Shim, J.J.; Shin, C.; In, K.H.; Kang, K.H.; Uhm, C.S.; Kim, H.K.; et al. The effect of adipose stem cell therapy on pulmonary fibrosis induced by repetitive intratracheal bleomycin in mice. Exp. Lung Res. 2014, 40, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Han, H.S.; Lee, H.; You, D.; Nguyen, V.Q.; Song, D.-G.; Oh, B.H.; Shin, S.; Choi, J.S.; Kim, J.D.; Pan, C.H.; et al. Human adipose stem cell-derived extracellular nanovesicles for treatment of chronic liver fibrosis. J. Control. Release 2020, 320, 328–336. [Google Scholar] [CrossRef]
- Qu, Y.; Zhang, Q.; Cai, X.; Li, F.; Ma, Z.; Xu, M.; Lu, L.G. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J. Cell. Mol. Med. 2017, 21, 2491–2502. [Google Scholar] [CrossRef]
- Bai, X.; Yan, Y.; Song, Y.-H.; Seidensticker, M.; Rabinovich, B.; Metzele, R.; Bankson, J.A.; Vykoukal, D.; Alt, E.U. Both cultured and freshly isolated adipose tissue-derived stem cells enhance cardiac function after acute myocardial infarction. Eur. Heart J. 2009, 31, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Eitta, R.S.A.; Ismail, A.A.; Abdelmaksoud, R.A.; Ghezlan, N.A.; Mehanna, R. Evaluation of autologous adipose-derived stem cells vs. fractional carbon dioxide laser in the treatment of post acne scars: A split-face study. Int. J. Dermatol. 2019, 58, 1212–1222. [Google Scholar] [CrossRef]
- Lee, J.W.; Park, S.H.; Lee, S.J.; Kim, S.H.; Suh, I.S.; Jeong, H.S. Clinical impact of highly condensed stromal vascular fraction injection in surgical management of depressed and contracted scars. Aesthetic Plast. Surg. 2018, 42, 1689–1698. [Google Scholar] [CrossRef]
- Gentile, P.; De Angelis, B.; Pasin, M.; Cervelli, G.; Curcio, C.B.; Floris, M.; Di Pasquali, C.; Bocchini, I.; Balzani, A.; Nicoli, F.; et al. Adipose-derived stromal vascular fraction cells and platelet-rich plasma. J. Craniofa. Surg. 2014, 25, 267–272. [Google Scholar] [CrossRef]
- Williams, R.M.; Zipfel, W.R.; Webb, W.W. Interpreting second-harmonic generation images of collagen i fibrils. Biophys. J. 2004, 88, 1377–1386. [Google Scholar] [CrossRef] [Green Version]
- Cox, G.; Kable, E.; Jones, A.; Fraser, I.; Manconi, F.; Gorrell, M.D. 3-Dimensional imaging of collagen using second harmonic generation. J. Struct. Biol. 2003, 141, 53–62. [Google Scholar] [CrossRef]
- Chen, X.; Nadiarynkh, O.; Plotnikov, S.; Campagnola, P.J. Second harmonic generation microscopy for quantitative analysis of collagen fibrillar structure. Nat. Protoc. 2012, 7, 654–669. [Google Scholar] [CrossRef] [PubMed]
- Alkhouli, N.; Mansfield, J.; Green, E.; Bell, J.S.; Knight, B.; Liversedge, N.; Tham, J.C.; Welbourn, R.; Shore, A.C.; Kos, K.; et al. The mechanical properties of human adipose tissues and their relationships to the structure and composition of the extracellular matrix. Am. J. Physiol. Metab. 2013, 305, E1427–E1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundarakrishnan, A.; Chen, Y.; Black, L.D.; Aldridge, B.B.; Kaplan, D.L. Engineered cell and tissue models of pulmonary fibrosis. Adv. Drug Deliv. Rev. 2018, 129, 78–94. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Huang, Y.; Yang, X.; Yang, X.; Lin, H.; Feng, G.; Zhu, X.; Zhang, X.D. Viscoelasticity in natural tissues and engineered scaffolds for tissue reconstruction. Acta Biomater. 2019, 97, 74–92. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Material | Cell Type | Cell Source | Key Features |
|---|---|---|---|
| Poly (lactic-co-glycolic acid) (PLGA) | ASCs | Rat [83,84] Human [85] | They successfully showed that this PLGA could foster adipose tissue growth and expansion in a short time frame but was not sustainable for long term cultures. At two months post-implantation, there was a dramatic decrease in adipose tissue in the scaffold. This decrease in adipose tissue is likely due to resorption by the environment. One reason for this is lack of vascularization. |
| Marrow Stromal Cells (MSCs) | Rat [86] | ||
| Hyaluronic-based Biomaterial (Hyaff-11) | ASCs | Human [87] | Over a month long experiment ASCs were able to mature into adipocytes and showed high cell density [87]. |
| Methacrylated Gelatin and Hyaluronan | ASCs | Human [88] | ASCs were able to remain viable and differentiate into adipocytes in a 4-week time span. This approach was limited by the lack of vascularization [88]. |
| Polycaprolactone (PCL) | Embryonic Stem Cells | Mouse [89] | These scaffolds were composed of nano fibers and had an average pore size of 30 um and porosity of about 88%. The stem cells differentiated into functional adipocytes and used the nano fibers similarly to how they use ECM proteins in vivo. |
| ASCs | Human [90] | By comparing random and aligned electrospun PCL fibers to 2D cultures, researchers found that the use of aligned fibers resulted in increased lipid accumulation, decreased proliferation, and closer to physiological glucose uptake in differentiated ASCs. | |
| Bacterial Cellulose Based Biomaterials | MSCs | Mouse [91] | By combining bacterial nanocellulose and alginate, stable porous scaffolds were created. Researchers were able to culture high numbers of adipocytes for 4 weeks. |
| ASCS, Microvascular Endothelial Cells | Human [92] | The co-culture of differentiated ASCs and human microvascular endothelial cells was performed with the goal of creating vascularized adipose tissue constructs. Vascular-like structures were seen in co-culture and when culturing endothelial cells only. | |
| Silk Fibroin | ASCs, MSCs | Human [93]. | Scaffolds seeded with ASCs or MSCs had higher levels of adipogenesis in vivo compared to collagen and poly (lactic acid) (PLA). In vitro studies showed a comparable level of differentiation of ASCs and MSCs in silk, collagen, and PLA. |
| Whole Adipose Tissue (Adipocytes, Stromal Cells, Endothelial Cells) | Human [94,95] | Seeding scaffolds with whole adipose tissue, rather than isolating a specific cell type, creates a more physiologically relevant model. The scaffolds had similar numbers of cells and triglycerides after 3 months of culture compared to after seeding. | |
| Methacrylated Gelatin | MSCs | Human [96] | Three-dimensional printing was used to create microporous methacrylated gelatin scaffolds with varying pore sizes from 230–531 µm. MSCs differentiated in scaffolds regardless of pore size, but there was better spatial distribution and the cells migrated deeper into the scaffolds with the largest pore sizes. |
| Adipose Tissue ECM | ASCs | Human [97,98,99] | These ECM scaffolds have a number of advantages, such as high pore interconnectivity and mechanical properties optimized for adipose tissue. However, processing can affect the scaffolds biocompatibility and be a complex, lengthy process. |
| Disease Treated | Model Species | Delivery Method | Outcomes | |
|---|---|---|---|---|
| Adipose Derived Stem Cells | Systemic Sclerosis [163] | Mouse | Intravenous | Decreased skin thickness and collagen content. |
| Renal Fibrosis [168,169] | Rat | Intravenous | Improved kidney function and reduced fibrotic tissue. | |
| Liver Cirrhosis [165] | Mouse | Intravenous | Increased expression of antifibrotic markers | |
| Cardiac Fibrosis [167,176] | Mouse | Intramyocardial Injection | Improved myocardial function and regeneration. | |
| Rat | ||||
| Pulmonary Fibrosis [173] | Mouse | Intraperitoneal Injection | Lowered production of profibrotic markers and improved symptoms, such as septal thickening and enlarged alveoli. | |
| Muscle Fibrosis [171] | Rabbit | Intramuscular | Lowered collagen fiber production and profibrotic markers. | |
| Dermal Scars [177,178,179] | Human | Subcutaneous Injection | Enhanced tissue regeneration, scar severity and area, and improved the overall cosmetic appearance. | |
| Conditioned Media | Pulmonary Fibrosis [172] | Rat | Intravenous | Found that conditioned media was as effective as ASCs at treating pulmonary fibrosis. |
| Exosomes | Liver Fibrosis [174,175] | Mouse | Intravenous | Downregulated fibrotic markers and reduced collagen deposits. |
| Intrasplenic Injection | ||||
| Secretome | Liver Fibrosis [166] | Mouse | Intravenous | Increased expression of antifibrotic, proliferation, and antioxidant activity markers in the liver. |
| Technique | Testing Method | Results |
|---|---|---|
| Sampling/Biopsies | Human subcutaneous adipose tissue samples can be gathered from live patients during bariatric surgery or cosmetic procedure (panniculectomy, abdominoplasty, liposuction, etc.) [21,79]. Needle biopsies can also be used while patients are under local anesthesia [79,156]. | |
| Staining | Hematoxylin and Eosin | Adipocyte morphology can be determined by staining with hematoxylin and eosin [79]. |
| Picrosirius Red | Collagen can be detected through histological imaging by staining with Picrosirius red [20,21,160]. The images can be analyzed to determine the collagen and adipocyte area in tissue cross sections. | |
| Masson’s Trichrome | Using Masson’s trichrome stain allows for collagen, mucus, nuclei, cytoplasm, keratin, muscle fibers, and erythrocytes to be stained [82]. | |
| Pimonidazole hydrochloride | Pimonidazole hydrochloride can be used to stain cells that are in a hypoxic environment [82]. | |
| Imaging | Polarized Light Microscopy | Polarized light microscopy can detect different collagen types. Under polarized light and stained with Sirius red, type I collagen fibers will appear orange to red, while type II collagen fibers will appear yellow to green [19,160]. |
| Confocal Microscopy | Though traditional histological approaches allow important information to be discerned, fully understanding collagen volume and dispersity can only be evaluated using 3D imaging approaches. Confocal microscopy can be used to discern the differences in adipocyte size and collagen amount between healthy and fibrotic tissues [20,81]. | |
| Second Harmonic Generation (SHG) Microscopy | SHG microscopy can be used to image collagen fibers without staining. This allows for 3-dimensional imaging to evaluate collagen dispersion and structure [81,180,181,182]. SHG microscopy has been paired with coherent anti-Stokes Raman scattering (CARS) and 2-photon fluorescence (TPF) to image adipocytes and elastin [183]. | |
| Scanning Electron Microscopy (SEM) | SEM can be used to visualize adipocytes and ECM fibers. Researchers have used SEM to image collagen fibers [28]. | |
| Transmission Electron Microscopy (TEM) | TEM can be used to see the interstitial space, caveolae, vasculature, and adipocytes [28]. | |
| Mechanical Testing | Tensile Testing | Tensile testing has been investigated as a method of measuring adipose tissue fibrosis [20]. By securing sections of fresh tissue between clamps the peak force and tensile strength can be determined. Samples with higher degrees of fibrosis will exhibit higher peak forces and tensile strengths. Custom made mechanical testing instruments have also been made [183]. |
| Rheological Testing | The storage modulus (G’) can be used to measure stiffness on the macroscale [81] which is correlated with increased collagen content. | |
| Atomic Force Microscopy (AFM) | AFM can be used to quantify stiffness on the microscale [184]. However, AFM only measures the surface stiffness rather than the interior of the sample. | |
| Shearwave Dispersion Ultrasound Vibrometry (SDUV) | SDUV allows tissue elasticity and viscosity to be measured noninvasively using imaging techniques [185]. | |
| Magnetic Resonance Elastography (MRE) | MRE is a magnetic resonance imaging (MRI) technique. MRE allows for mechanical properties, like stiffness, to be investigated noninvasively using imaging techniques [185]. | |
| Gene Expression | PCR, RT-PCR, qPCR, QRTPCR | Specific genes have been linked to adipose tissue fibrosis, such as TGFβ1, αSMA, COL1, and COL6. Biopsied adipose tissue samples can be analyzed through PCR [21,82,154,156]. |
| Assays | Hydroxyproline | Assays can be used to measure the abundance of hydroxyproline, a signature amino acid for fibrillar collagens [82,141]. |
| Glycerol | The levels of lipolysis can be quantified using a glycerol assay [81]. Lipolysis is thought to be affected by adipose tissue fibrosis with some studies having conflicting results [7,8,9]. | |
| Cell Type Frequencies | Flow Cytometry | By staining the cells with specific antibodies flow cytometry can be used to sort cells or count the number of cells in a population. Researchers have used this to quantify the frequency of SVF populations (stem cells, mast cells, and macrophages) [21,58]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeBari, M.K.; Abbott, R.D. Adipose Tissue Fibrosis: Mechanisms, Models, and Importance. Int. J. Mol. Sci. 2020, 21, 6030. https://doi.org/10.3390/ijms21176030
DeBari MK, Abbott RD. Adipose Tissue Fibrosis: Mechanisms, Models, and Importance. International Journal of Molecular Sciences. 2020; 21(17):6030. https://doi.org/10.3390/ijms21176030
Chicago/Turabian StyleDeBari, Megan K., and Rosalyn D. Abbott. 2020. "Adipose Tissue Fibrosis: Mechanisms, Models, and Importance" International Journal of Molecular Sciences 21, no. 17: 6030. https://doi.org/10.3390/ijms21176030
APA StyleDeBari, M. K., & Abbott, R. D. (2020). Adipose Tissue Fibrosis: Mechanisms, Models, and Importance. International Journal of Molecular Sciences, 21(17), 6030. https://doi.org/10.3390/ijms21176030