Age-Associated Salivary MicroRNA Biomarkers for Oculopharyngeal Muscular Dystrophy

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
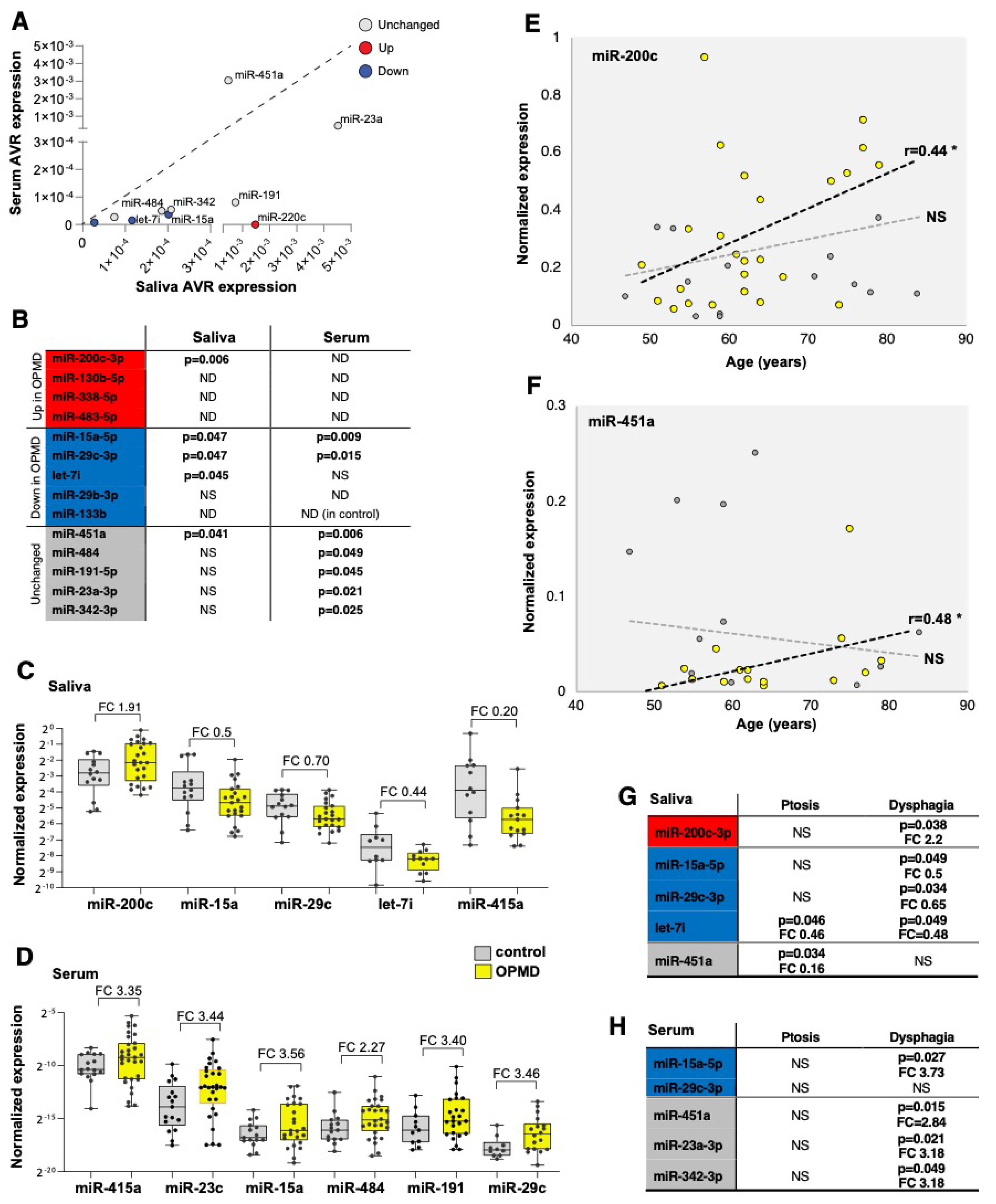
2. Results
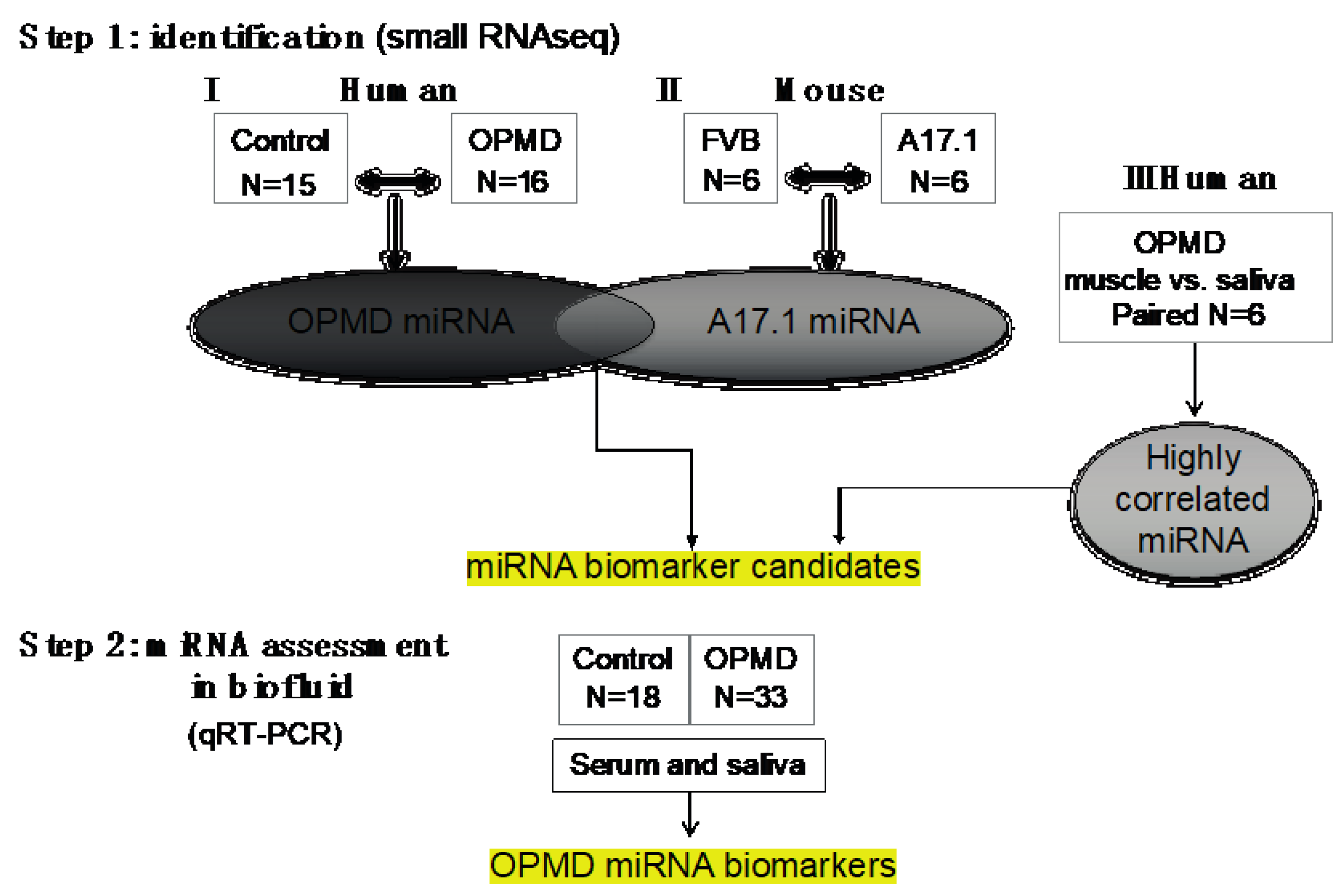
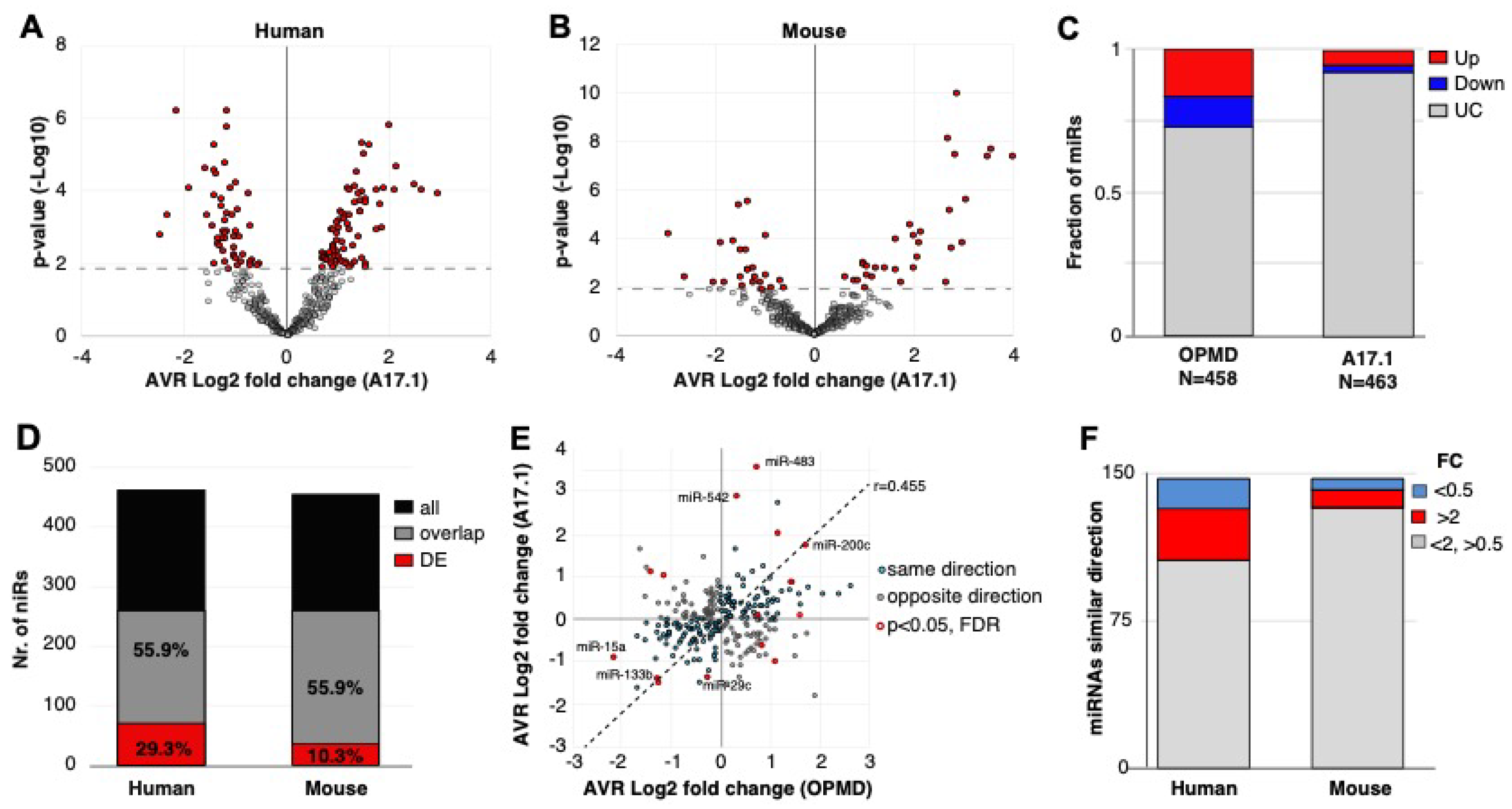
2.1. Small RNAseq in OPMD Patients and the OPMD Mouse Model Revealed Limited Similarities between Differentially Expressed miRNAs
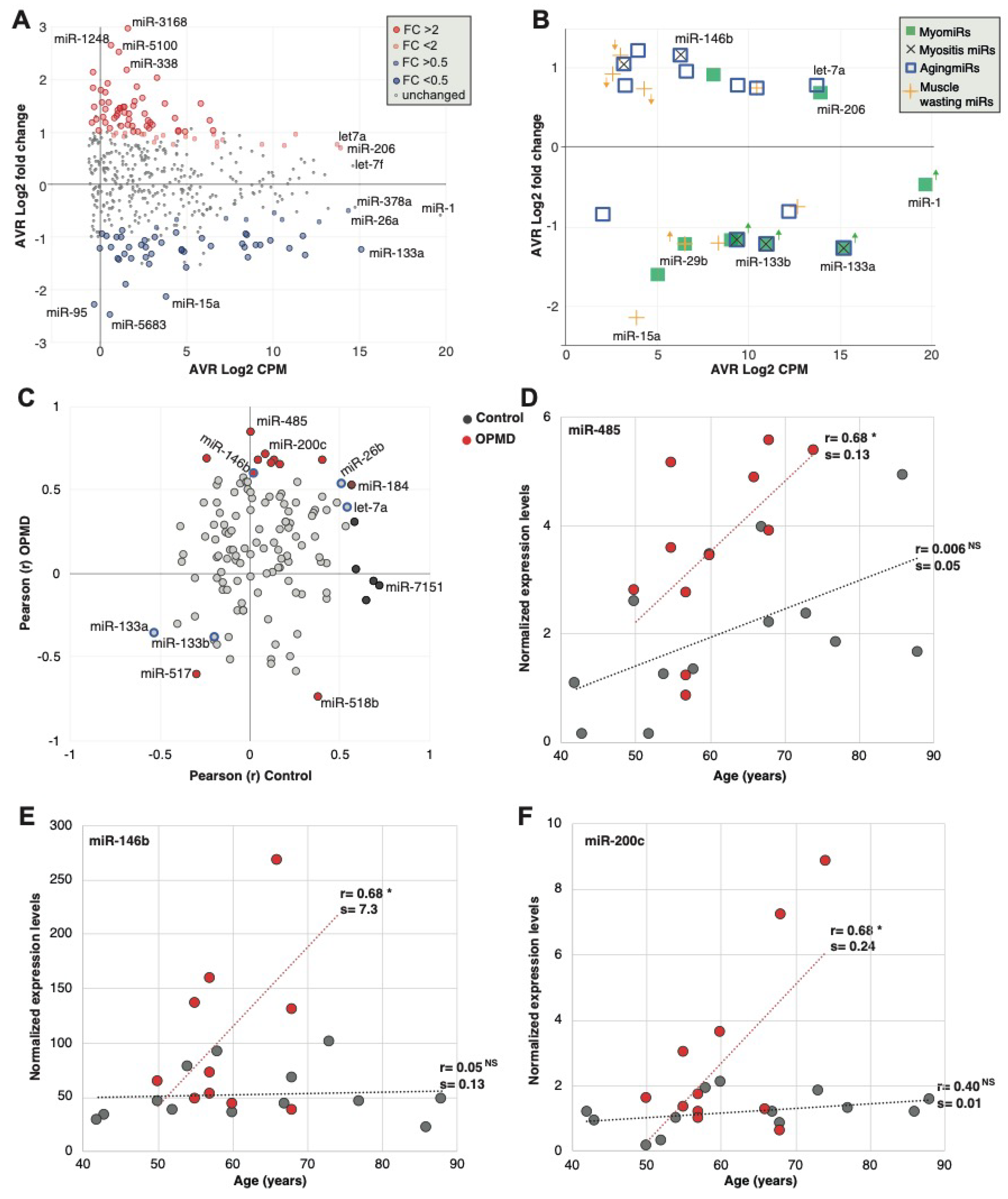
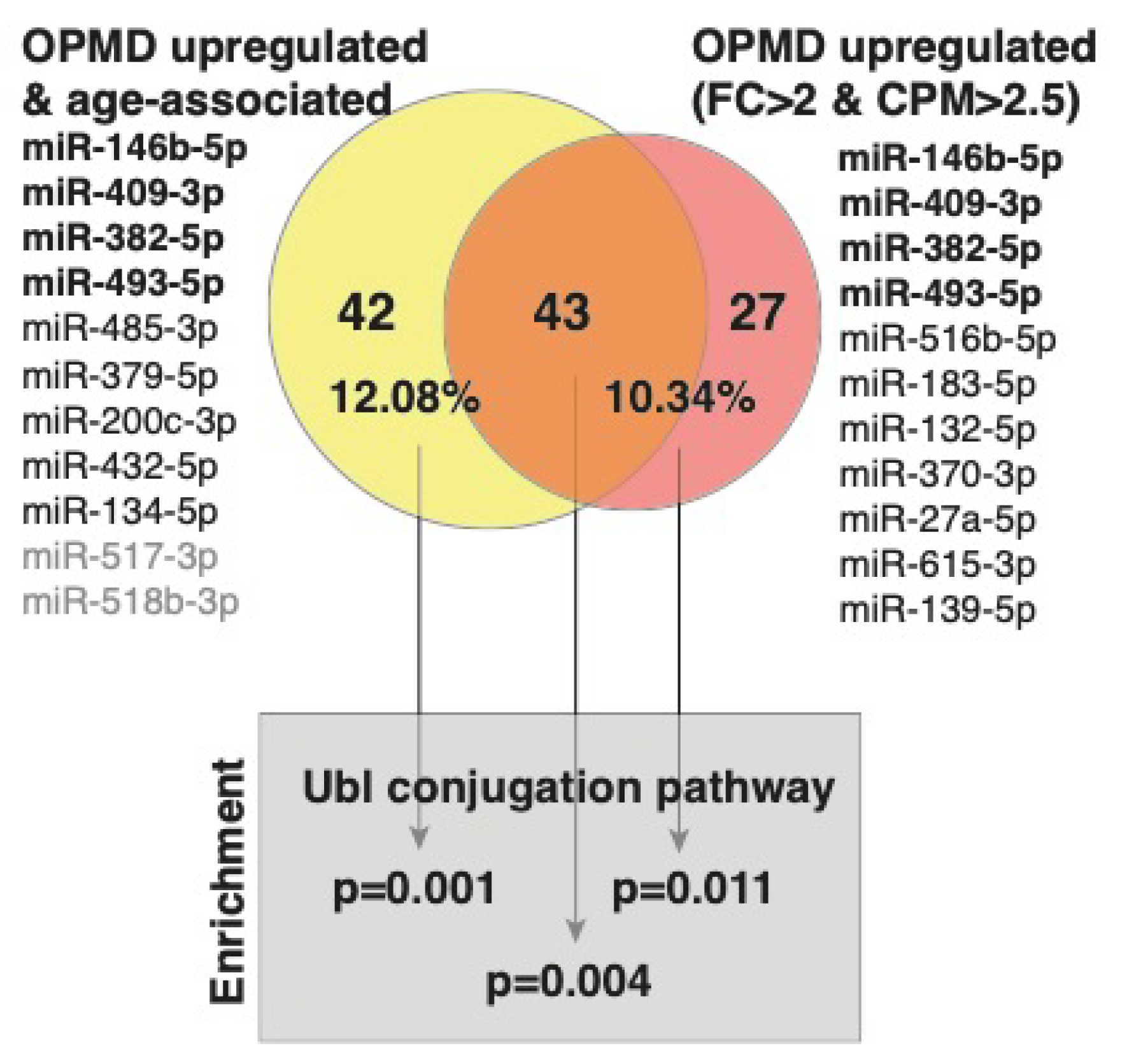
2.2. Characteristics of the OPMD-Deregulated miRNAs in Humans
2.3. Assessment and Verification of miRNA Expression in Biofluids
3. Discussion
4. Material and Methods
4.1. Muscle Tissue and Biofluid Collection
- Only subjects older than 18 years were included.
- OPMD patients were clinically diagnosed by the neurologist based on the common clinical OPMD symptoms. OPMD was genetically confirmed using a genetic test as described in [51]. Patients with OPMD-like symptoms without a genetic confirmation were excluded.
- Healthy controls were the partners of the OPMD patients.
4.2. RNAseq Analysis
4.3. qRT-PCR
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| cpm | Counts per million |
| FDR | False discovery rate |
| IBM | Inclusion body myositis |
| miRNA | MicroRNA |
| NMDs | Neuromuscular disorders |
| OPMD | Oculopharyngeal muscular dystrophy |
| PCA | Principal component analysis |
| QC | Quality control |
| RNAseq | RNA sequencing |
| TA | Tibialis anterior |
| VL | Vastus lateralis |
References
- Shivdasani, R.A. MicroRNAs: Regulators of gene expression and cell differentiation. Blood 2006, 108, 3646–3653. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.K.S.; Phua, Q.H.; Soh, B.-S. Applications of miRNAs in cardiac development, disease progression and regeneration. Stem Cell Res. Ther. 2019, 10, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef]
- Selleck, M.J.; Senthil, M.; Wall, N.R. Making Meaningful Clinical Use of Biomarkers. Biomark. Insights 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, H.B. Predicting Clinical Outcomes Using Molecular Biomarkers. Biomark. Cancer 2016, 8, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Sole, K. Levels of molecular biomarkers can predict rate of disease progression in osteoarthritis. Nat. Clin. Pract. Rheumatol. 2006, 2, 291. [Google Scholar] [CrossRef]
- Lashley, T.; Schott, J.M.; Weston, P.; Murray, C.E.; Wellington, H.; Keshavan, A.; Foti, S.C.; Foiani, M.; Toombs, J.; Rohrer, J.D.; et al. Molecular biomarkers of Alzheimer’s disease: Progress and prospects. Dis. Models Mech. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, M.S.; Kunkel, L.M. Skeletal Muscle MicroRNAs: Their Diagnostic and Therapeutic Potential in Human Muscle Diseases. J. Neuromuscul. Dis. 2015, 2, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, J.J. The MyomiR network in skeletal muscle plasticity. Exerc. Sport Sci. Rev. 2011, 39, 150–154. [Google Scholar] [CrossRef]
- Eisenberg, I.; Eran, A.; Nishino, I.; Moggio, M.; Lamperti, C.; Amato, A.A.; Lidov, H.G.; Kang, P.B.; North, K.N.; Mitrani-Rosenbaum, S.; et al. Distinctive patterns of microRNA expression in primary muscular disorders. Proc. Natl. Acad. Sci. USA 2007, 104, 17016–17021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgantas, R.W.; Streicher, K.; Greenberg, S.A.; Greenlees, L.M.; Zhu, W.; Brohawn, P.Z.; Higgs, B.W.; Czapiga, M.; Morehouse, C.A.; Amato, A.; et al. Inhibition of myogenic microRNAs 1, 133, and 206 by inflammatory cytokines links inflammation and muscle degeneration in adult inflammatory myopathies. Arthritis Rheumatol. 2014, 66, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, J.; Koulmann, N.; Banzet, S. Circulating myomiRs: A new class of biomarkers to monitor skeletal muscle in physiology and medicine. J. Cachexia Sarcopenia Muscle 2018, 9, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.J.; Chaillou, T.; McCarthy, J.J. The role of microRNAs in skeletal muscle health and disease. Front. Biosci. 2015, 20, 37–77. [Google Scholar]
- Brais, B. Oculopharyngeal muscular dystrophy: A polyalanine myopathy. Curr. Neurol. Neurosci. Rep. 2009, 9, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.-Y. Oculopharyngeal muscular dystrophy—An under-diagnosed disease in China? Report a China-born Chinese with PABPN1 mutation and epidemiology review of the literature. J. Formos. Med. Assoc. 2012, 111, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Ruegg, S.; Lehky Hagen, M.; Hohl, U.; Kappos, L.; Fuhr, P.; Plasilov, M.; Muller, H.; Heinimann, K. Oculopharyngeal muscular dystrophy—an under-diagnosed disorder? Swiss. Med. Wkly. 2005, 135, 574–586. [Google Scholar] [PubMed]
- Malerba, A.; Klein, P.; Lu-Nguyen, N.; Cappellari, O.; Strings-Ufombah, V.; Harbaran, S.; Roelvink, P.; Suhy, D.; Trollet, C.; Dickson, G. Established PABPN1 intranuclear inclusions in OPMD muscle can be efficiently reversed by AAV-mediated knockdown and replacement of mutant expanded PABPN1. Hum. Mol. Genet. 2019, 28, 3301–3308. [Google Scholar] [CrossRef]
- Abu-Baker, A.; Kharma, N.; Perreault, J.; Grant, A.; Shekarabi, M.; Maios, C.; Dona, M.; Neri, C.; Dion, P.A.; Parker, A.; et al. RNA-Based Therapy Utilizing Oculopharyngeal Muscular Dystrophy Transcript Knockdown and Replacement. Mol. Ther. Nucleic Acids 2019, 15, 12–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anvar, S.Y.; Raz, Y.; Verway, N.; van der Sluijs, B.; Venema, A.; Goeman, J.J.; Vissing, J.; van der Maarel, S.M.; t Hoen, P.A.; van Engelen, B.G.; et al. A decline in PABPN1 induces progressive muscle weakness in oculopharyngeal muscle dystrophy and in muscle aging. Aging (Albany NY) 2013, 5, 412–426. [Google Scholar] [CrossRef] [Green Version]
- Raz, V.; Sterrenburg, E.; Routledge, S.; Venema, A.; van der Sluijs, B.M.; Trollet, C.; Dickson, G.; van Engelen, B.G.; van der Maarel, S.M.; Antoniou, M.N. Nuclear entrapment and extracellular depletion of PCOLCE is associated with muscle degeneration in oculopharyngeal muscular dystrophy. BMC Neurol. 2013, 13, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trollet, C.; Anvar, S.Y.; Venema, A.; Hargreaves, I.P.; Foster, K.; Vignaud, A.; Ferry, A.; Negroni, E.; Hourde, C.; Baraibar, M.A.; et al. Molecular and phenotypic characterization of a mouse model of oculopharyngeal muscular dystrophy reveals severe muscular atrophy restricted to fast glycolytic fibres. Hum. Mol. Genet. 2010, 19, 2191–2207. [Google Scholar] [CrossRef] [PubMed]
- Koutsoulidou, A.; Mastroyiannopoulos, N.P.; Furling, D.; Uney, J.B.; Phylactou, L.A. Expression of miR-1, miR-133a, miR-133b and miR-206 increases during development of human skeletal muscle. BMC Dev. Biol. 2011, 11, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilahi, E.; Adamecz, Z.; Bodoki, L.; Griger, Z.; Póliska, S.; Nagy-Vincze, M.; Dankó, K. Dysregulated expression profile of myomiRs in the skeletal muscle of patients with polymyositis. EJIFCC 2019, 30, 237–245. [Google Scholar]
- Zanotti, S.; Gibertini, S.; Curcio, M.; Savadori, P.; Pasanisi, B.; Morandi, L.; Cornelio, F.; Mantegazza, R.; Mora, M. Opposing roles of miR-21 and miR-29 in the progression of fibrosis in Duchenne muscular dystrophy. Biochim. Biophys. Acta. Mol. Basis. Dis. 2015, 1852, 1451–1464. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Raz, Y.; van Putten, M.; Paniagua-Soriano, G.; Krom, Y.D.; Florea, B.I.; Raz, V. PABPN1-Dependent mRNA Processing Induces Muscle Wasting. PLoS Genet. 2016, 12, e1006031. [Google Scholar] [CrossRef] [Green Version]
- Vest, K.E.; Phillips, B.L.; Banerjee, A.; Apponi, L.H.; Dammer, E.B.; Xu, W.; Zheng, D.; Yu, J.; Tian, B.; Pavlath, G.K.; et al. Novel mouse models of oculopharyngeal muscular dystrophy (OPMD) reveal early onset mitochondrial defects and suggest loss of PABPN1 may contribute to pathology. Hum. Mol. Genet. 2017, 26, 3235–3252. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chan, M.C.; Yu, Y.; Bei, Y.; Chen, P.; Zhou, Q.; Cheng, L.; Chen, L.; Ziegler, O.; Rowe, G.C.; et al. miR-29b contributes to multiple types of muscle atrophy. Nat. Commun. 2017, 8, 15201. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhang, C.; Zhang, A.; Cai, H.; Price, S.R.; Wang, X.H. MicroRNA-23a and MicroRNA-27a Mimic Exercise by Ameliorating CKD-Induced Muscle Atrophy. J. Am. Soc. Nephrol. 2017, 28, 2631–2640. [Google Scholar] [CrossRef]
- Li, Z.; Cai, B.; Abdalla, B.A.; Zhu, X.; Zheng, M.; Han, P.; Nie, Q.; Zhang, X. LncIRS1 controls muscle atrophy via sponging miR-15 family to activate IGF1-PI3K/AKT pathway. J. Cachexia. Sarcopeni. 2019, 10, 391–410. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Wada, S.; Oikawa, S.; Suzuki, K.; Ushida, T.; Akimoto, T. Loss of microRNA-23–27–24 clusters in skeletal muscle is not influential in skeletal muscle development and exercise-induced muscle adaptation. Sci. Rep. 2019, 9, 1092. [Google Scholar] [CrossRef] [PubMed]
- Needham, M.; Mastaglia, F.L. Sporadic inclusion body myositis: A review of recent clinical advances and current approaches to diagnosis and treatment. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2016, 127, 1764–1773. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; McCarthy, J.J.; Sinha, M.; Spratt, H.M.; Volpi, E.; Esser, K.A.; Rasmussen, B.B. Aging and microRNA expression in human skeletal muscle: A microarray and bioinformatics analysis. Physiol. Genom. 2011, 43, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkes, J.E.; Day, P.J.; Chinoy, H.; Lamb, J.A. The role of microRNAs in the idiopathic inflammatory myopathies. Curr. Opin. Rheumatol. 2015, 27, 608–615. [Google Scholar] [CrossRef]
- Jung, H.J.; Lee, K.P.; Kwon, K.S.; Suh, Y. (2019) MicroRNAs in Skeletal Muscle Aging: Current Issues and Perspectives. J. Gerontol. A. Biol. Sci. Med. Sci. 2019, 74, 1008–1014. [Google Scholar] [CrossRef]
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.J.; Parker, R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev. 2006, 20, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019, 20, 18. [Google Scholar] [CrossRef]
- Anvar, S.Y.; t Hoen, P.A.; Venema, A.; van der Sluijs, B.; van Engelen, B.; Snoeck, M.; Vissing, J.; Trollet, C.; Dickson, G.; Chartier, A.; et al. Deregulation of the ubiquitin-proteasome system is the predominant molecular pathology in OPMD animal models and patients. Skelet Muscle 2011, 1, 15. [Google Scholar] [CrossRef] [Green Version]
- Anvar, S.Y.; Tucker, A.; Vinciotti, V.; Venema, A.; van Ommen, G.J.; van der Maarel, S.M.; Raz, V.; t Hoen, P.A. Interspecies translation of disease networks increases robustness and predictive accuracy. PLoS Comput. Biol. 2011, 7, e1002258. [Google Scholar] [CrossRef]
- Raz, V.; Buijze, H.; Raz, Y.; Verwey, N.; Anvar, S.Y.; Aartsma-Rus, A.; van der Maarel, S.M. A novel feed-forward loop between ARIH2 E3-ligase and PABPN1 regulates aging-associated muscle degeneration. Am. J. Pathol. 2014, 184, 1119–1131. [Google Scholar] [CrossRef]
- Riaz, M.; Raz, Y.; van der Slujis, B.; Dickson, G.; van Engelen, B.; Vissing, J.; Raz, V. Cytokine genes as potential biomarkers for muscle weakness in OPMD. Hum. Mol. Genet. 2016, 25, 4282–4287. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Valdez, G.; Moresi, V.; Qi, X.; McAnally, J.; Elliott, J.L.; Bassel-Duby, R.; Sanes, J.R.; Olson, E.N. MicroRNA-206 Delays ALS Progression and Promotes Regeneration of Neuromuscular Synapses in Mice. Science 2009, 326, 1549–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Williams, A.H.; Maxeiner, J.M.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. microRNA-206 promotes skeletal muscle regeneration and delays progression of Duchenne muscular dystrophy in mice. J. Clin. Invest. 2012, 122, 2054–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, K.N.; Mendell, J.T.; Mendell, J.R.; Rodino-Klapac, L.R. MicroRNA-29 overexpression by adeno-associated virus suppresses fibrosis and restores muscle function in combination with micro-dystrophin. JCI Insight 2017, 2, e93309. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, M.; Torcinaro, A.; Madaro, L.; Marchetti, L.; Sileno, S.; Beji, S.; Salis, C.; Proietti, D.; Imeneo, G.; Capogrossi, M.C.; et al. Role of miR-200c in Myogenic Differentiation Impairment via p66Shc: Implication in Skeletal Muscle Regeneration of Dystrophic mdx Mice. Oxid. Med. Cell. Longev. 2018, 2018, 4814696. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Ye, X.; Xiong, Y.; Zhu, H.; Miao, J.; Zhang, W.; Wan, J. The protective role of microRNA-200c in Alzheimer’s disease pathologies is induced by beta Amyloid-triggered Endoplasmic Reticulum Stress. Front. Mol. Neurosci. 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, Y.; Zhao, L.; Zhang, D.; Yao, X.; Zhang, H.; Wang, Y.-c.; Wang, X.-y.; Xia, H.; Yan, J.; et al. Circulating Muscle-specific miRNAs in Duchenne Muscular Dystrophy Patients. Mol. Ther. Nucleic Acids 2014, 3, e177. [Google Scholar] [CrossRef]
- Cappelletti, C.; Galbardi, B.; Bruttini, M.; Salerno, F.; Canioni, E.; Pasanisi, M.B.; Rodolico, C.; Brizzi, T.; Mora, M.; Renieri, A.; et al. Aging-associated genes and let-7 microRNAs: A contribution to myogenic program dysregulation in oculopharyngeal muscular dystrophy. FASEB J. 2019, 33, 7155–7167. [Google Scholar] [CrossRef]
- Gidaro, T.; Negroni, E.; Perié, S.; Mirabella, M.; Lainé, J.; Lacau St Guily, J.; Butler-Browne, G.; Mouly, V.; Trollet, C. Atrophy, fibrosis, and increased PAX7-positive cells in pharyngeal muscles of oculopharyngeal muscular dystrophy patients. J. Neuropathol. Exp. Neurol. 2013, 72, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Van der Sluijs, B.M.; Raz, V.; Lammens, M.; van den Heuvel, L.P.; Voermans, N.C.; van Engelen, B.G. Intranuclear Aggregates Precede Clinical Onset in Oculopharyngeal Muscular Dystrophy. J. Neuromuscul. Dis. 2016, 3, 101–109. [Google Scholar] [CrossRef]
- De Leeuw, R.H.; Garnier, D.; Kroon, R.; Horlings, C.G.C.; De Meijer, E.; Buermans, H.; Van Engelen, B.G.M.; De Knijff, P.; Raz, V. Diagnostics of short tandem repeat expansion variants using massively parallel sequencing and componential tools. Eur. J. Hum. Genet. EJHG 2019, 27, 400–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raz, V.; Kroon, R.H.M.J.M.; Mei, H.; Riaz, M.; Buermans, H.; Lassche, S.; Horlings, C.; Swart, B.D.; Kalf, J.; Harish, P.; et al. Age-Associated Salivary MicroRNA Biomarkers for Oculopharyngeal Muscular Dystrophy. Int. J. Mol. Sci. 2020, 21, 6059. https://doi.org/10.3390/ijms21176059
Raz V, Kroon RHMJM, Mei H, Riaz M, Buermans H, Lassche S, Horlings C, Swart BD, Kalf J, Harish P, et al. Age-Associated Salivary MicroRNA Biomarkers for Oculopharyngeal Muscular Dystrophy. International Journal of Molecular Sciences. 2020; 21(17):6059. https://doi.org/10.3390/ijms21176059
Chicago/Turabian StyleRaz, Vered, Rosemarie H. M. J. M. Kroon, Hailiang Mei, Muhammad Riaz, Henk Buermans, Saskia Lassche, Corinne Horlings, Bert De Swart, Johanna Kalf, Pradeep Harish, and et al. 2020. "Age-Associated Salivary MicroRNA Biomarkers for Oculopharyngeal Muscular Dystrophy" International Journal of Molecular Sciences 21, no. 17: 6059. https://doi.org/10.3390/ijms21176059
APA StyleRaz, V., Kroon, R. H. M. J. M., Mei, H., Riaz, M., Buermans, H., Lassche, S., Horlings, C., Swart, B. D., Kalf, J., Harish, P., Vissing, J., Kielbasa, S., & van Engelen, B. G. M. (2020). Age-Associated Salivary MicroRNA Biomarkers for Oculopharyngeal Muscular Dystrophy. International Journal of Molecular Sciences, 21(17), 6059. https://doi.org/10.3390/ijms21176059