Exhaustive Search of the Receptor Ligands by the CyCLOPS (Cytometry Cell-Labeling Operable Phage Screening) Technique

 , and
, and 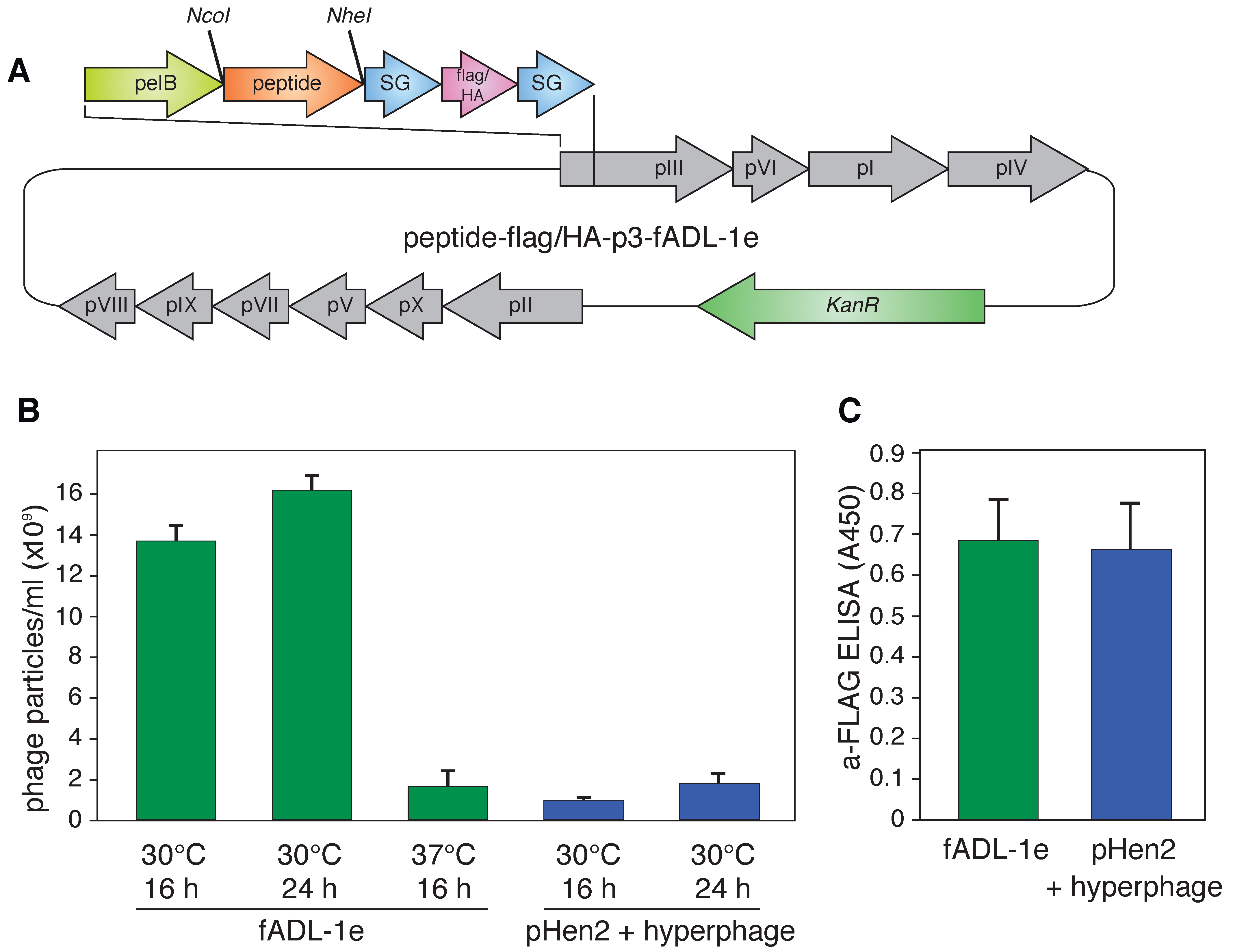{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Engineering of the fADL-1e Phage Vector for the Efficient Production of Bacteriophages Carrying Exposed Ligands
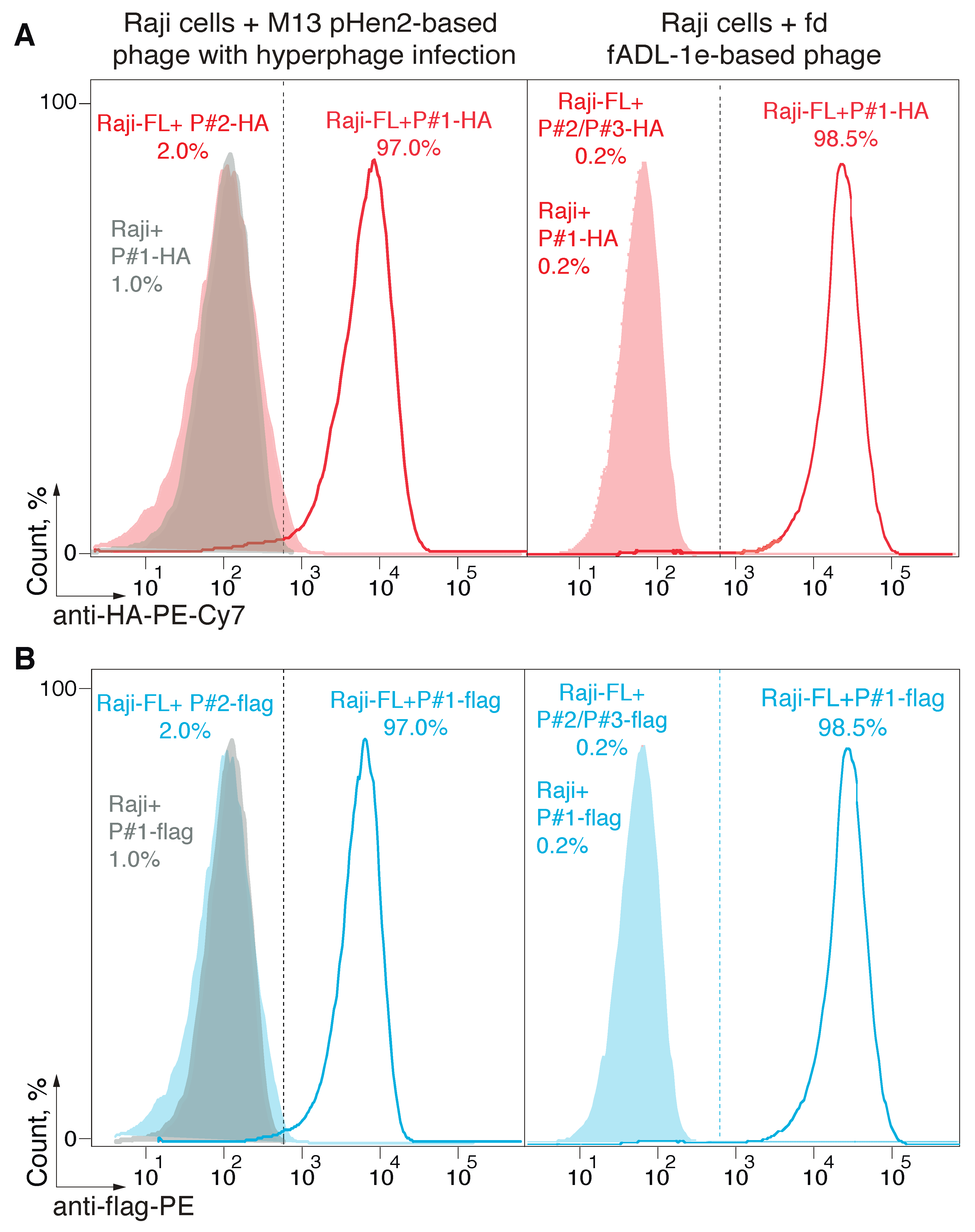
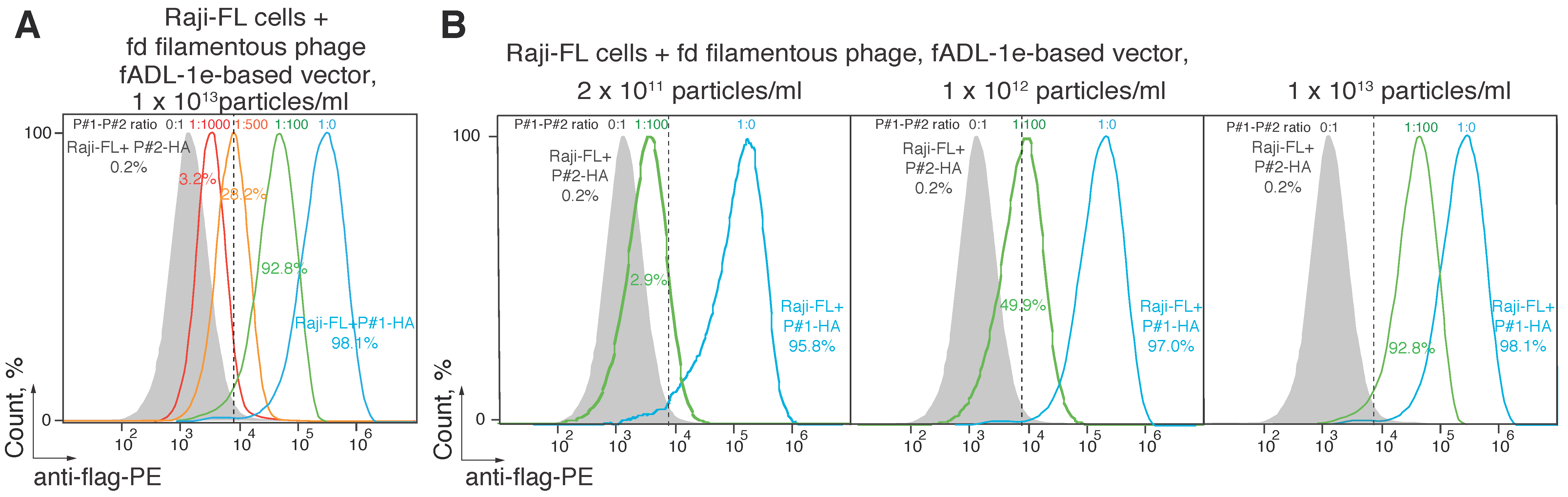
2.2. Utilization of the fADL-1e-Based Protocol Significantly Decreases False-Positive Staining of the Antigen-Specific B Cells
2.3. CyCLOPS Is Configured to Simultaneously Analyze up to the Ten Thousand Individual Antigens
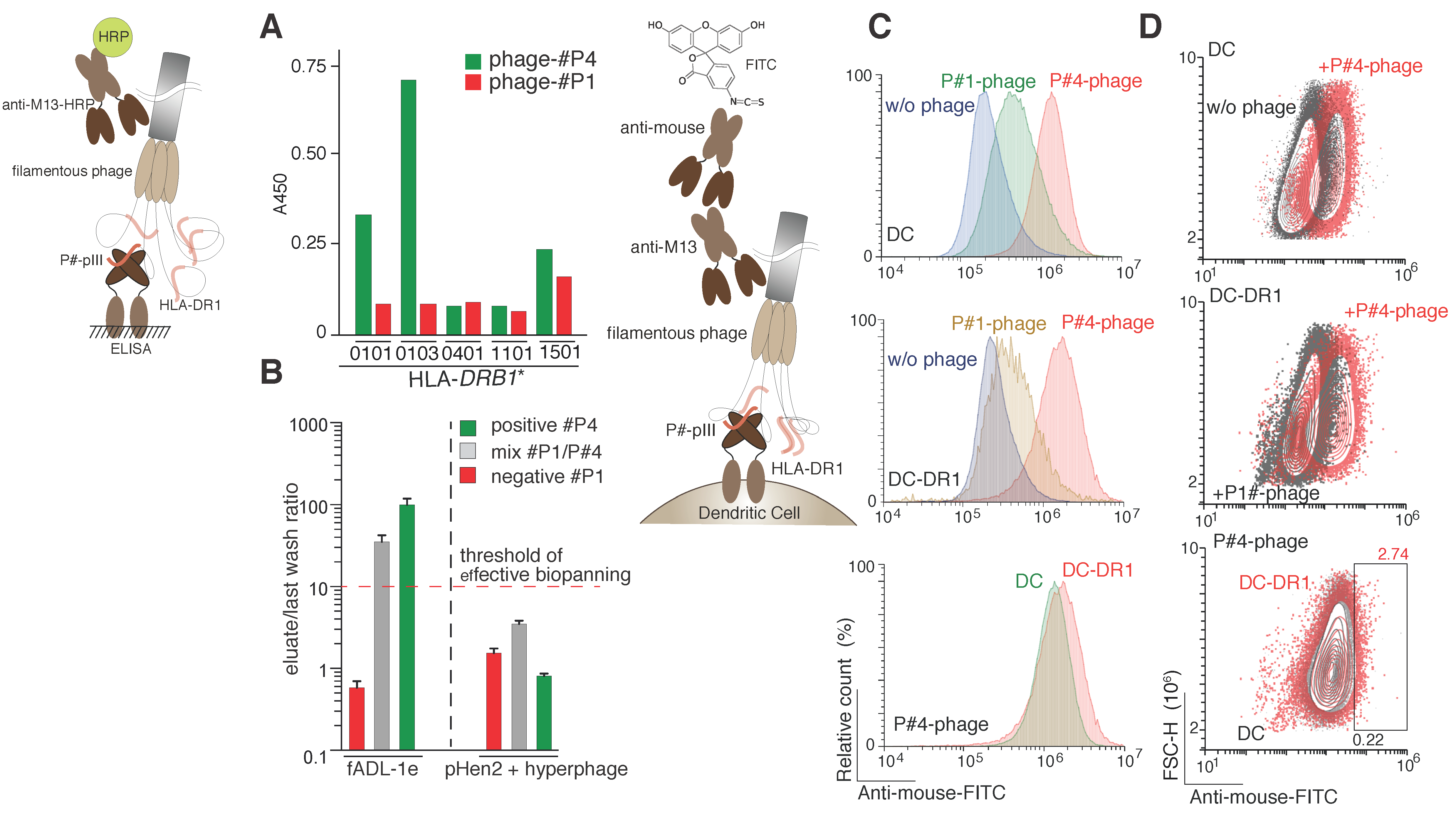
2.4. CyCLOPS May Be Adopted for the Selection of the HLA Class II Ligands
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Plasmids
4.3. Expression and Purification of Recombinant Bacteriophages
4.4. ELISA
4.5. Staining of Antigen-Specific Cells with Bacteriophages Followed by Flow Cytometry
4.6. Enrichment and Sequencing of Antigen Ligands Exposed on Bacteriophage Surface
4.7. HLA Preparation
4.8. Selection of Peptide Substrates on HLA II by Phage Display
4.9. Binding Assay of Dendritic Cells, Expressing Recombinant DR1 Molecules with Bacteriophages, Exposing HLA II-Ligands by FACS
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, B.; DeKosky, B.J.; Timm, M.R.; Lee, J.; Normandin, E.; Misasi, J.; Kong, R.; McDaniel, J.R.; Delidakis, G.; Leigh, K.E.; et al. Functional interrogation and mining of natively paired human VH:VL antibody repertoires. Nat. Biotechnol. 2018, 36, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.S.; Mizrahi, R.A.; Spindler, M.J.; Adams, M.S.; Asensio, M.A.; Edgar, R.C.; Leong, J.; Leong, R.; Johnson, D.S. Rare, high-affinity mouse anti-PD-1 antibodies that function in checkpoint blockade, discovered using microfluidics and molecular genomics. mAbs 2017, 9, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.S.; Mizrahi, R.A.; Spindler, M.J.; Adams, M.S.; Asensio, M.A.; Edgar, R.C.; Leong, J.; Leong, R.; Roalfe, L.; White, R.; et al. Rare, high-affinity anti-pathogen antibodies from human repertoires, discovered using microfluidics and molecular genomics. mAbs 2017, 9, 1282–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, L.D.; Chen, Y.J.; Wu, J.; Chaudhuri, S.; Hsiao, Y.C.; Schneider, K.; Hoi, K.H.; Lin, Z.; Guerrero, S.; Jaiswal, B.S.; et al. Massively parallel single-cell B-cell receptor sequencing enables rapid discovery of diverse antigen-reactive antibodies. Commun. Biol. 2019, 2, 304. [Google Scholar] [CrossRef] [Green Version]
- Setliff, I.; Shiakolas, A.R.; Pilewski, K.A.; Murji, A.A.; Mapengo, R.E.; Janowska, K.; Richardson, S.; Oosthuysen, C.; Raju, N.; Ronsard, L.; et al. High-Throughput Mapping of B Cell Receptor Sequences to Antigen Specificity. Cell 2019, 179, 1636–1646.e15. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, M.Y.; Belyanina, T.A.; Sokolov, A.V.; Kiselev, I.S.; Mamedov, A.E. The Contribution of Major Histocompatibility Complex Class II Genes to an Association with Autoimmune Diseases. Acta Nat. 2019, 11, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.K.; Andreatta, M.; Marcatili, P.; Buus, S.; Greenbaum, J.A.; Yan, Z.; Sette, A.; Peters, B.; Nielsen, M. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology 2018, 154, 394–406. [Google Scholar] [CrossRef]
- Wahlstrom, J.; Dengjel, J.; Persson, B.; Duyar, H.; Rammensee, H.G.; Stevanovic, S.; Eklund, A.; Weissert, R.; Grunewald, J. Identification of HLA-DR-bound peptides presented by human bronchoalveolar lavage cells in sarcoidosis. J. Clin. Investig. 2007, 117, 3576–3582. [Google Scholar] [CrossRef] [Green Version]
- Davenport, M.P.; Quinn, C.L.; Valsasnini, P.; Sinigaglia, F.; Hill, A.V.; Bell, J.I. Analysis of peptide-binding motifs for two disease associated HLA-DR13 alleles using an M13 phage display library. Immunology 1996, 88, 482–486. [Google Scholar] [CrossRef]
- Hammer, J.; Valsasnini, P.; Tolba, K.; Bolin, D.; Higelin, J.; Takacs, B.; Sinigaglia, F. Promiscuous and allele-specific anchors in HLA-DR-binding peptides. Cell 1993, 74, 197–203. [Google Scholar] [CrossRef]
- Kowalewski, D.J.; Schuster, H.; Backert, L.; Berlin, C.; Kahn, S.; Kanz, L.; Salih, H.R.; Rammensee, H.G.; Stevanovic, S.; Stickel, J.S. HLA ligandome analysis identifies the underlying specificities of spontaneous antileukemia immune responses in chronic lymphocytic leukemia (CLL). Proc. Natl. Acad. Sci. USA 2015, 112, E166–E175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Tian, T.; Liu, W.; Zhu, Z.; J. Yang, C. Advance in phage display technology for bioanalysis. Biotechnol. J. 2016, 11, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Baeriswyl, V.; Heinis, C. Phage selection of cyclic peptide antagonists with increased stability toward intestinal proteases. Protein Eng. Des. Sel. PEDS 2013, 26, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Huovinen, T.; Syrjanpaa, M.; Sanmark, H.; Seppa, T.; Akter, S.; Khan, L.M.; Lamminmaki, U. The selection performance of an antibody library displayed on filamentous phage coat proteins p9, p3 and truncated p3. BMC Res. Notes 2014, 7, 661. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, Y.; Konthur, Z. Design and screening of M13 phage display cDNA libraries. Molecules 2011, 16, 1667–1681. [Google Scholar] [CrossRef]
- Urban, J.H.; Moosmeier, M.A.; Aumüller, T.; Thein, M.; Bosma, T.; Rink, R.; Groth, K.; Zulley, M.; Siegers, K.; Tissot, K.; et al. Phage display and selection of lanthipeptides on the carboxy-terminus of the gene-3 minor coat protein. Nat. Commun. 2017, 8, 1500. [Google Scholar] [CrossRef] [Green Version]
- Bratkovic, T. Progress in phage display: Evolution of the technique and its application. Cell. Mol. Life Sci. CMLS 2010, 67, 749–767. [Google Scholar] [CrossRef]
- Hufton, S.E.; Moerkerk, P.T.; Meulemans, E.V.; de Bruine, A.; Arends, J.W.; Hoogenboom, H.R. Phage display of cDNA repertoires: The pVI display system and its applications for the selection of immunogenic ligands. J. Immunol. Methods 1999, 231, 39–51. [Google Scholar] [CrossRef]
- Gao, C.; Mao, S.; Lo, C.H.; Wirsching, P.; Lerner, R.A.; Janda, K.D. Making artificial antibodies: A format for phage display of combinatorial heterodimeric arrays. Proc. Natl. Acad. Sci. USA 1999, 96, 6025–6030. [Google Scholar] [CrossRef] [Green Version]
- Hoogenboom, H.R.; Griffiths, A.D.; Johnson, K.S.; Chiswell, D.J.; Hudson, P.; Winter, G. Multi-subunit proteins on the surface of filamentous phage: Methodologies for displaying antibody (Fab) heavy and light chains. Nucleic Acids Res. 1991, 19, 4133–4137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, J.; Messing, J. Production of single-stranded plasmid DNA. Methods Enzymol. 1987, 153, 3–11. [Google Scholar] [CrossRef]
- Rondot, S.; Koch, J.; Breitling, F.; Dubel, S. A helper phage to improve single-chain antibody presentation in phage display. Nat. Biotechnol. 2001, 19, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Jestin, J.L.; Volioti, G.; Winter, G. Improving the display of proteins on filamentous phage. Res. Microbiol. 2001, 152, 187–191. [Google Scholar] [CrossRef]
- Rakonjac, J.; Jovanovic, G.; Model, P. Filamentous phage infection-mediated gene expression: Construction and propagation of the gIII deletion mutant helper phage R408d3. Gene 1997, 198, 99–103. [Google Scholar] [CrossRef]
- Baek, H.; Suk, K.H.; Kim, Y.H.; Cha, S. An improved helper phage system for efficient isolation of specific antibody molecules in phage display. Nucleic Acids Res. 2002, 30, e18. [Google Scholar] [CrossRef] [Green Version]
- Soltes, G.; Barker, H.; Marmai, K.; Pun, E.; Yuen, A.; Wiersma, E.J. A new helper phage and phagemid vector system improves viral display of antibody Fab fragments and avoids propagation of insert-less virions. J. Immunol. Methods 2003, 274, 233–244. [Google Scholar] [CrossRef]
- Petrenko, V.A. Landscape Phage: Evolution from Phage Display to Nanobiotechnology. Viruses 2018, 10, 311. [Google Scholar] [CrossRef] [Green Version]
- Huie, M.A.; Cheung, M.C.; Muench, M.O.; Becerril, B.; Kan, Y.W.; Marks, J.D. Antibodies to human fetal erythroid cells from a nonimmune phage antibody library. Proc. Natl. Acad. Sci. USA 2001, 98, 2682–2687. [Google Scholar] [CrossRef] [Green Version]
- Lomakin, Y.A.; Kaminskaya, A.N.; Stepanov, A.V.; Shmidt, A.A.; Gabibov, A.G.; Belogurov, A.A., Jr. Probing Surface Membrane Receptors Using Engineered Bacteriophage Bioconjugates. Bioconjugate Chem. 2019, 30, 1500–1506. [Google Scholar] [CrossRef]
- Lomakin, Y.A.; Kaminskaya, A.N.; Stepanov, A.V.; Shmidt, A.A.; Gabibov, A.G.; Belogurov, A.A., Jr. High-Throughput Platform for B-Cell Screening Based on Fluorescent Phage-Display Technology. Bull. Exp. Biol. Med. 2019, 167, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Stepanov, A.V.; Markov, O.V.; Chernikov, I.V.; Gladkikh, D.V.; Zhang, H.; Jones, T.; Sen’kova, A.V.; Chernolovskaya, E.L.; Zenkova, M.A.; Kalinin, R.S.; et al. Autocrine-based selection of ligands for personalized CAR-T therapy of lymphoma. Sci. Adv. 2018, 4, eaau4580. [Google Scholar] [CrossRef] [Green Version]
- Kalandadze, A.; Galleno, M.; Foncerrada, L.; Strominger, J.L.; Wucherpfennig, K.W. Expression of recombinant HLA-DR2 molecules. Replacement of the hydrophobic transmembrane region by a leucine zipper dimerization motif allows the assembly and secretion of soluble DR alpha beta heterodimers. J. Biol. Chem. 1996, 271, 20156–20162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellins, E.D.; Stern, L.J. HLA-DM and HLA-DO, key regulators of MHC-II processing and presentation. Curr. Opin. Immunol. 2014, 26, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, L.J.; Brown, J.H.; Jardetzky, T.S.; Gorga, J.C.; Urban, R.G.; Strominger, J.L.; Wiley, D.C. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature 1994, 368, 215–221. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, D.; Arrhenius, T.; Sidney, J.; Del Guercio, M.F.; Albertson, M.; Wall, M.; Oseroff, C.; Southwood, S.; Colón, S.M.; Gaeta, F.C.; et al. On the interaction of promiscuous antigenic peptides with different DR alleles. Identification of common structural motifs. J. Immunol. 1991, 147, 2663–2669. [Google Scholar] [PubMed]
- Watanabe, N.; Suzuki, Y.; Yonezu, T.; Nakagawa, Y.; Shiina, T.; Hirayama, N.; Inokuchi, S.; Inoue, S. A cell-based high-throughput screening assay system for inhibitor compounds of antigen presentation by HLA class II molecule. Sci. Rep. 2017, 7, 6798. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680. [Google Scholar] [CrossRef]
- Zacher, A.N., 3rd; Stock, C.A.; Golden, J.W., 2nd; Smith, G.P. A new filamentous phage cloning vector: Fd-tet. Gene 1980, 9, 127–140. [Google Scholar] [CrossRef]
- Mamedov, A.; Vorobyeva, N.; Filimonova, I.; Zakharova, M.; Kiselev, I.; Bashinskaya, V.; Baulina, N.; Boyko, A.; Favorov, A.; Kulakova, O.; et al. Protective Allele for Multiple Sclerosis HLA-DRB1*01:01 Provides Kinetic Discrimination of Myelin and Exogenous Antigenic Peptides. Front. Immunol. 2019, 10, 3088. [Google Scholar] [CrossRef]
- Mamedov, A.E.; Ponomarenko, N.A.; Belogurov, A.A., Jr.; Gabibov, A.G. Erratum to: Heterodimer HLA-DM Fused with Constant Fragment of the Heavy Chain of the Human Immunoglobulin Accelerates Influenza Hemagglutinin Peptide HA306-318 Loading to HLA-DR1. Bull. Exp. Biol. Med. 2016, 161, 442–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishina, I.A.; Filimonova, I.N.; Zakharova, M.Y.; Ovchinnikova, L.A.; Mamedov, A.E.; Lomakin, Y.A.; Belogurov, A.A., Jr. Exhaustive Search of the Receptor Ligands by the CyCLOPS (Cytometry Cell-Labeling Operable Phage Screening) Technique. Int. J. Mol. Sci. 2020, 21, 6258. https://doi.org/10.3390/ijms21176258
Ishina IA, Filimonova IN, Zakharova MY, Ovchinnikova LA, Mamedov AE, Lomakin YA, Belogurov AA Jr. Exhaustive Search of the Receptor Ligands by the CyCLOPS (Cytometry Cell-Labeling Operable Phage Screening) Technique. International Journal of Molecular Sciences. 2020; 21(17):6258. https://doi.org/10.3390/ijms21176258
Chicago/Turabian StyleIshina, Irina A., Ioanna N. Filimonova, Maria Y. Zakharova, Leyla A. Ovchinnikova, Azad E. Mamedov, Yakov A. Lomakin, and Alexey A. Belogurov, Jr. 2020. "Exhaustive Search of the Receptor Ligands by the CyCLOPS (Cytometry Cell-Labeling Operable Phage Screening) Technique" International Journal of Molecular Sciences 21, no. 17: 6258. https://doi.org/10.3390/ijms21176258
APA StyleIshina, I. A., Filimonova, I. N., Zakharova, M. Y., Ovchinnikova, L. A., Mamedov, A. E., Lomakin, Y. A., & Belogurov, A. A., Jr. (2020). Exhaustive Search of the Receptor Ligands by the CyCLOPS (Cytometry Cell-Labeling Operable Phage Screening) Technique. International Journal of Molecular Sciences, 21(17), 6258. https://doi.org/10.3390/ijms21176258