Expression and Functionality Study of 9 Toll-Like Receptors in 33 Drug-Naïve Non-Affective First Episode Psychosis Individuals: A 3-Month Study

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Differences in Sociodemographic and Clinical Variables between Groups
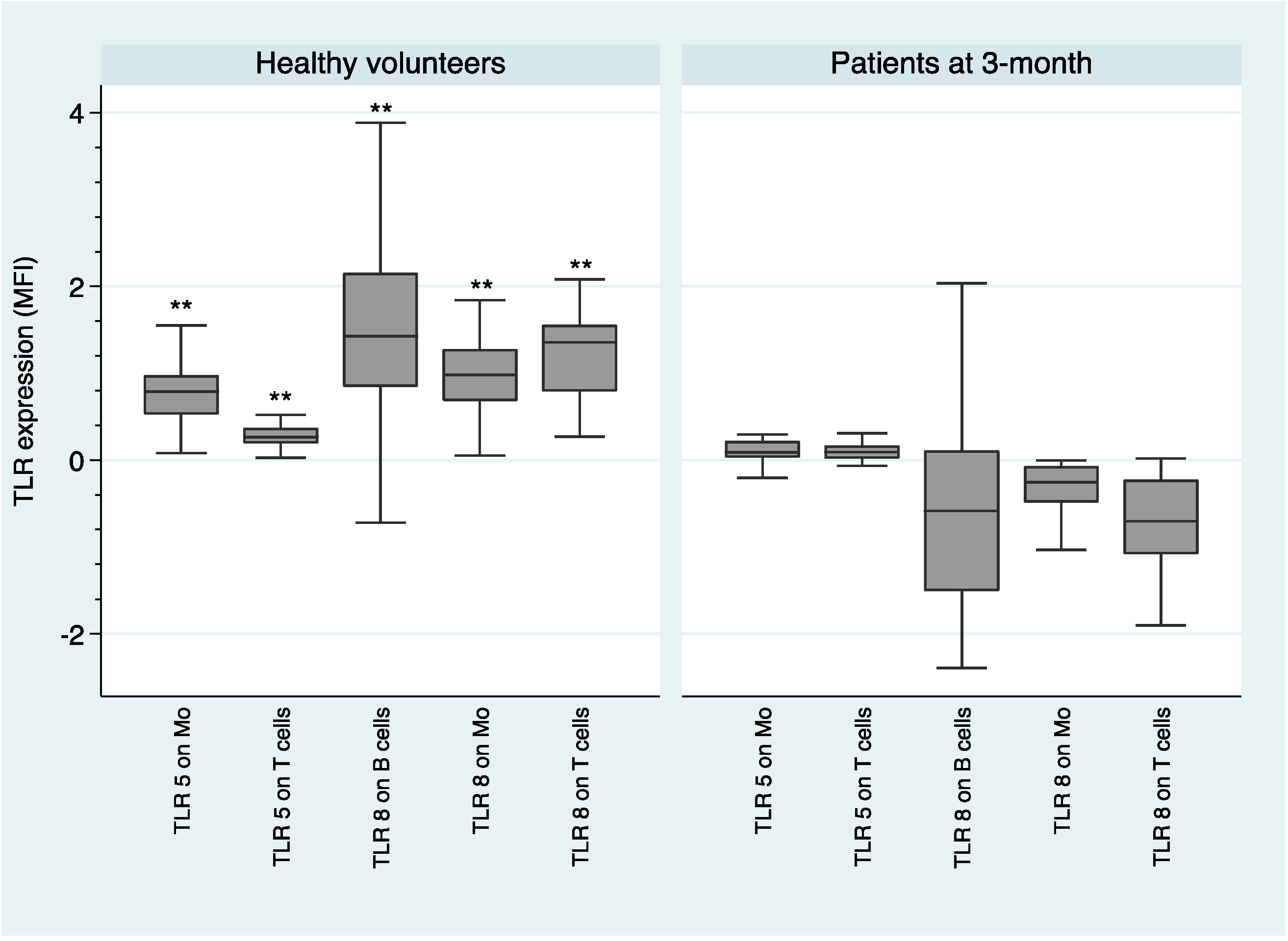
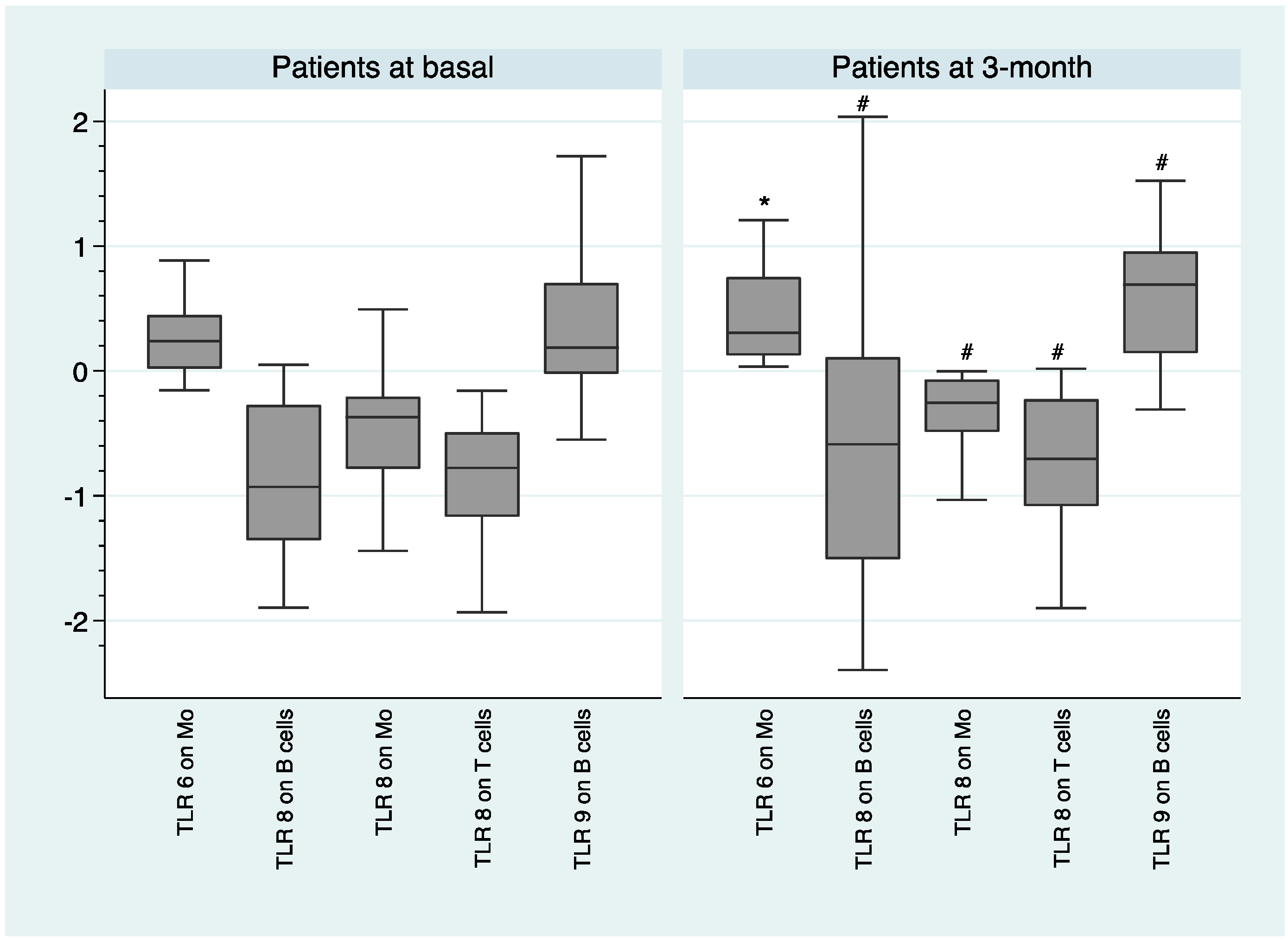
2.2. Comparison of TLR Expression Healthy Volunteers and Patients
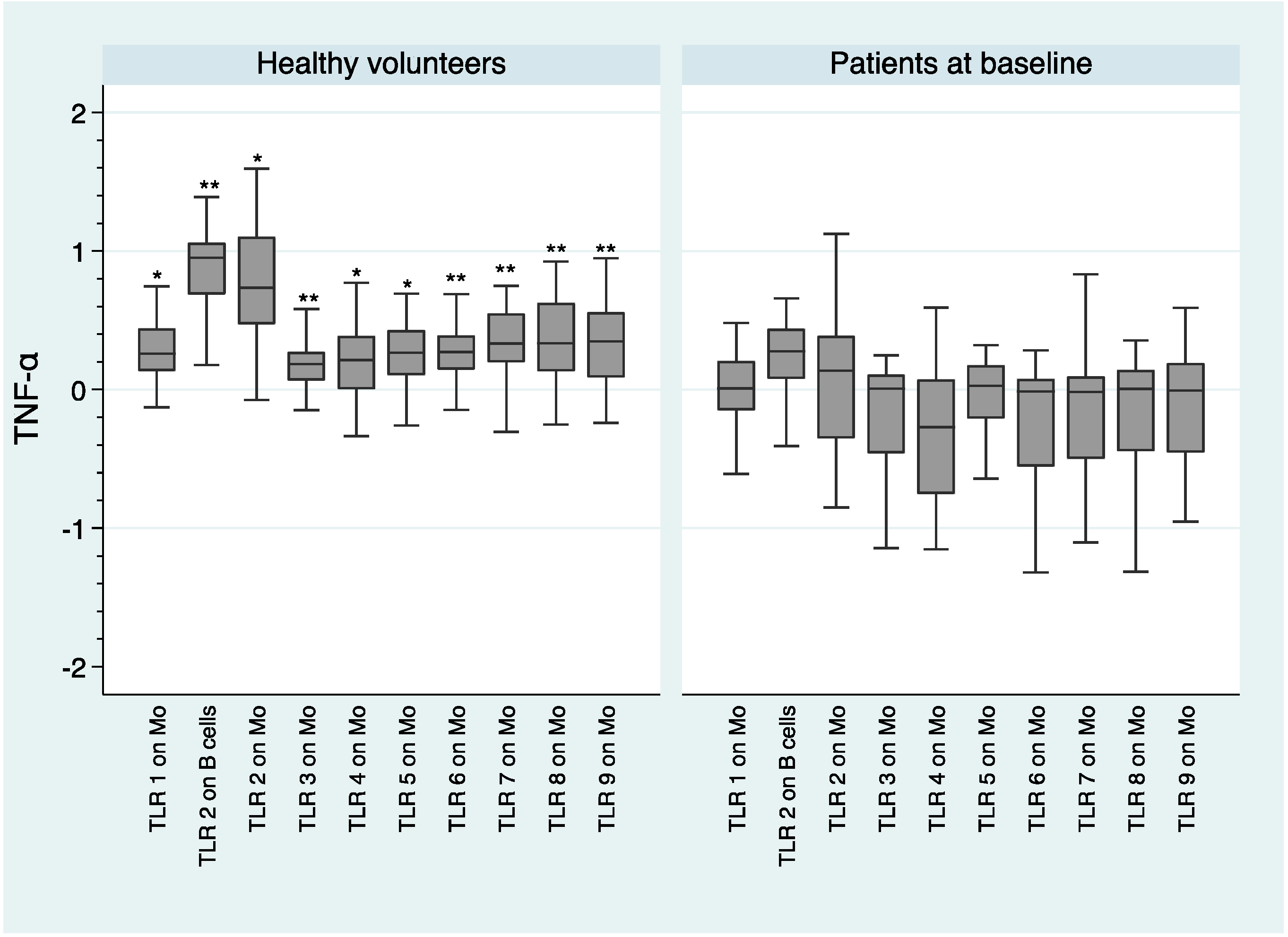
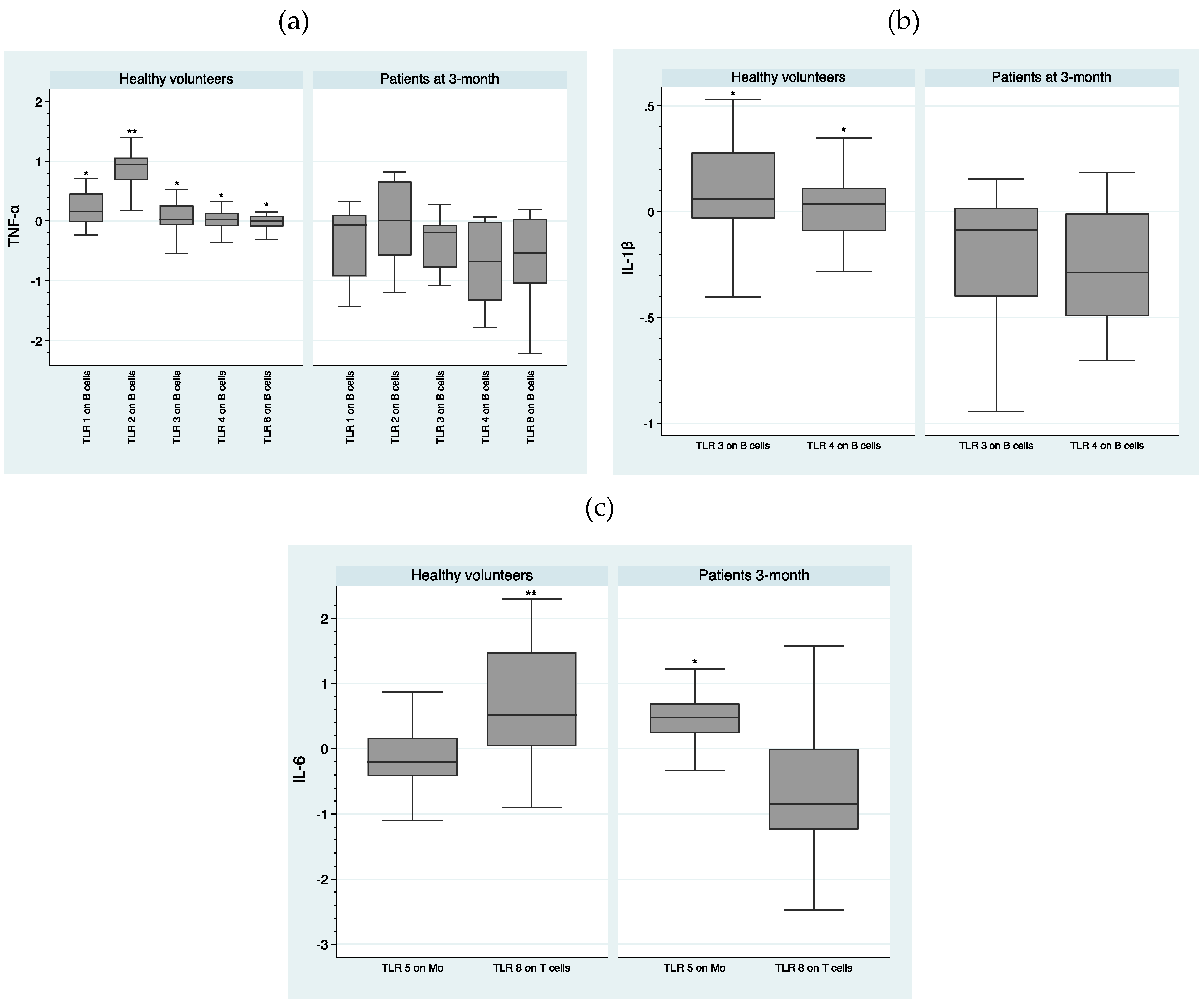
2.3. Comparison of IL-1β, IL-6, and TNF-α Intracellular Concentrations Following TLR Stimulation between Healthy Volunteers and Patients
3. Discussion
4. Materials and Methods
4.1. Study Setting
4.2. Subjects
4.3. Study Design
4.4. Assessments
4.4.1. Expression of TLRs in PBMCs
4.4.2. Assessment of TLR Functionality in Circulation PBMCs
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | Body mass index |
| PBMC | peripheral blood mononuclear cells |
| CNS | Central nervous system |
| FEP | First-episode psychosis |
| PAFIP | Programa de Atención a Fases Iniciales de Psicosis |
| TLR | Toll-like receptors |
References
- Menninger, W.C. The role of psychiatry in the world today. Am. J. Psychiatry 1994, 151, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.M.; Lewis, S.W. Is schizophrenia a neurodevelopmental disorder? Br. Med. J. 1987, 295, 681–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberger, D.R. Implications of normal brain development for the pathogenesis of schizophrenia. Arch. Gen. Psychiatry 1987, 44, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidya, M.K.; Kumar, V.G.; Sejian, V.; Bagath, M.; Krishnan, G.; Bhatta, R. Toll-like receptors: Significance, ligands, signaling pathways, and functions in mammals. Int. Rev. Immunol. 2018, 37, 20–36. [Google Scholar] [CrossRef]
- Bowman, C.C.; Rasley, A.; Tranguch, S.L.; Marriott, I. Cultured astrocytes express toll-like receptors for bacterial products. Glia 2003, 43, 281–291. [Google Scholar] [CrossRef]
- Kumar, V. Toll-like receptors in the pathogenesis of neuroinflammation. J. Neuroimmunol. 2019, 332, 16–30. [Google Scholar] [CrossRef]
- Crespo-Facorro, B.; Carrasco-Marin, E.; Perez-Iglesias, R.; Pelayo-Terán, J.M.; Fernandez-Prieto, L.; Leyva-Cobian, F.; Vázquez-Barquero, J.L. Interleukin-12 plasma levels in drug-naïve patients with a first episode of psychosis: Effects of antipsychotic drugs. Psychiatry Res. 2008, 158, 206–216. [Google Scholar] [CrossRef]
- Petrikis, P.; Voulgari, P.V.; Tzallas, A.T.; Boumba, V.A.; Archimandriti, D.T.; Zambetas, D.; Papadopoulos, I.; Tsoulos, I.; Skapinakis, P.; Mavreas, V. Changes in the cytokine profile in first-episode, drug-naïve patients with psychosis after short-term antipsychotic treatment. Psychiatry Res. 2017, 256, 378–383. [Google Scholar] [CrossRef]
- Juncal-Ruiz, M.; Riesco-Dávila, L.; De La Foz, V.O.-G.; Martínez-García, O.; Ramirez-Bonilla, M.; Ocejo-Viñals, J.G.; Leza, J.C.; López-Hoyos, M.; Crespo-Facorro, B. Comparison of the anti-inflammatory effect of aripiprazole and risperidone in 75 drug-naïve first episode psychosis individuals: A 3 months randomized study. Schizophr. Res. 2018, 202, 226–233. [Google Scholar] [CrossRef]
- Miller, B.J.; Buckley, P.; Seabolt, W.; Mellor, A.; Kirkpatrick, B. Meta-Analysis of Cytokine Alterations in Schizophrenia: Clinical Status and Antipsychotic Effects. Boil. Psychiatry 2011, 70, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Capuzzi, E.; Bartoli, F.; Crocamo, C.; Clerici, M.; Carrà, G. Acute variations of cytokine levels after antipsychotic treatment in drug-naïve subjects with a first-episode psychosis: A meta-analysis. Neurosci. Biobehav. Rev. 2017, 77, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Tangerås, L.H.; Stødle, G.S.; Olsen, G.D.; Leknes, A.-H.; Gundersen, A.S.; Skei, B.; Vikdal, A.J.; Ryan, L.; Steinkjer, B.; Myklebost, M.F.; et al. Functional Toll-like receptors in primary first-trimester trophoblasts. J. Reprod. Immunol. 2014, 106, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pudney, J.; He, X.; Masheeb, Z.; Kindelberger, D.W.; Kuohung, W.; Ingalls, R.R. Differential expression of toll-like receptors in the human placenta across early gestation. Placenta 2016, 46, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Olmos-Ortiz, A.; Flores-Espinosa, P.; Mancilla-Herrera, I.; Vega-Sánchez, R.; Díaz, L.; Zaga-Clavellina, V. Innate Immune Cells and Toll-like Receptor–Dependent Responses at the Maternal–Fetal Interface. Int. J. Mol. Sci. 2019, 20, 3654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monji, A.; Kato, T.A.; Kanba, S. Cytokines and schizophrenia: Microglia hypothesis of schizophrenia. Psychiatry Clin. Neurosci. 2009, 63, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Kahn, R.S.; Sommer, I.E. The Nerubiology and Treatment of First-Episode Schizophrenia. Mol. Psychiatry 2015, 20, 84–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinney, D.K.; Hintz, K.; Shearer, E.M.; Barch, D.H.; Riffin, C.; Whitley, K.; Butler, R. A unifying hypothesis of schizophrenia: Abnormal immune system development may help explain roles of prenatal hazards, post-pubertal onset, stress, genes, climate, infections, and brain dysfunction. Med. Hypotheses 2010, 74, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, E.; Agier, J.; Wysokiński, A.; Łucka, A.; Sobierajska, K.; Brzezińska-Błaszczyk, E. The expression of toll-like receptors in peripheral blood mononuclear cells is altered in schizophrenia. Psychiatry Res. Neuroimaging 2019, 272, 540–550. [Google Scholar] [CrossRef]
- Kanchanatawan, B.; Sriswasdi, S.; Thika, S.; Sirivichayakul, S.; Carvalho, A.F.; Geffard, M.; Kubera, M.; Maes, M. Deficit schizophrenia is a discrete diagnostic category defined by neuro-immune and neurocognitive features: Results of supervised machine learning. Metab. Brain Dis. 2018, 33, 1053–1067. [Google Scholar] [CrossRef]
- McKernan, D.P.; Dennison, U.; Gaszner, G.; Cryan, J.F.; Dinan, T.G. Enhanced peripheral toll-like receptor responses in psychosis: Further evidence of a pro-inflammatory phenotype. Transl. Psychiatry 2011, 1, e36. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Wagner, J.K.; Krause, D.; Weidinger, E.; Wildenauer, A.; Obermeier, M.; Dehning, S.; Gruber, R.; Schwarz, M.J. Impaired monocyte activation in schizophrenia. Psychiatry Res. 2012, 198, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-H.; Chiang, S.-Y.; Chiu, C.-C.; Tsai, C.-C.; Tsai, H.-H.; Huang, C.-Y.; Hsu, T.-C.; Tzang, B.-S. Expression of anti-cardiolipin antibodies and inflammatory associated factors in patients with schizophrenia. Psychiatry Res. 2011, 187, 341–346. [Google Scholar] [CrossRef]
- García-Bueno, B.; Gassó, P.; MacDowell, K.S.; Callado, L.F.; Mas, S.; Bernardo, M.; Lafuente, A.; Meana, J.J.; Leza, J.C. Evidence of activation of the Toll-like receptor-4 proinflammatory pathway in patients with schizophrenia. J. Psychiatry Neurosci. 2016, 41, E46–E55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kéri, S.; Szabó, C.; Kelemen, O. Antipsychotics influence Toll-like receptor (TLR) expression and its relationship with cognitive functions in schizophrenia. Brain Behav. Immun. 2017, 62, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Benros, M.E.; Mortensen, P.B. Role of Infection, Autoimmunity, Atopic Disorders, and the Immune System in Schizophrenia: Evidence from Epidemiological and Genetic Studies. Curr. Top. Behav. Neurosci. 2019, 44, 141–159. [Google Scholar] [CrossRef]
- Hannestad, J.; DellaGioia, N.; Bloch, M. The Effect of Antidepressant Medication Treatment on Serum Levels of Inflammatory Cytokines: A Meta-Analysis. Neuropsychopharmacology 2011, 36, 2452–2459. [Google Scholar] [CrossRef]
- Fulde, M.; Sommer, F.; Chassaing, B.; Van Vorst, K.; Dupont, A.; Hensel, M.; Basic, M.; Klopfleisch, R.; Rosenstiel, P.; Bleich, A.; et al. Neonatal selection by Toll-like receptor 5 influences long-term gut microbiota composition. Nature 2018, 560, 489–493. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Aitken, J.D.; Vijay-Kumar, M.; Gewirtz, A.T. Toll-Like Receptor–Gut Microbiota Interactions: Perturb at Your Own Risk! Annu. Rev. Physiol. 2012, 74, 177–198. [Google Scholar] [CrossRef]
- Kelly, J.R.; Minuto, C.; Cryan, J.F.; Clarke, G.; Dinan, T.G. Cross Talk: The Microbiota and Neurodevelopmental Disorders. Front. Mol. Neurosci. 2017, 11, 490. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, E.; Maukonen, J.; Hyytiäinen, T.; Kieseppä, T.; Orešič, M.; Sabunciyan, S.; Mantere, O.; Saarela, M.; Yolken, R.; Suvisaari, J.M. Analysis of microbiota in first episode psychosis identifies preliminary associations with symptom severity and treatment response. Schizophr. Res. 2018, 192, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, B.I.; McIntosh, G.; Weich, S.; Singh, S.; Rees, K. The association between first-episode psychosis and abnormal glycaemic control: Systematic review and meta-analysis. Lancet Psychiatry 2016, 3, 1049–1058. [Google Scholar] [CrossRef] [Green Version]
- Pillinger, T.; Beck, K.; Gobjila, C.; Donocik, J.G.; Jauhar, S.; Howes, O.D. Impaired Glucose Homeostasis in First-Episode Schizophrenia: A Systematic Review and Meta-analysis. JAMA Psychiatry 2017, 74, 261–269. [Google Scholar] [CrossRef]
- Tomasik, J.; Lago, S.G.; Vázquez-Bourgon, J.; Papiol, S.; Suárez-Pinilla, P.; Crespo-Facorro, B.; Bahn, S. Association of Insulin Resistance with Schizophrenia Polygenic Risk Score and Response to Antipsychotic Treatment. JAMA Psychiatry 2019, 76, 864–867. [Google Scholar] [CrossRef]
- Sutmuller, R.; Garritsen, A.; Adema, G.J. Regulatory T cells and toll-like receptors: Regulating the regulators. Ann. Rheum. Dis. 2007, 66, iii91–iii95. [Google Scholar] [CrossRef] [PubMed]
- DeMaria, O.; Pagni, P.P.; Traub, S.; De Gassart, A.; Branzk, N.; Murphy, A.; Valenzuela, D.M.; Yancopoulos, G.D.; Flavell, R.A.; Alexopoulou, L. TLR8 deficiency leads to autoimmunity in mice. J. Clin. Investig. 2010, 120, 3651–3662. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.L.; Manzin-Lorenzi, C.; Santiago-Raber, M.-L. Toll-like receptor 8 deletion accelerates autoimmunity in a mouse model of lupus through a Toll-like receptor 7-dependent mechanism. Immunology 2015, 145, 60–70. [Google Scholar] [CrossRef]
- Rawlings, D.J.; Metzler, G.; Wray-Dutra, M.; Jackson, S.W. Altered B cell signalling in autoimmunity. Nat. Rev. Immunol. 2017, 17, 421–436. [Google Scholar] [CrossRef] [Green Version]
- Benros, M.E.; Nielsen, P.R.; Nordentoft, M.; Eaton, W.W.; Dalton, S.O.; Mortensen, P.B. Autoinmmune Diseases and Severe Infections as Risk Factors for Schizophrenia: A 30-Year Population-Based Register Study. Am. J. Psychiatry 2011, 168, 1303–1310. [Google Scholar] [CrossRef]
- Jeppesen, R.; Benros, M.E. Autoimmune Diseases and Psychotic Disorders. Front. Psychol. 2019, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Khandaker, G.M.; Zimbron, J.; Lewis, G.; Jones, P.B. Prenatal maternal infection, neurodevelopment and adult schizophrenia: A systematic review of population-based studies. Psychol Med. 2013, 43, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leza, J.C.; García-Bueno, B.; Bioque, M.; Arango, C.; Parellada, M.; Do, K.; O’Donnell, P.; Bernardo, M. Inflammation in schizophrenia: A question of balance. Neurosci. Biobehav. Rev. 2015, 55, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Pelayo-Terán, J.M.; Pérez-Iglesias, R.; Ramirez-Bonilla, M.; González-Blanch, C.; Martínez-Garcia, O.; Pardo-García, G.; Rodríguez-Sánchez, J.M.; Roiz-Santiañez, R.; Tordesillas-Gutierrez, D.; Mata, I.; et al. Epidemiological factors associated with treated incidence of first-episode non-affective psychosis in Cantabria: Insights from the Clinical Programme on Early Phases of Psychosis. Early Interv. Psychiatry 2008, 2, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Rodriguez, L.; López-Hoyos, M.; Garcia-Unzueta, M.; Amado, J.A.; Cacho, P.M.; Martínez-Taboada, V.M. Age and low levels of circulating vitamin D are associated with impaired innate immune function. J. Leukoc. Boil. 2012, 91, 829–838. [Google Scholar] [CrossRef]
- Juncal-Ruiz, M.; Riesco-Dávila, L.; De La Foz, V.O.-G.; Ramirez-Bonilla, M.; Martínez-García, O.; Irure-Ventura, J.; Leza, J.C.; Lopez-Hoyos, M.; Crespo-Facorro, B. The effect of excess weight on circulating inflammatory cytokines in drug-naïve first-episode psychosis individuals. J. Neuroinflamm. 2018, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entire Simple (n = 59), n (%) | Healthy Volunteers Group (n = 26), n (%) | Patients Group, (n = 33), n (%) | Statistics | |
|---|---|---|---|---|
| Sex (Male) | 37 (62.7) | 19 (73.1) | 18 (54.6) | Χ2 = 2.14; p = 0.144 (1) |
| Cannabis Users | 27 (46.6) | 12 (46.2) | 15 (46.9) | Χ2 = 0.00; p = 0.956 (1) |
| Tobacco Users | 30 (53.6) | 15 (57.7) | 15 (50) | Χ2 = 033; p = 0.565 (1) |
| Baseline Treatment (Aripiprazole) | 19 (57.6) | z = 0.87; p = 0.384 (3) | ||
| Mean (SD) | Mean (SD) | Mean (SD) | Statistics | |
| Age at Onset (Years) * | 29.9 (7.86) | 29.6 (6.30) | 30.2 (8.99) | t = 0.25; p = 0.803 (1) |
| BMI (kg/m2) | 23.7 (3.94) | 24.6 (4.16) | 23 (3.69) (a) | t = −1.52; p = 0.135 (1) |
| 25.3 (3.94) (b) | t = 3.71; p = 0.002 (2) | |||
| Total BPRS score | 75.3 (19.0) (a) | U = −3.52; p = 0.000 (2) | ||
| 36.9 (15.59) (b) | ||||
| Total SAPS Score | 17 (5.17) (a) | U = −3.52; p = 0.000 (2) | ||
| 1.94 (3.86) (b) | ||||
| Total SANS Score | 5.7 (5.42) (a) | U = 0.89; p = 0.376 (2) | ||
| 5.8 (6.47) (b) | ||||
| CGI | 6.8 (0.44) (a) | U = −3.56; p = 0.000 (2) | ||
| 2.6 (1.84) (b) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juncal-Ruiz, M.; Riesco-Davila, L.; Vazquez-Bourgon, J.; Ortiz-Garcia de la Foz, V.; Mayoral-Van Son, J.; Ayesa-Arriola, R.; Setien-Suero, E.; Leza, J.C.; Lopez-Hoyos, M.; Crespo-Facorro, B. Expression and Functionality Study of 9 Toll-Like Receptors in 33 Drug-Naïve Non-Affective First Episode Psychosis Individuals: A 3-Month Study. Int. J. Mol. Sci. 2020, 21, 6106. https://doi.org/10.3390/ijms21176106
Juncal-Ruiz M, Riesco-Davila L, Vazquez-Bourgon J, Ortiz-Garcia de la Foz V, Mayoral-Van Son J, Ayesa-Arriola R, Setien-Suero E, Leza JC, Lopez-Hoyos M, Crespo-Facorro B. Expression and Functionality Study of 9 Toll-Like Receptors in 33 Drug-Naïve Non-Affective First Episode Psychosis Individuals: A 3-Month Study. International Journal of Molecular Sciences. 2020; 21(17):6106. https://doi.org/10.3390/ijms21176106
Chicago/Turabian StyleJuncal-Ruiz, Maria, Laura Riesco-Davila, Javier Vazquez-Bourgon, Victor Ortiz-Garcia de la Foz, Jacqueline Mayoral-Van Son, Rosa Ayesa-Arriola, Esther Setien-Suero, Juan Carlos Leza, Marcos Lopez-Hoyos, and Benedicto Crespo-Facorro. 2020. "Expression and Functionality Study of 9 Toll-Like Receptors in 33 Drug-Naïve Non-Affective First Episode Psychosis Individuals: A 3-Month Study" International Journal of Molecular Sciences 21, no. 17: 6106. https://doi.org/10.3390/ijms21176106
APA StyleJuncal-Ruiz, M., Riesco-Davila, L., Vazquez-Bourgon, J., Ortiz-Garcia de la Foz, V., Mayoral-Van Son, J., Ayesa-Arriola, R., Setien-Suero, E., Leza, J. C., Lopez-Hoyos, M., & Crespo-Facorro, B. (2020). Expression and Functionality Study of 9 Toll-Like Receptors in 33 Drug-Naïve Non-Affective First Episode Psychosis Individuals: A 3-Month Study. International Journal of Molecular Sciences, 21(17), 6106. https://doi.org/10.3390/ijms21176106