1. Introduction
Retinitis pigmentosa (RP) is an incurable inherited retinal disease (IRD) that affects approximately 1 in 4000 people [
1] and it most commonly arises from genetic mutations in genes specific to rod photoreceptor cells. It is a rod-cone dystrophy, beginning with rod degeneration and the loss of night vision and peripheral vision. The cone photoreceptor cells then undergo secondary degeneration. In many forms of RP, this secondary cell loss occurs despite the mutated gene not being expressed in cone photoreceptors. The loss of cone photoreceptors is often more debilitating than rod photoreceptor cells loss, as cone photoreceptors are responsible for the central field of vision, high acuity vision and color vision [
2,
3].
The understanding of cone photoreceptor cell survival and health following the degeneration of rod photoreceptor cells is still being elucidated [
4,
5]. Studies of mouse models of RP have identified key components of cone photoreceptor cells but their influence on survival is still poorly understood [
5,
6]. RP can occur as a result of a sporadic mutation or from a variety of inheritance patterns. There are over 200 genes associated with RP (
RetNet [
7]) and a great variety of mutations identified in each gene. The heterogeneity of RP genotypes limits the ability to prepare appropriate gene therapy treatments for all patients. A common intervention approach to preserve cone cell function and prevent cone cell apoptosis would be valuable—a “mutation-independent” strategy. Such a broad treatment could be more widely applied than a narrow therapy targeting only a single mutant variant of RP. Although it would not address the loss of rod photoreceptors, it would potentially offer the opportunity to maintain central visual acuity. In order for this to be a viable gene therapy approach, a greater understanding of the molecular profile of cone photoreceptor cells prior to and during degeneration is critical.
Punzo et al. [
8] performed a broad characterization of gene pathway changes in the retinal transcriptome across four mouse models of RP during cone cell death (
Pde6rd1,
Pde6g−/−,
Rho−/− and
RhoP23H). A third of the downregulated genes identified from microarray data were genes involved in metabolism.
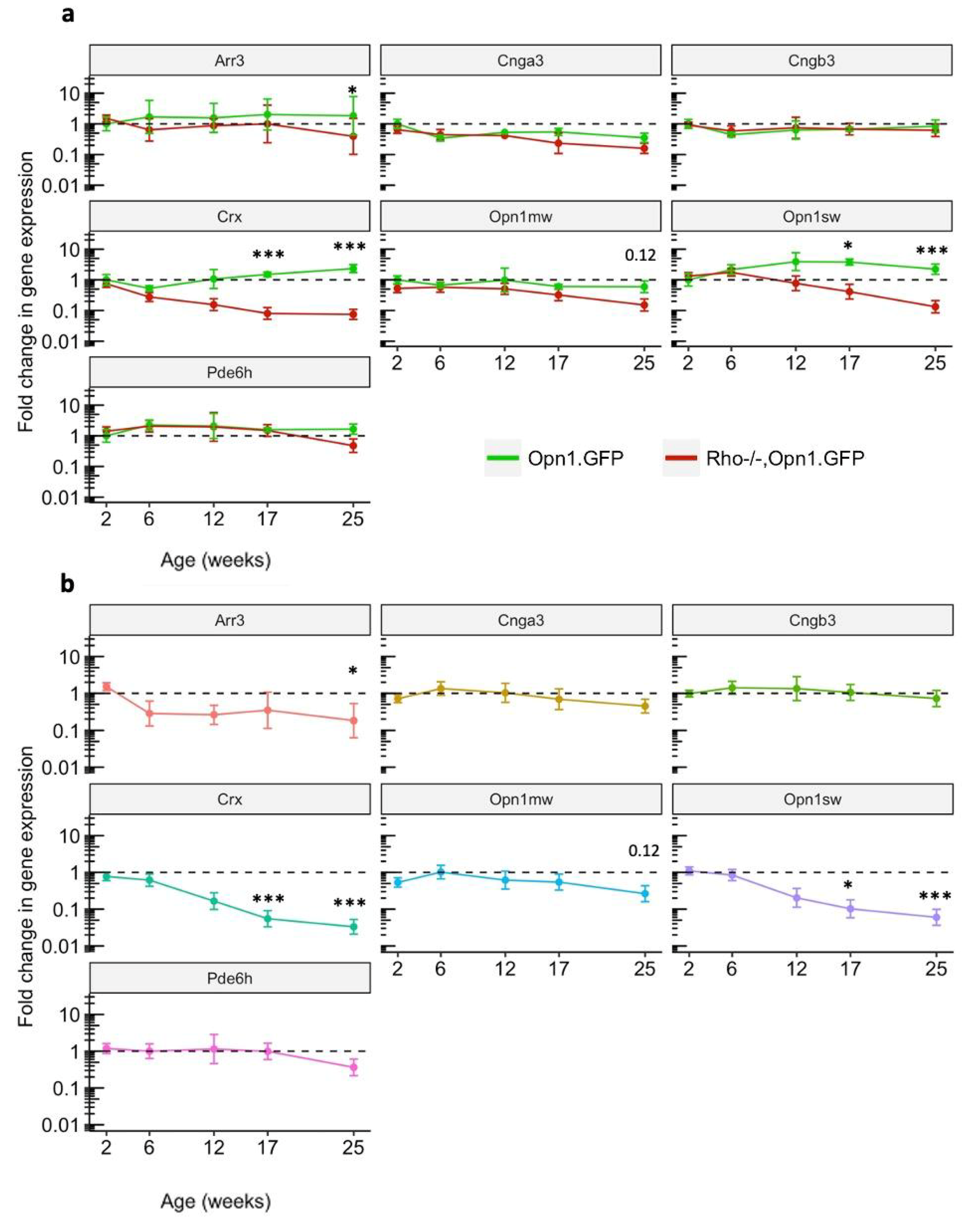
Opn1sw mRNA was downregulated across the different mouse lines, but a cone-specific control for cone cell loss was not included. No papers to date have yet examined changes in cone phototransduction gene expression and correlated those changes with changes in visual or electrophysiological phenotype.
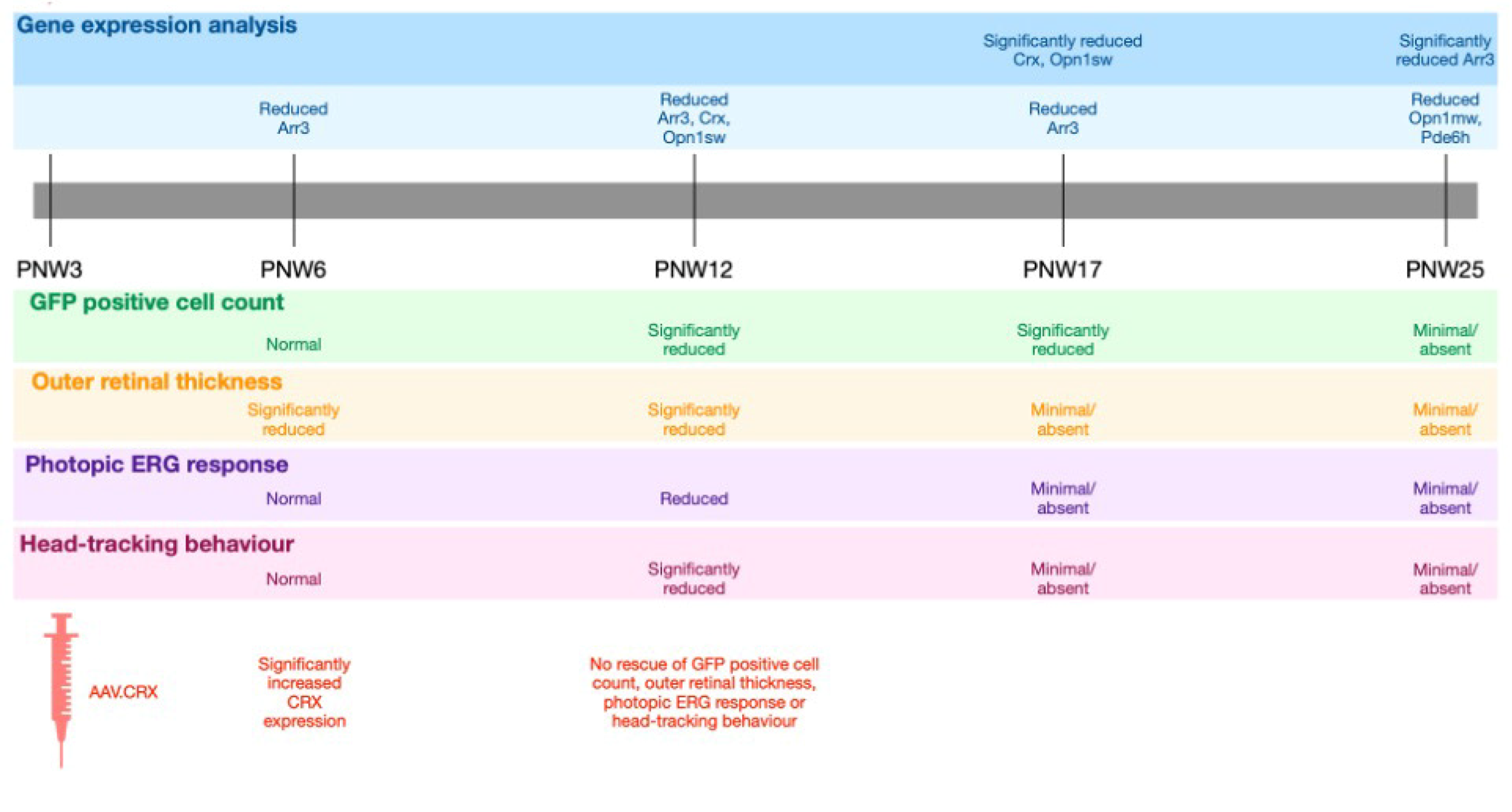
This study measured the changes in expression of genes involved in the cone phototransduction cascade during the period of cone photosensitivity loss in the Rho−/− mouse model. The Rho−/− mouse model used in this report also expressed GFP under the control of the OPN1 promoter in cone photoreceptor cells, providing a control in gene expression profiling that would account for the declining total retinal cone cell count over time in the Rho−/− mouse model.
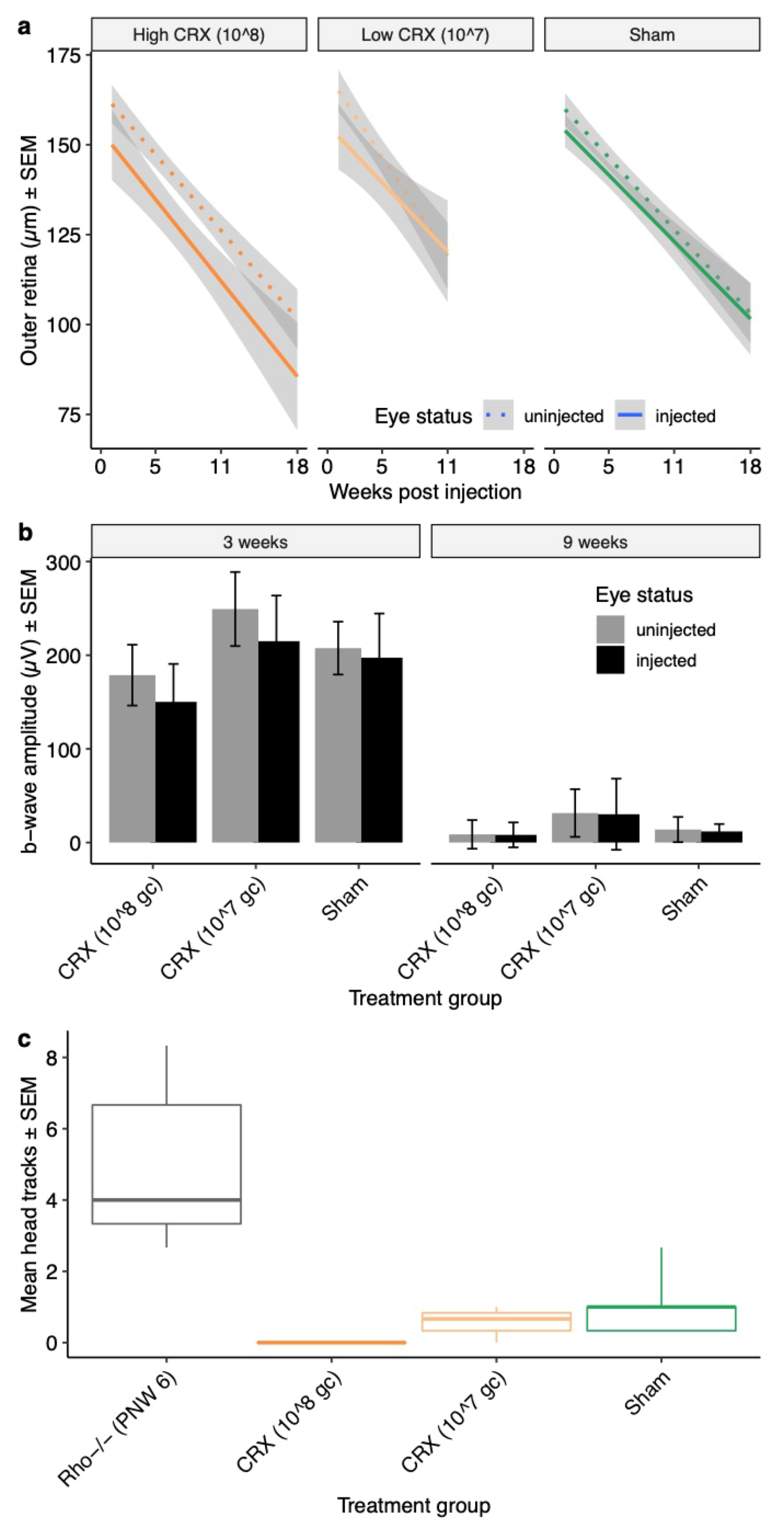
This report identifies significant downregulation of the genes Crx and Opn1sw relative to OPN1-GFP expression levels in the Rho−/− mouse model. These changes occurred prior to detectable changes in cone function and survival. As broad therapeutic approaches are being pursued that might preserve cone photoreceptor cells following primary loss of rod photoreceptor cells, we were interested to identify key regulators that might determine cone cell fate. Since Crx acts upstream of Opn1, we tested the hypothesis that reversing Crx downregulation might delay cone cell death when occurring secondary to rod loss in the Rho−/− mouse model. This did not however prevent cone cell degeneration, implying that downregulation of cone-specific genes such as Crx may be a global secondary effect of cone stress rather than being primary drivers of cone cell degeneration.
3. Discussion
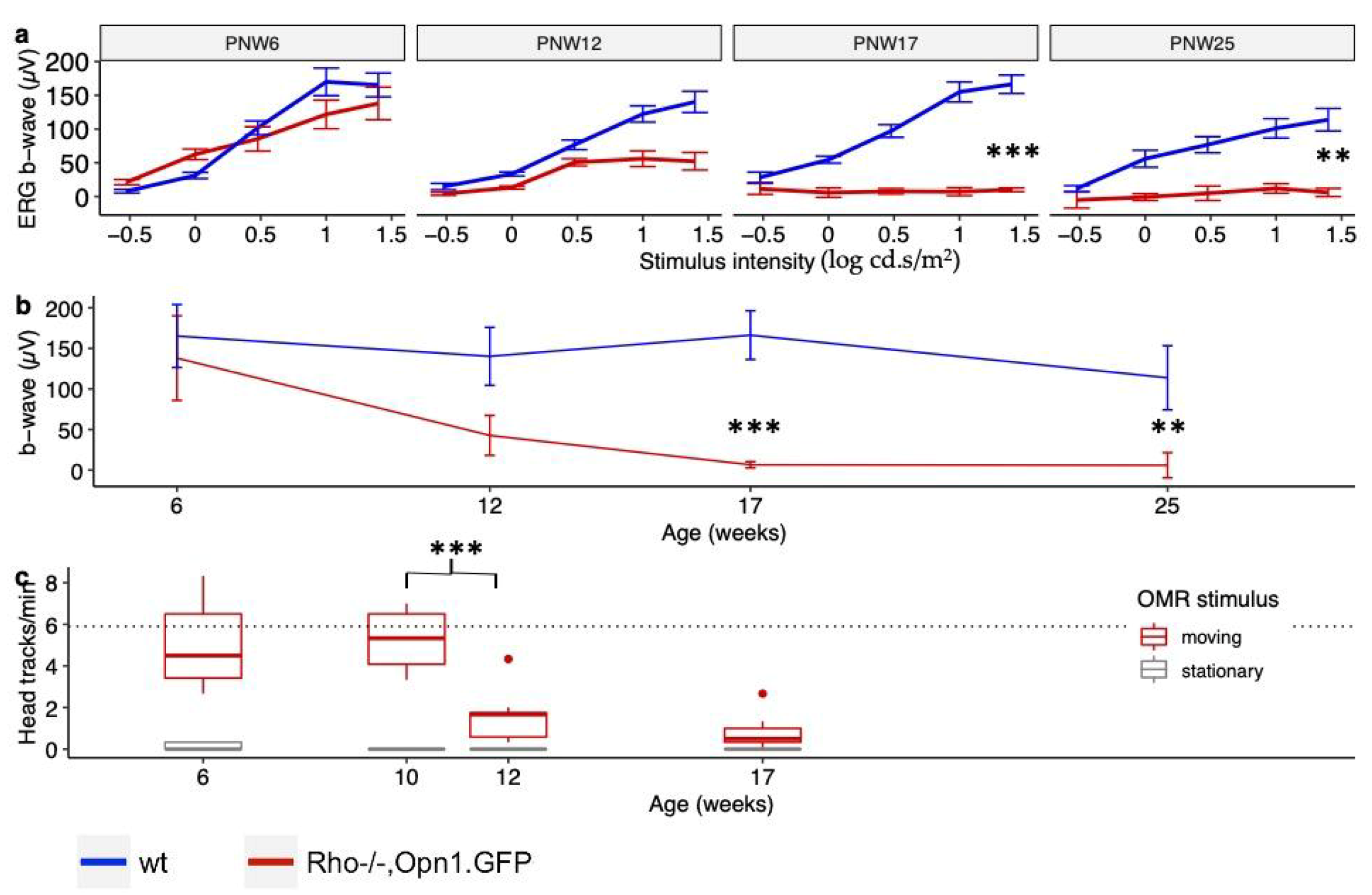
This study details the timeline of cone function loss and the declining expression of cone phototransduction transcripts in the
Rho−/−OPN1-GFP mouse. Data presented here show that cone ERG function in
Rho−/−OPN1-GFP mice declines precipitously between PNW6 and PNW12, coinciding with loss of OMR head-tracking behavior between PNW10 and PNW12. The complete loss of cone function precedes the loss of GFP labelled cones demonstrated in vivo imaging by a number of months. Taken together, this suggests that dormant cone photoreceptors exist in the retina of RP mice long after the extinction of photosensitivity,
Figure 7. Of interest, we identified that the
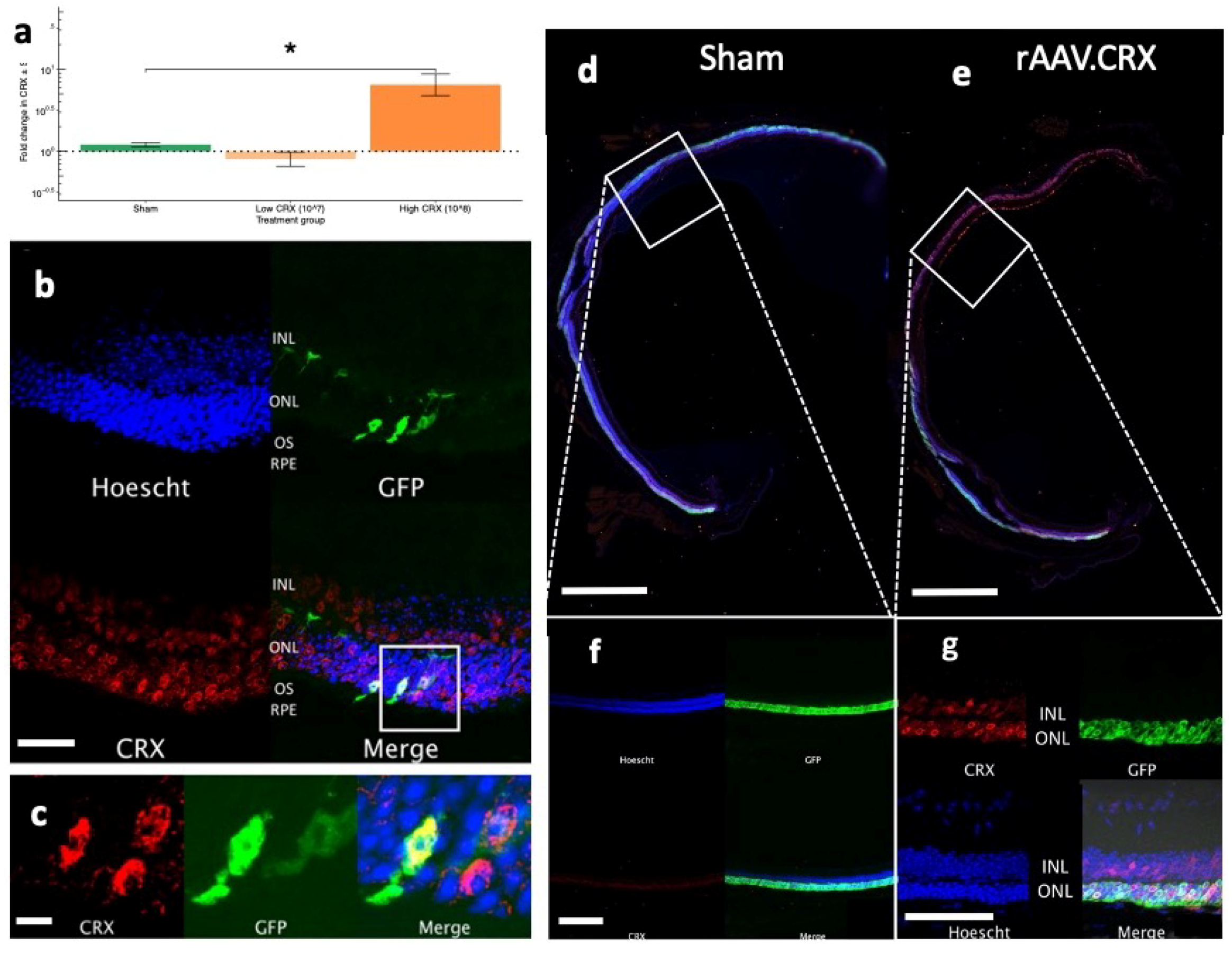
Crx gene was the first of the panel tested to be significantly downregulated. In an attempt to reactivate dormant cone photoreceptors we subsequently delivered
CRX to these mice but observed no improvements in cone photoreceptor survival or function. It is worth noting that cone photoreceptors constitute such a small proportion of the retinal cell population so that even a true cone rescue effect of a portion of retinal cone cells may not be detectable using these measurements.
The rAAV vector was shown to successfully express the
CRX transgene within three weeks of subretinal injection. So too, CRX protein expression was demonstrated in the photoreceptors beginning within weeks and persisting to at least 10 weeks beyond injection. However, the inability of rAAV.
CRX to subsequently rescue or prolong cone photosensitivity raises interesting questions about the role of this gene in the loss of cone function.
CRX is a transcription factor crucial for the differentiation and maintenance of both rod and cone photoreceptors and its downregulation could reasonably contribute to loss of cone photoreceptor function. Humans with
CRX mutations demonstrate cone-rod retinal dystrophy [
13]. In mice,
Crx synergistically modulates
Op1sw and
Opn1sw expression in cones [
14]. Mice with
Crx mutations have down-regulation of genes in the phototransduction cascade, including
Opn1sw,
Opn1mw, and
Arr3 [
15]. However,
Crx is expressed in both cone and rod photoreceptors. The magnitude of
Crx downregulation observed may be mostly caused by rod photoreceptor death, hence the absence of a cone rescue effect from rAAV.
CRX injections.
Arr3 transcript levels appeared lower in Rho−/−OPN1-GFP mice from PNW6 onwards, but never reached significance. Downregulation of Arr3 may be an early and sustained response to declining cone photosensitivity, thereby prolonging G-protein coupled phototransduction signaling.
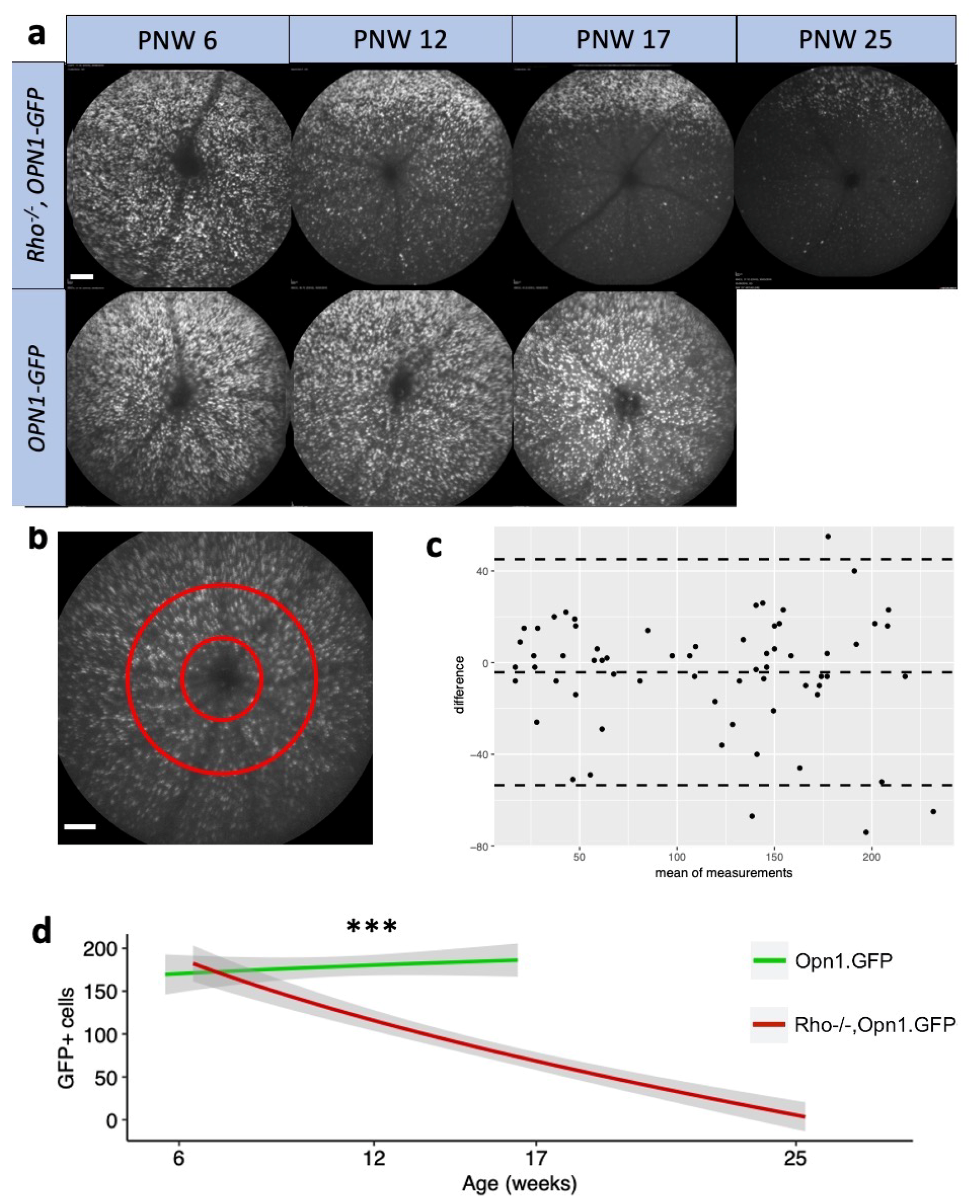
The loss of in vivo cone GFP signal in
Rho−/−OPN1-GFP mice followed a ventral-dorsal pattern (
Figure 3a) similar to cone death kinetics shown histologically elsewhere [
8]. Notably, the relatively unchanged GFP mRNA expression in
Supplementary Figure S1 differed from the clear difference of in vivo GFP protein expression in
Figure 3a,d. The differences between transcript and protein detection could be due to the global reduction in protein translation observed in cells undergoing pro-apoptotic stress [
16]. The differences may also arise from protein mis-localization and misfolding—and thus loss of GFP fluorescence—observed in cone photoreceptors during degeneration [
8].
Opn1mw is expressed throughout the cones of the dorsal and ventral retina, whereas Opn1sw is expressed mainly in the dorsal retinal cones. The significant downregulation of dorsal
Opn1sw mRNA differed from the unchanged ventral-dorsal
Opn1mw mRNA levels (
Figure 1) and, taken together, deviated from the ventral-dorsal pattern of in vivo cone GFP signal loss (
Figure 3a)
. This difference may reflect differential regulation of the transcriptome in S-cones and M-cones, or alternatively suggests that S-cones may be more vulnerable to cell death in RP than M-cones. The unchanged
Opn1mw mRNA levels is further evidence that loss of in vivo GFP protein expression in
Figure 3a is due to translational mechanisms, not only cone cell loss.
Punzo et al. demonstrated that genes involved in metabolism are greatly down-regulated across multiple mouse models of RP. In particular, insulin/mTOR glucose metabolism pathways are most affected [
8]. Similarly, Rod derived Cone Viability Factor (RdCVF) deficiency accelerates cone death in RP rodent models [
17], whilst RdCVF supplementation stimulates aerobic glycolysis in cones [
18]. The work on insulin/mTOR and RdCVF both converge on the central importance of metabolic starvation in cone photoreceptor death in RP. Both papers identify the high energetic demands of photoreceptor outer segment turnover as early casualties of starving photoreceptors. Our data confirms this negative impact of metabolic starvation on cone opsin transcripts, as well as reduced translation and function of cone proteins such as GFP.
The further characterisation and treatment of upstream common causes—such as metabolic starvation—appear more promising than addressing cone photoreceptor specific transcriptome changes. Developing cone-specific therapies remain important until the wider search for effective genetic and molecular therapies for rod rescue in RP is successful.
4. Materials and Methods
4.1. Mice and Breeding
In order to study the effect of rod degeneration on the cone phototransduction cascade, the Rho−/− mouse model of RP also expressed GFP in a subset of cone photoreceptors. This GFP expression provided a control for any potential decline in total retinal cone cell count.
All animals used in this study were treated humanely in accordance with the UK Home Office Regulations and the ARVO statement for the use of animals in ophthalmic and visual research. Animal ethics were approved under UK Home Office project license (PPL) and were also approved by the Oxford animal and welfare review board (PPL 30/2808 from 21 December 2010 to 21 December 2015 and PPL 30/3363 from 21 December 2015 to 21 December 2020). Animals were housed in a 12:12h light-dark cycle.
Rhotm1Phm mice (herein known as
Rho−/− mice) have targeted disruption of the
Rhodopsin gene, resulting in rod degeneration [
10]. Tg(OPN1LW-EGFP)85933Hue mice express enhanced GFP in a subset of cone photoreceptors and are herein known as
OPN1-GFP mice [
19]. Tg(Nrl-EGFP)1Asw mice express enhanced GFP only in the rod photoreceptors and are herein known as
Nrl-GFP mice [
20]. All mice were genotyped using the published PCR protocols or cSLO screening around the age of weaning.
OPN1-GFP breeders were crossed with
Rho−/− mice to produce F1 progeny with the RP phenotype and GFP expression in the cone photoreceptors (
Rho−/−OPN1-GFP mice).
Nrl-GFP breeders were crossed with Rho
−/− mice to produce F1 progeny with the RP phenotype and GFP expression in rod photoreceptors.
4.2. Electroretinography
As a means of assessing cone electrophysiological function during the period of cone photosensitivity loss, electroretinography (ERG) was performed on anaesthetized mice. Mice were positioned on a heated mat in front of the testing console (Colordome Electroretinography machine; Diagnosys LLC, Vision Park, Cambridge, UK) and isolated cone function was tested as described elsewhere [
12]. Single-flash stimuli after light adaptation consisted of 25 (1.5 log) cd.s/m
2; 20 responses were averaged, with an ISI of 1 s.
4.3. Optomotor Response Testing
As a method of assessing integrated cone function during the period of cone photosensitivity loss, a custom optomotor system was produced, consisting of a rotating cylinder that allowed rotation speed to be specified. The cylinder was lined with a square-wave grating pattern of vertical black and white stripes, with a spatial frequency of the grating pattern of 0.1 cycles per degree. The test was performed under photopic conditions: the chamber was illuminated from above with a halogen white light source such that the illumination measured at the position of the animal was 2574 lux. To isolate cone responses, mice were light-adapted on the elevated testing platform for five minutes beforehand and then their head-tracking response to the rotating drum testing protocol quantified as described elsewhere [
21].
4.4. Confocal Scanning Laser Ophthalmoscopy (cSLO) and Optical Coherence Tomography (OCT)
To accurately measure the rod degeneration and any decline in cone photoreceptor cell counts over time, the retinal phenotype was assessed in vivo using confocal scanning laser ophthalmoscopy and spectral domain optical coherence tomography (Spectralis HRA; Heidelberg Engineering, Heidelberg, Germany) as previously described [
22]. A standardized region of interest was sampled from the en face fundus images captured with cSLO (
Appendix A) and counted manually using the ImageJ cell counter plugin to determine the surviving proportion of GFP-positive cones. Total retinal and outer retinal thickness measurements from the OCT were made manually using calipers on Heidelberg software. Total retinal thickness was measured from the inner margin of the nerve fiber layer to the inner margin of the RPE. Outer retinal thickness was measured from the inner margin of the outer nuclear layer to the inner margin of the RPE.
4.5. mRNA Extraction and cDNA Reverse Transcription from Retinal Tissue
To reliably isolate the retinal transcriptome for analysis at the desired timepoints, groups of animals were euthanized at postnatal week (PNW) 2, 6, 12, 17 or 25, the whole neurosensory retina of each eye was harvested by dissection and stored in RNAlater (Thermo Fisher Scientific, Waltham, MA, USA) at −20 °C. Fluorescence-activated cell sorting (FACS) was avoided to prevent cone cell loss and processing-induced changes in the cone transcriptome. RNA was extracted from the retinas using the manufacturer’s instructions for the RNeasy mini kit (QIAgen, Manchester, UK) and the recommended on-column DNase digest with the RNase-Free DNase set (QIAgen, Manchester, UK). The retinal samples were homogenized with a rotor and 30-gauge needle prior to processing. Eluted retinal RNA was reverse transcribed to cDNA using the Superscript III synthesis system (Life Technologies, Paisley, UK) and the included oligo-dT primer. The manufacturer’s instructions were used.
4.6. Transcriptome Analysis Using qPCR
To measure the changes in expression of genes involved in cone function in the
Rho−/− mouse model, custom oligonucleotide primers for qPCR were designed to target six genes of the cone phototransduction cascade:
Opn1mws,
Opn1sws,
Arr3,
Pde6h,
Cnga3,
Cngb3; as well as
Crx (
Supplementary Table S2). By selecting cone-specific genes, there is no need to remove other cells using FACS, thereby avoiding loss of cells and reducing processing-induced changes in the cone transcriptome. These genes are most relevant because they provide cone photoreceptor function.
Model validation, normalization and reference primer pairs were also designed for
GFP and
Actb. Four to six pairs for each gene were first tested and optimized using non-degenerate
OPN1-GFP mouse retinal cDNA dilution series. The nucleotide sequences of the qPCR primers are provided in
Supplementary Table S2; from a list of candidates for each gene, the primer pair with 95–103% efficiency was chosen for the test experiments.
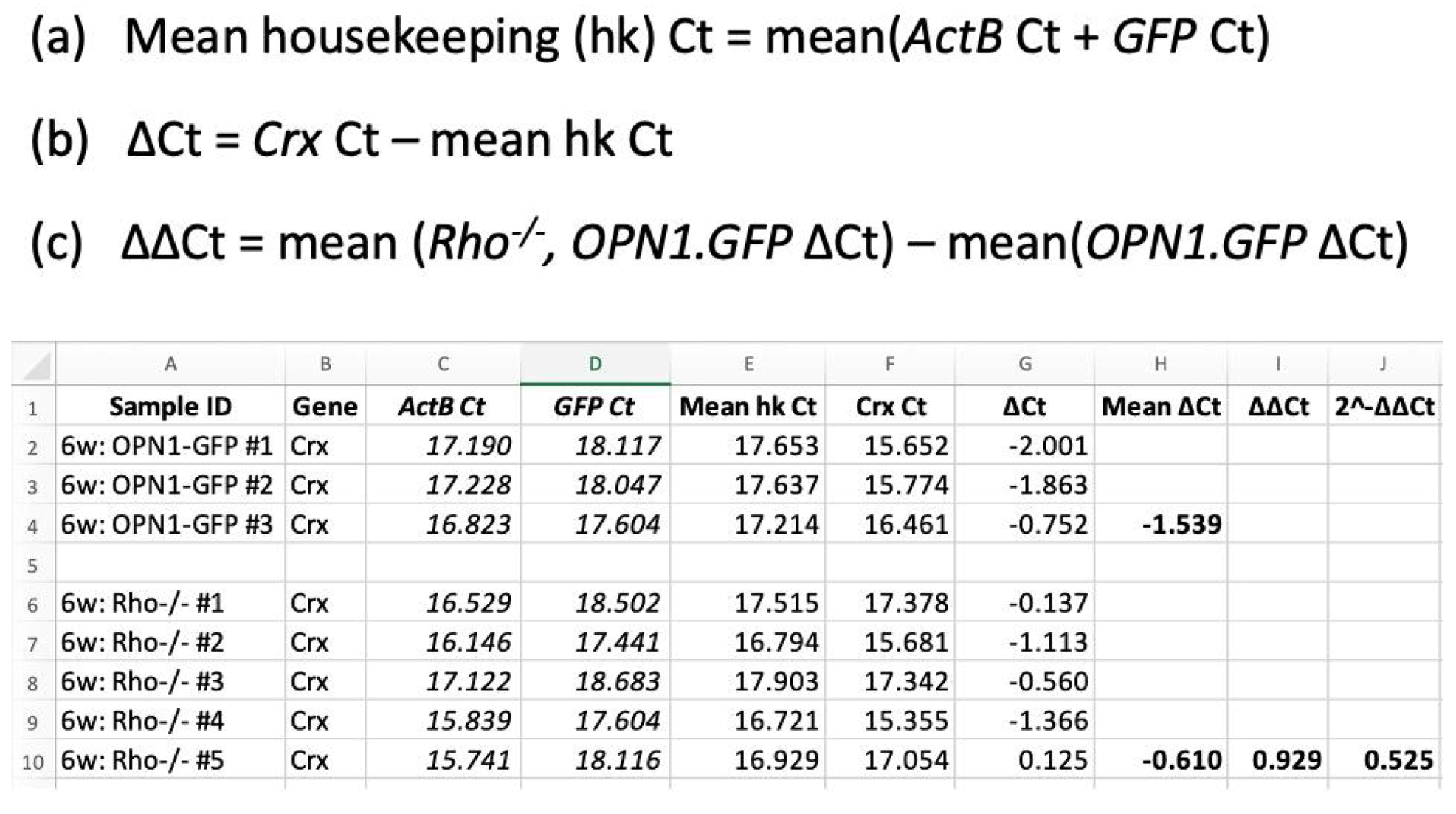
All reactions used a commercial qPCR kit (SYBR Green PCR Master Mix; Applied Biosystems, Foster City, CA, USA) and 2 µM final concentration of each forward and reverse primers. All qPCR experiments were performed in triplicate using a commercial real-time PCR machine (CFX Connect Optics Module, BioRad, Hercules, CA, USA). Reactions were performed with the following settings: an initial denaturation step of 95 °C for 10 min, followed by 40 cycles of: 95 °C for 30 s, an annealing temperature of 55 °C and extension at 72 °C for 30 s. Values obtained for the target genes were reported using the 2
−ΔΔCt method [
9], as detailed in the worked example in
Figure A1. Firstly, test gene Ct values were normalized to the geometric mean of the two housekeeping genes,
ActB and
GFP, to calculate ΔCt. This provided a double normalization that accounted for variability Secondly, ΔCt values for each mouse type at each timepoint were compared to the
OPN1-GFP mice at PNW2 to calculate ΔΔCt. Results were interpreted alongside the
GFP results to account for the decline in cone numbers.
4.7. Recombinant AAV Production and Intraocular Injection
The most down-regulated gene (CRX) was cloned into an rAAV vector and over-expressed in the retina of Rho−/− mice to assess the effect on prolonging or preserving cone photosensitivity in RP. The individual effects of over-expressing CRX was explored across two rAAV doses. This study used an early practical time point, postnatal day (PND) 21, for subretinal injection in order to maximize the chance of transgene expression prior to endogenous cone gene downregulation.
rAAV2/2.CAG.CRX.WPRE.pA (AAV.CRX) was produced in HEK293T cells. Cultures were transfected with plasmids containing the expression cassette, RepCap and helper sequences. The HEK293T cells were pelleted, lysed, purified using an Iodixanol gradient, and then purified using an Amicon Ultra 100K filter (Merck, Germany).
The rAAV were validated before in vivo use (
Supplementary Figure S3). In vitro expression of transgenic protein was confirmed by transducing confluent HEK293T cells with rAAV. After culture for three days, immunocytochemistry was performed with primary anti-CRX (PA5-32182, Thermo Fisher Scientific, Waltham, MA, USA) antibody. Secondary staining was performed with Alexa-Fluor 568 donkey anti-rabbit (A10042, Thermo Fisher Scientific, Waltham, MA, USA).
rAAV capsid purity was confirmed with SDS-PAGE. The rAAV concentrate was mixed with protein loading buffer (National Diagnostics, Atlanta, GA, USA), heat denatured and separated electrophoretically on a precast gel (Bio-Rad, Hertfordshire, UK). The capsid protein bands were stained using EZBlue (Sigma-Aldrich, Dorset, UK).
Viruses titre was measured using optimised custom qPCR primers targeting CRX (FW-5′-GACAGCAGCAGAAACAGCAG-3′ and RC-5′-GGCTCCAGATGGACACAGTG-3′). rAAV was treated with DNAse I (New England Biolabs, Ipswich, MA, USA) before the capsids were denatured by heating. qPCR was performed as described above and the titre was measured against a known dilution series of pAAV.CAG.CRX.WPRE.pA plasmid.
rAAV suspended in phosphate buffered solution (PBS) to give the desired dose in 1.5 µL was delivered by subretinal injection to Rho−/−, OPN1-GFP mice at PND 21. After the onset of anesthesia, the pupil was dilated. A 6mm circular glass coverslip (VWR International, East Grinstead, UK) was positioned on a viscous coupling gel (Viscotears liquid gel, 0.2 mg/g polyacrylic acid; Alcon Laboratories Ltd., Camberley, UK) and the fundus was visualized using an operating microscope (Leica Biosystems, Wetzlar, Germany).
A Nanofil 10 µL syringe (World Precision Instruments, Sarasota, FL, USA) was assembled with the Nanofil 35G beveled needle (WPI). 1.5 µL of PBS sham or rAAV vector at the chosen titre was delivered by trans-scleral injection into the subretinal space to create a hemiretinal detachment. Antibiotic eye drops (Chloramphenicol 0.5%; Bausch & Lomb, Rochester, NY, USA) and Viscotears were applied topically.
4.8. Immunohistochemistry
In order to demonstrate successful retinal transduction, as well as accurately examine the localization of the CRX transgene, a subset of injected eyes were collected for immunohistochemistry. Following euthanasia and enucleation, the cornea was excised at the limbus. The zonules were blunt dissected with Vannas scissors and the lens extruded whilst avoiding retinal traction. The eyecups were then transferred to 4% (w/v) PFA (Thermo Fisher Scientific, Waltham, MA, USA) for 30 min for initial fixation. The eyecups were then sequentially transferred to 10%, 20% and 30% sucrose and then embedded in OCT compound (VWR International, East Grinstead, UK) and stored at −80 °C until sectioning.
Sample blocks were sectioned in 20 µM slices at −22 °C (Cryotome LSE; Thermo Fisher Scientific, Waltham, MA, USA), placed onto polysine-coated glass slides and allowed to dry overnight. All sections were then permeabilized with dilute Triton-X, blocked with donkey serum and incubated with primary anti-CRX (PA5-32182, Thermo Fisher Scientific, Waltham, MA, USA). Secondary staining was performed with Alexa-Fluor 568 donkey anti-rabbit (A10042, Thermo Fisher Scientific, Waltham, MA, USA). The slides were then counter-stained with 4’,6-diamidino-2-phenylindole (DAPI; Invitrogen, Carlsbad, CA, USA). Coverslips were applied with ProLong Diamond Antifade Mountant (Life Technologies, Paisley, UK).
Retinal sections were viewed on a confocal microscope (LSM710; Zeiss, Oberkochen, Germany). Fluorescent cells were located using fluorescence illumination before taking a series of 0.5 µm thickness overlapping XY optical sections. Fluorescence of Hoechst, GFP, and Alexa 568 were sequentially excited and a stack built. Image processing was performed using Volocity (Perkin-Elmer, Waltham, MA, USA) and ImageJ (version 1.43; National Institutes of Health,
https://imagej.nih.gov/ij) [
23].
4.9. Statistical Analysis
Statistical analysis was performed using R software (v3.5.1) [
24]. Hierarchical linear and non-linear modelling was performed using the
nlme package [
25,
26].

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







