Platelets Boost Recruitment of CD133+ Bone Marrow Stem Cells to Endothelium and the Rodent Liver—The Role of P-Selectin/PSGL-1 Interactions

 ,
,  ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Platelets Augment CD133+BMSC Adhesion to Human Micro-Endothelium under Shear Stress In Vitro
2.2. The Relevance of the P-Selectin/PSGL-1-Axis for the Effect of Platelets to Improve CD133+BMSC Adhesion to Human Micro-Endothelium
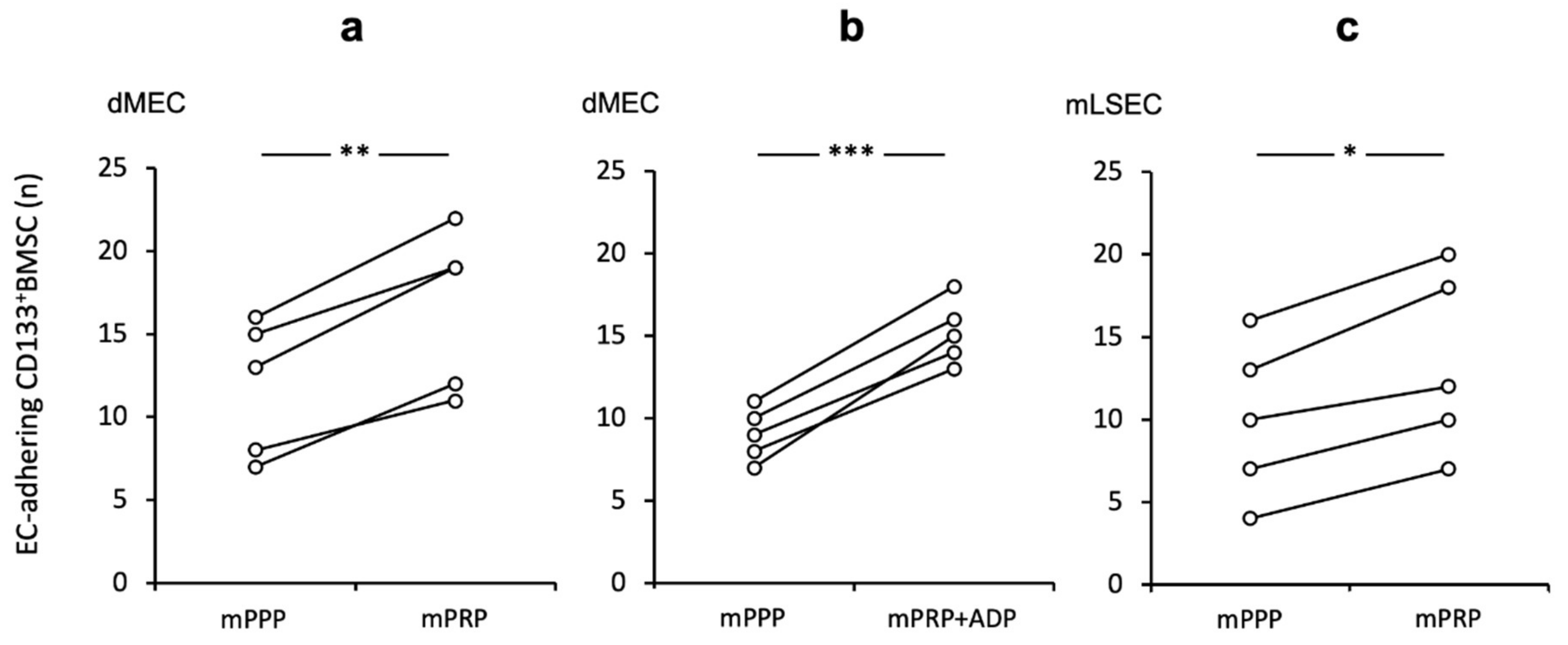
2.3. Platelet Promoting Effect In Vitro on CD133+BMSC Adhesion to Endothelium is Conserved for Rodent Micro Endothelium and LSEC Independent of Further Stimulation
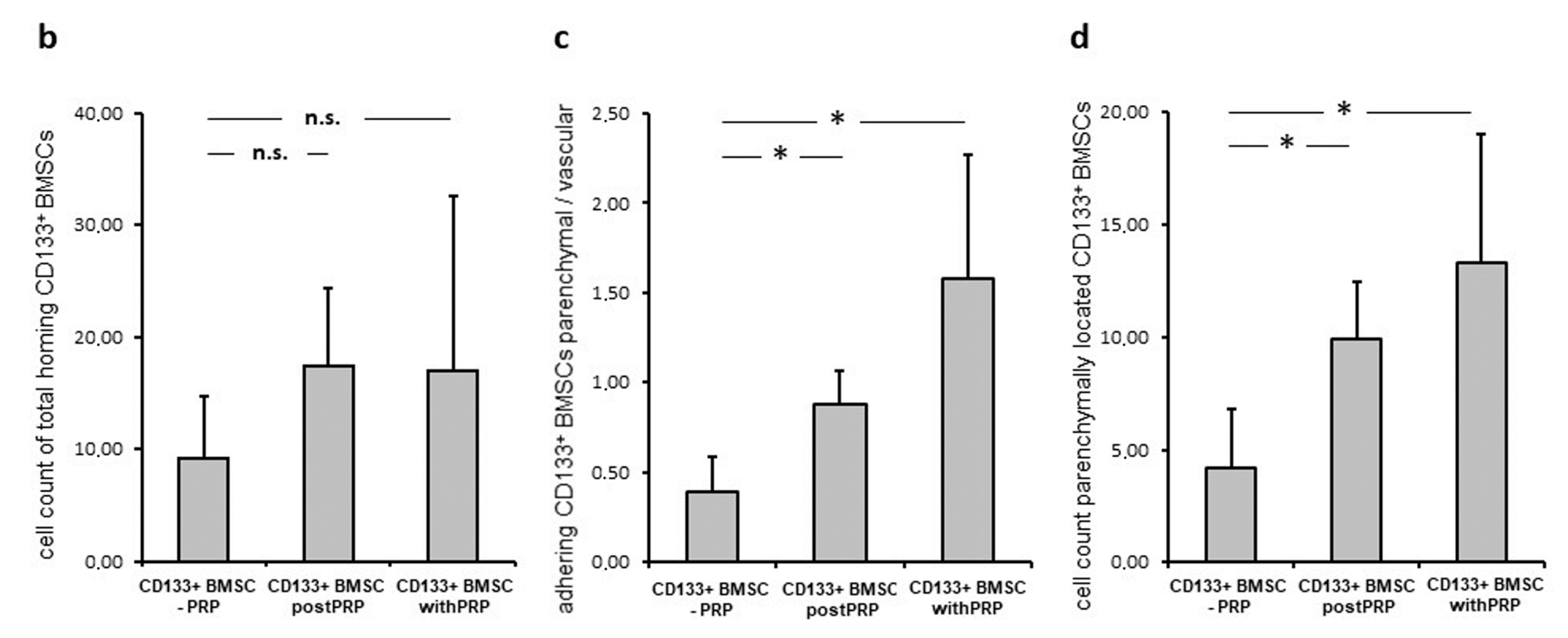
2.4. Platelets Promote Hepatic Homing and Extravasation of CD133+BMSC in the Course of Reperfusion Subsequent to Warm Ischemia
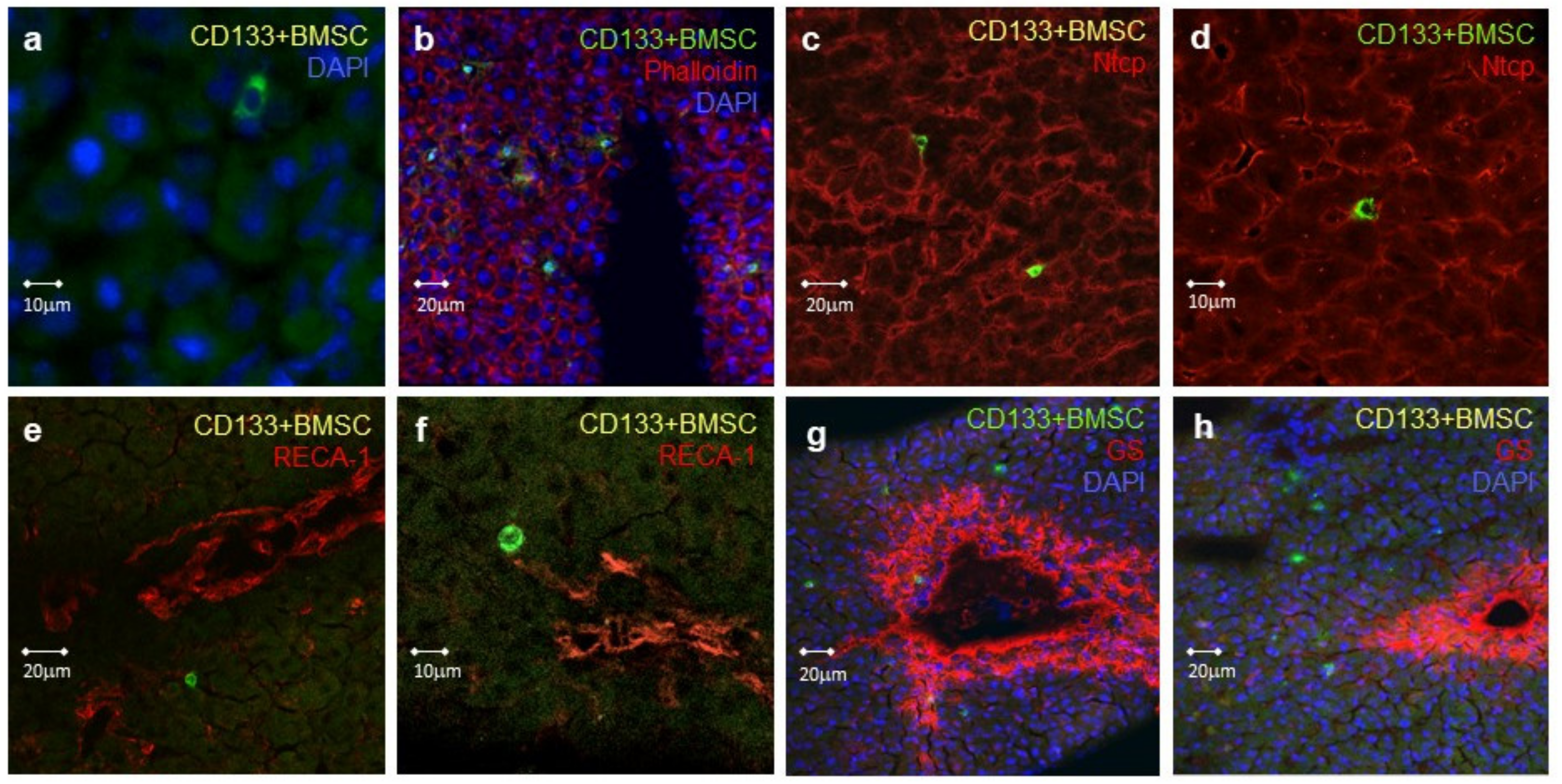
2.5. Localization of Human CD133+BMSC Homing during Hepatic Warm Ischemia Reperfusion Injury
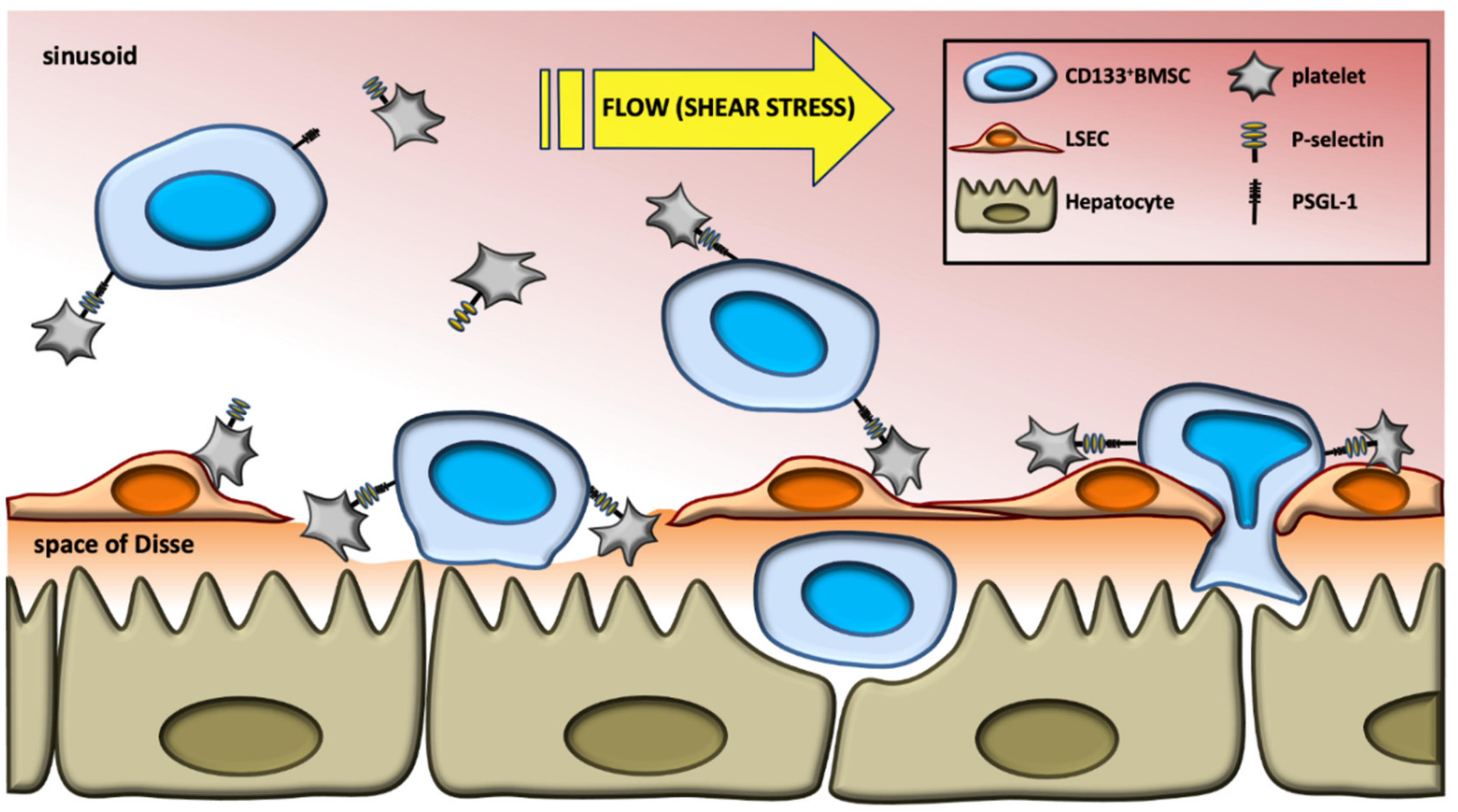
3. Discussion
4. Materials and Methods
4.1. Cells and Cell Isolation
4.2. Co-Culture Live Cell Assay
4.3. Isolated Perfused Rat Liver (IPRL)
4.4. Confocal Fluorescence Microscopy and In Situ Imaging
4.5. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMSC | bone marrow stem cells |
| dMEC | dermal microvasculature endothelial cells |
| EC | endothelial cells |
| EPC | endothelial progenitor cells |
| HMEC-1 | human dermal microvascular endothelial cells |
| h | hours |
| hPPP | human platelet poor plasma |
| hPRP | human platelet rich plasma |
| HSC | hematopoietic stem cells |
| IPRL | isolated perfused rat liver |
| LSEC | liver sinusoidal endothelial cells |
| MEC | micro-endothelial cells |
| min | minutes |
| mLSEC | mouse hepatic sinusoidal endothelial cells |
| mPPP | mouse platelet poor plasma |
| mPRP | mouse platelet rich plasma |
| n.s. | not significant |
| PECAM-1 | platelet-endothelial-adhesion-molecule-1 |
| PPP | platelet poor plasma |
| PRP | platelet rich plasma |
| PSGL-1 | P-Selectin-Glyco-Ligand-1 |
| SDF-1 | stroma-derived factor-1 |
References
- Schmelzle, M.; Duhme, C.; Junger, W.; Salhanick, S.D.; Chen, Y.; Wu, Y.; Toxavidis, V.; Csizmadia, E.; Han, L.; Bian, S.; et al. CD39 modulates hematopoietic stem cell recruitment and promotes liver regeneration in mice and humans after partial hepatectomy. Ann. Surg. 2013, 257, 693–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Am Esch, J.S.; Knoefel, W.T.; Klein, M.; Ghodsizad, A.; Fuerst, G.; Poll, L.W.; Piechaczek, C.; Burchardt, E.R.; Feifel, N.; Stoldt, V.; et al. Portal application of autologous CD133+ bone marrow cells to the liver: A novel concept to support hepatic regeneration. Stem. Cells 2005, 23, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Am Esch, J.S.; Schmelzle, M.; Fürst, G.; Robson, S.C.; Krieg, A.; Duhme, C.; Tustas, R.Y.; Alexander, A.; Klein, H.M.; Topp, S.A.; et al. Infusion of CD133+ bone marrow-derived stem cells after selective portal vein embolization enhances functional hepatic reserves after extended right hepatectomy: A retrospective single-center study. Ann. Surg. 2012, 255, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Fürst, G.; Schulte am Esch, J.; Poll, L.W.; Hosch, S.B.; Fritz, L.B.; Klein, M.; Godehardt, E.; Krieg, A.; Wecker, B.; Stoldt, V.; et al. Portal vein embolization and autologous CD133+ bone marrow stem cells for liver regeneration: Initial experience. Radiology 2007, 243, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Knoefel, W.T.; Alexander, A.; Tustas, R.Y.; Schmelzle, M.; Klein, H.M.; Krieg, A.; Topp, S.A.; Eisenberger, C.F.; Fuerst, G.; Schulte am Esch, J. Stem cell-induced liver regeneration. Zent. Chir. 2013, 138, 166–172. [Google Scholar] [CrossRef]
- Lehwald, N.; Duhme, C.; Wildner, M.; Kuhn, S.; Fürst, G.; Forbes, S.J.; Jonas, S.; Robson, S.C.; Knoefel, W.T.; Schmelzle, M.; et al. HGF and SDF-1-mediated mobilization of CD133+ BMSC for hepatic regeneration following extensive liver resection. Liver Int. Off. J. Int. Assoc. Study Liver 2014, 34, 89–101. [Google Scholar] [CrossRef]
- Oodi, A.; Nikougoftar Zarif, M.; Amani, M.; Amirizadeh, N. Expression of CXCR4 in cord blood-derived CD133+ cells treated with platelet micro-particles. Artif. Cells Nanomed. Biotechnol. 2016, 44, 1702–1707. [Google Scholar]
- Duhme, C.; Lehwald, N.; Kehrel, B.E.; Bauchrowitz, E.; Ngepi, A.; Schmelzle, M.; Kolokotronis, T.; Benhidjeb, T.; Krüger, M.; Jurk, K.; et al. CD133(+) bone marrow stem cells (BMSC) control platelet activation—Role of ectoNTPDase-1 (CD39). Blood Cells Mol. Dis. 2019, 77, 142–148. [Google Scholar] [CrossRef]
- Am Esch, J.S.; Tustas, R.Y.; Robson, S.C.; Hosch, S.B.; Akyildiz, A.; Bröring, D.C.; Fischer, L.; Knoefel, W.T.; Rogiers, X. Recipient levels and function of von Willebrand factor prior to liver transplantation and its consumption in the course of grafting correlate with hepatocellular damage and outcome. Transpl. Int. Off. J. Eur. Soc. Organ Transplant. 2005, 18, 1258–1265. [Google Scholar]
- Schulte am Esch, J.; Akyildiz, A.; Tustas, R.Y.; Ganschow, R.; Schmelzle, M.; Krieg, A.; Robson, S.C.; Topp, S.A.; Rogiers, X.; Knoefel, W.T.; et al. ADP-dependent platelet function prior to and in the early course of pediatric liver transplantation and persisting thrombocytopenia are positively correlated with ischemia/reperfusion injury. Transpl. Int. Off. J. Eur. Soc. Organ Transplant. 2010, 23, 745–752. [Google Scholar] [CrossRef]
- Esch, J.S.; Jurk, K.; Knoefel, W.T.; Roeder, G.; Voss, H.; Tustas, R.Y.; Schmelzle, M.; Krieg, A.; Eisenberger, C.F.; Topp, S.; et al. Platelet activation and increased tissue factor expression on monocytes in reperfusion injury following orthotopic liver transplantation. Platelets 2010, 21, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Lesurtel, M.; Graf, R.; Aleil, B.; Walther, D.J.; Tian, Y.; Jochum, W.; Gachet, C.; Bader, M.; Clavien, P.A. Platelet-derived serotonin mediates liver regeneration. Science 2006, 312, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Nocito, A.; Georgiev, P.; Dahm, F.; Jochum, W.; Bader, M.; Graf, R.; Clavien, P.A. Platelets and platelet-derived serotonin promote tissue repair after normothermic hepatic ischemia in mice. Hepatology 2007, 45, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Adams, D.H.; Watson, S.P.; Lalor, P.F. Platelets: No longer bystanders in liver disease. Hepatology 2016, 64, 1774–1784. [Google Scholar] [CrossRef]
- Murata, S.; Ohkohchi, N.; Matsuo, R.; Ikeda, O.; Myronovych, A.; Hoshi, R. Platelets promote liver regeneration in early period after hepatectomy in mice. World J. Surg. 2007, 31, 808–816. [Google Scholar] [CrossRef]
- Myronovych, A.; Murata, S.; Chiba, M.; Matsuo, R.; Ikeda, O.; Watanabe, M.; Hisakura, K.; Nakano, Y.; Kohno, K.; Kawasaki, T.; et al. Role of platelets on liver regeneration after 90% hepatectomy in mice. J. Hepatol. 2008, 49, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Alkozai, E.M.; Nijsten, M.W.; de Jong, K.P.; de Boer, M.T.; Peeters, P.M.; Slooff, M.J.; Porte, R.J.; Lisman, T. Immediate postoperative low platelet count is associated with delayed liver function recovery after partial liver resection. Ann. Surg. 2010, 251, 300–306. [Google Scholar] [CrossRef]
- Matsuo, R.; Nakano, Y.; Ohkohchi, N. Platelet administration via the portal vein promotes liver regeneration in rats after 70% hepatectomy. Ann. Surg. 2011, 253, 759–763. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Park, H.W.; Song, J.H.; Gwak, M.S.; Lee, W.J.; Kim, G.; Lee, S.K.; Ko, J.S. Association between intraoperative platelet transfusion and early graft regeneration in living donor liver transplantation. Ann. Surg. 2016, 264, 1065–1072. [Google Scholar] [CrossRef]
- Kawasaki, T.; Murata, S.; Takahashi, K.; Nozaki, R.; Ohshiro, Y.; Ikeda, N.; Pak, S.; Myronovych, A.; Hisakura, K.; Fukunaga, K.; et al. Activation of human liver sinusoidal endothelial cell by human platelets induces hepatocyte proliferation. J. Hepatol. 2010, 53, 648–654. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.; Lejmi, E.; Fontana, P.; Morel, P.; Gonelle-Gispert, C.; Buhler, L. A focus on the role of platelets in liver regeneration: Do platelet-endothelial cell interactions initiate the regenerative process? J. Hepatol. 2015, 63, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Beldi, G.; Wu, Y.; Sun, X.; Imai, M.; Enjyoji, K.; Csizmadia, E.; Candinas, D.; Erb, L.; Robson, S.C. Regulated catalysis of extracellular nucleotides by vascular CD39/ENTPD1 is required for liver regeneration. Gastroenterology 2008, 135, 1751–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, D.; Piasecki, A. Rat platelets contain growth factor(s) distinct from PDGF which stimulate DNA synthesis in primary adult rat hepatocyte cultures. Exp. Cell Res. 1984, 154, 95–100. [Google Scholar] [CrossRef]
- Strain, A.J.; McGowan, J.A.; Bucher, N.L. Stimulation of DNA synthesis in primary cultures of adult rat hepatocytes by rat platelet-associated substance(s). Vitro 1982, 18, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Lisman, T.; Porte, R.J. Mechanisms of platelet-mediated liver regeneration. Blood 2016, 128, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Slaba, I.; Wang, J.; Kolaczkowska, E.; McDonald, B.; Lee, W.Y.; Kubes, P. Imaging the dynamic platelet-neutrophil response in sterile liver injury and repair in mice. Hepatology 2015, 62, 1593–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalor, P.F.; Herbert, J.; Bicknell, R.; Adams, D.H. Hepatic sinusoidal endothelium avidly binds platelets in an integrin-dependent manner, leading to platelet and endothelial activation and leukocyte recruitment. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G469–G478. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.S.; Howell, D.N.; Steeber, D.A.; Harland, R.C.; Tedder, T.F.; Clavien, P.A. P-Selectin mediates reperfusion injury through neutrophil and platelet sequestration in the warm ischemic mouse liver. Hepatology 1999, 29, 1494–1502. [Google Scholar] [CrossRef]
- McGregor, L.; Martin, J.; McGregor, J.L. Platelet-leukocyte aggregates and derived microparticles in inflammation, vascular remodelling and thrombosis. Front. Biosci. 2006, 11, 830–837. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, M.P.A.; van Gils, J.M.; Mol, A.; Hordijk, P.L.; Zwaginga, J.J. Platelet binding to monocytes increases the adhesive properties of monocytes by up-regulating the expression and functionality of beta1 and beta2 integrins. J. Leukoc. Biol. 2006, 79, 499–507. [Google Scholar]
- Sopova, K.; Tatsidou, P.; Stellos, K. Platelets and platelet interaction with progenitor cells in vascular homeostasis and inflammation. Curr. Vasc. Pharmacol. 2012, 10, 555–562. [Google Scholar] [CrossRef] [PubMed]
- De Boer, H.C.; Verseyden, C.; Ulfman, L.H.; Zwaginga, J.J.; Bot, I.; Biessen, E.A.; Rabelink, T.J.; van Zonneveld, A.J. Fibrin and activated platelets cooperatively guide stem cells to a vascular injury and promote differentiation towards an endothelial cell phenotype. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1653–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Song, X.H.; Liu, P.; Zeng, C.L.; Huang, Z.S.; Zhu, L.J.; Jiang, Y.Z.; Ouyang, H.W.; Hu, H. Platelet-mediated mesenchymal stem cells homing to the lung reduces monocrotaline-induced rat pulmonary hypertension. Cell Transplant. 2012, 21, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Han, J.K.; Kim, B.K.; Won, J.Y.; Shin, Y.; Choi, S.B.; Hwang, I.; Kang, J.; Lee, H.J.; Koh, S.J.; Lee, J.; et al. Interaction between platelets and endothelial progenitor cells via LPA-Edg-2 axis is augmented by PPAR-delta activation. J. Mol. Cell Cardiol. 2016, 97, 266–277. [Google Scholar] [CrossRef]
- Shido, K.; Chavez, D.; Cao, Z.; Ko, J.; Rafii, S.; Ding, B.S. Platelets prime hematopoietic and vascular niche to drive angiocrine-mediated liver regeneration. Signal Transduct. Target. Ther. 2017, 2, 16044. [Google Scholar] [CrossRef]
- Benoit, M.R.; Conant, C.G.; Ionescu-Zanetti, C.; Schwartz, M.; Matin, A. New device for high-throughput viability screening of flow biofilms. Appl. Environ. Microbiol. 2010, 76, 4136–4142. [Google Scholar] [CrossRef] [Green Version]
- Thorgeirsson, S.S.; Grisham, J.W. Hematopoietic cells as hepatocyte stem cells: A critical review of the evidence. Hepatology 2006, 43, 2–8. [Google Scholar] [CrossRef]
- Grompe, M. The role of bone marrow stem cells in liver regeneration. Semin. Liver Dis. 2003, 23, 363–372. [Google Scholar]
- Almeida-Porada, G.; Porada, C.D.; Chamberlain, J.; Torabi, A.; Zanjani, E.D. Formation of human hepatocytes by human hematopoietic stem cells in sheep. Blood 2004, 104, 2582–2590. [Google Scholar] [CrossRef]
- Petersen, B.E.; Bowen, W.C.; Patrene, K.D.; Mars, W.M.; Sullivan, A.K.; Murase, N.; Boggs, S.S.; Greenberger, J.S.; Goff, J.P. Bone marrow as a potential source of hepatic oval cells. Science 1999, 284, 1168–1170. [Google Scholar] [CrossRef]
- Theise, N.D.; Nimmakayalu, M.; Gardner, R.; Illei, P.B.; Morgan, G.; Teperman, L.; Henegariu, O.; Krause, D.S. Liver from bone marrow in humans. Hepatology 2000, 32, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theise, N.D.; Badve, S.; Saxena, R.; Henegariu, O.; Sell, S.; Crawford, J.M.; Krause, D.S. Derivation of hepatocytes from bone marrow cells in mice after radiation-induced myeloablation. Hepatology 2000, 31, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Lagasse, E.; Connors, H.; Al-Dhalimy, M.; Reitsma, M.; Dohse, M.; Osborne, L.; Wang, X.; Finegold, M.; Weissman, I.L.; Grompe, M. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat. Med. 2000, 6, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Vig, P.; Russo, F.P.; Edwards, R.J.; Tadrous, P.J.; Wright, N.A.; Thomas, H.C.; Alison, M.R.; Forbes, S.J. The sources of parenchymal regeneration after chronic hepatocellular liver injury in mice. Hepatology 2006, 43, 316–324. [Google Scholar] [CrossRef]
- Willenbring, H.; Bailey, A.S.; Foster, M.; Akkari, Y.; Dorrell, C.; Olson, S.; Finegold, M.; Fleming, W.H.; Grompe, M. Myelomonocytic cells are sufficient for therapeutic cell fusion in liver. Nat. Med. 2004, 10, 744–748. [Google Scholar] [CrossRef]
- Wang, X.; Willenbring, H.; Akkari, Y.; Torimaru, Y.; Foster, M.; Al-Dhalimy, M.; Lagasse, E.; Finegold, M.; Olson, S.; Grompe, M. Cell fusion is the principal source of bone-marrow-derived hepatocytes. Nature 2003, 422, 897–901. [Google Scholar] [CrossRef]
- Harb, R.; Xie, G.; Lutzko, C.; Guo, Y.; Wang, X.; Hill, C.K.; Kanel, G.C.; DeLeve, L.D. Bone marrow progenitor cells repair rat hepatic sinusoidal endothelial cells after liver injury. Gastroenterology 2009, 137, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Forbes, S.J.; Vig, P.; Poulsom, R.; Alison, M.R.; Wright, N.A. Bone marrow-derived liver stem cells: Their therapeutic potential. Gastroenterology 2002, 123, 654–655. [Google Scholar] [CrossRef]
- Gehling, U.M.; Willems, M.; Dandri, M.; Petersen, J.; Berna, M.; Thill, M.; Wulf, T.; Müller, L.; Pollok, J.M.; Schlagner, K.; et al. Partial hepatectomy induces mobilization of a unique population of haematopoietic progenitor cells in human healthy liver donors. J. Hepatol. 2005, 43, 845–853. [Google Scholar] [CrossRef]
- Broxmeyer, H.E.; Kim, C.H.; Cooper, S.H.; Hangoc, G.; Hromas, R.; Pelus, L.M. Effects of CC, CXC, C, and CX3C chemokines on proliferation of myeloid progenitor cells, and insights into SDF-1-induced chemotaxis of progenitors. Ann. N. Y. Acad. Sci. 1999, 872, 142–162. [Google Scholar] [CrossRef]
- Broxmeyer, H.E.; Kim, C.H. Regulation of hematopoiesis in a sea of chemokine family members with a plethora of redundant activities. Exp. Hematol. 1999, 27, 1113–1123. [Google Scholar] [CrossRef]
- Hu, Z.; Evarts, R.P.; Fujio, K.; Marsden, E.R.; Thorgeirsson, S.S. Expression of hepatocyte growth factor and c-met genes during hepatic differentiation and liver development in the rat. Am. J. Pathol. 1993, 142, 1823–1830. [Google Scholar] [PubMed]
- Hatch, H.M.; Zheng, D.; Jorgensen, M.L.; Petersen, B.E. SDF-1alpha/CXCR4: A mechanism for hepatic oval cell activation and bone marrow stem cell recruitment to the injured liver of rats. Cloning Stem. Cells 2002, 4, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Dalakas, E.; Newsome, P.N.; Harrison, D.J.; Plevris, J.N. Hematopoietic stem cell trafficking in liver injury. FASEB J. 2005, 19, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Kollet, O.; Shivtiel, S.; Chen, Y.Q.; Suriawinata, J.; Thung, S.N.; Dabeva, M.D.; Kahn, J.; Spiegel, A.; Dar, A.; Samira, S.; et al. HGF, SDF-1, and MMP-9 are involved in stress-induced human CD34+ stem cell recruitment to the liver. J. Clin. Invest. 2003, 112, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Petit, I.; Szyper-Kravitz, M.; Nagler, A.; Lahav, M.; Peled, A.; Habler, L.; Kahn, J.; Spiegel, A.; Dar, A.; Samira, S.; et al. G-CSF induces stem cell mobilization by decreasing bone marrow SDF-1 and up-regulating CXCR4. Nat. Immunol. 2002, 3, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Herencia, C.; Rodriguez-Ariza, A.; Canalejo, A.; Naranjo, A.; Briceno, F.J.; Lopez-Cillero, P.; De la Mata, M.; Muñoz-Castañeda, J.R. Differential bone marrow hematopoietic stem cells mobilization in hepatectomized patients. J. Gastrointest. Surg. 2011, 15, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.; Schulte, E.J.; Schmelzle, M.; Tustas, R.Y.; El-Karmi, A.A.; Stefan, B.; Hosch, S.; Knoefel, W.T. Stem Cell Factor (SCF) levels do increase in patients subsequent to hepatectomy with the extend of parenchymal loss. Transplant. Proc. 2006, 38, 3556–3558. [Google Scholar] [CrossRef] [PubMed]
- Lev, E.I.; Estrov, Z.; Aboulfatova, K.; Harris, D.; Granada, J.F.; Alviar, C.; Kleiman, N.S.; Dong, J.F. Potential role of activated platelets in homing of human endothelial progenitor cells to subendothelial matrix. Thromb. Haemost. 2006, 96, 498–504. [Google Scholar] [CrossRef]
- Zannettino, A.C.; Berndt, M.C.; Butcher, C.; Butcher, E.C.; Vadas, M.A.; Simmons, P.J. Primitive human hematopoietic progenitors adhere to P-selectin (CD62P). Blood 1995, 85, 3466–3477. [Google Scholar] [CrossRef] [Green Version]
- Laszik, Z.; Jansen, P.J.; Cummings, R.D.; Tedder, T.F.; McEver, R.P.; Moore, K.L. P-selectin glycoprotein ligand-1 is broadly expressed in cells of myeloid, lymphoid, and dendritic lineage and in some nonhematopoietic cells. Blood 1996, 88, 3010–3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, S.M.; Gschmeissner, S.E.; Bates, P.A. PECAM-1: Its expression and function as a cell adhesion molecule on hemopoietic and endothelial cells. Leuk. Lymphoma 1995, 17, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Watt, S.M.; Williamson, J.; Genevier, H.; Fawcett, J.; Simmons, D.L.; Hatzfeld, A.; Nesbitt, S.A.; Coombe, D.R. The heparin binding PECAM-1 adhesion molecule is expressed by CD34+ hematopoietic precursor cells with early myeloid and B-lymphoid cell phenotypes. Blood 1993, 82, 2649–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albelda, S.M.; Muller, W.A.; Buck, C.A.; Newman, P.J. Molecular and cellular properties of PECAM-1 (endoCAM/CD31): A novel vascular cell-cell adhesion molecule. J. Cell Biol. 1991, 114, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, H.A.; Gadd, S.J.; Eher, R.E.; Majdic, O.T.; Schreiber, W.; Kasinrerk, W.; Strass, B.; Schnabl, E.; Knapp, W. Molecular characterization and functional analysis of the leukocyte surface protein CD31. J. Immunol. 1990, 145, 3889–3897. [Google Scholar] [PubMed]
- Liao, F.; Huynh, H.K.; Eiroa, A.; Greene, T.; Polizzi, E.; Muller, W.A. Migration of monocytes across endothelium and passage through extracellular matrix involve separate molecular domains of PECAM-1. J. Exp. Med. 1995, 182, 1337–1343. [Google Scholar] [CrossRef]
- Vaporciyan, A.A.; de Lisser, H.M.; Yan, H.C.; Jones, M.L.; Ward, P.A.; Albelda, S.M. Involvement of platelet-endothelial cell adhesion molecule-1 in neutrophil recruitment in vivo. Science 1993, 262, 1580–1582. [Google Scholar] [CrossRef]
- Yong, K.L.; Watts, M.; Shaun, T.N.; Sullivan, A.; Ings, S.; Linch, D.C. Transmigration of CD34+ cells across specialized and nonspecialized endothelium requires prior activation by growth factors and is mediated by PECAM-1 (CD31). Blood 1998, 91, 1196–1205. [Google Scholar] [CrossRef] [Green Version]
- Stellos, K.; Langer, H.; Daub, K.; Schoenberger, T.; Gauss, A.; Geisler, T.; Bigalke, B.; Mueller, I.; Schumm, M.; Schaefer, I.; et al. Platelet-derived stromal cell-derived factor-1 regulates adhesion and promotes differentiation of human CD34+ cells to endothelial progenitor cells. Circulation 2008, 117, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Massberg, S.; Konrad, I.; Schurzinger, K.; Lorenz, M.; Schneider, S.; Zohlnhoefer, D.; Hoppe, K.; Schiemann, M.; Kennerknecht, E.; Sauer, S.; et al. Platelets secrete stromal cell-derived factor 1alpha and recruit bone marrow-derived progenitor cells to arterial thrombi in vivo. J. Exp. Med. 2006, 203, 1221–1233. [Google Scholar] [CrossRef]
- Jorbenadze, R.; Schleicher, E.; Bigalke, B.; Stellos, K.; Gawaz, M. Expression of platelet-bound stromal-cell derived factor-1 (SDF-1) and number of CD34(+) progenitor cells in patients with congestive heart failure. Platelets 2014, 25, 409–415. [Google Scholar] [CrossRef]
- Faraday, N.; Scharpf, R.B.; Dodd-o, J.M.; Martinez, E.A.; Rosenfeld, B.A.; Dorman, T. Leukocytes can enhance platelet-mediated aggregation and thromboxane release via interaction of P-selectin glycoprotein ligand 1 with P-selectin. Anesthesiology 2001, 94, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Maziero, A.M.; Lorenzetti, R.; Donato, J.L.; Lilla, S.; de Nucci, G. Inhibition of human platelet aggregation by eosinophils. Life Sci. 2013, 93, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiang, L.X.; Shao, J.Z.; Pan, R.L.; Wang, Y.X.; Dong, X.J.; Zhang, G.R. Recruitment of endogenous bone marrow mesenchymal stem cells towards injured liver. J. Cell. Mol. Med. 2010, 14, 1494–1508. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhou, X.; Shi, Y.; Li, J.; Zheng, L.; Cui, L.; Zhang, J.; Wang, L.; Han, Z.; Han, Y.; et al. In vivo tracking and comparison of the therapeutic effects of MSCs and HSCs for liver injury. PLoS ONE 2013, 8, e62363. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Takeichi, T.; Asonuma, K.; Tanaka, K.; Kusunoki, M.; Inomata, Y. Intravital imaging of neutrophil recruitment in hepatic ischemia-reperfusion injury in mice. Transplantation 2013, 95, 551–558. [Google Scholar] [CrossRef] [PubMed]
- March, S.; Garcia-Pagán, J.C.; Massaguer, A.; Pizcueta, P.; Panés, J.; Engel, P.; Bosch, J. P-selectin mediates leukocyte rolling in concanavalin-A-induced hepatitis. Liver Int. 2005, 25, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Truong, N.H.; Nguyen, N.H.; Le, T.V.; Vu, N.B.; Huynh, N.; Nguyen, T.V.; Le, H.M.; Phan, N.K.; Pham, P.V. Comparison of the treatment efficiency of bone marrow-derived mesenchymal stem cell transplantation via tail and portal veins in CCl4-induced mouse liver fibrosis. Stem. Cells Int. 2016, 2016, 5720413. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.E.; Elswefy, S.E.; Rashed, L.A.; Younis, N.N.; Shaheen, M.A.; Ghanim, A.M. Bone marrow-derived mesenchymal stem cells effectively regenerate fibrotic liver in bile duct ligation rat model. Exp. Biol. Med. 2016, 241, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.M.; Lian, C.H.; Wu, C.S.; Ji, A.F.; Xiang, J.J.; Wang, X.Y. Effects of bone marrow-derived mesenchymal stem cells transplanted via the portal vein or tail vein on liver injury in rats with liver cirrhosis. Exp. Ther. Med. 2015, 9, 1292–1298. [Google Scholar] [CrossRef] [Green Version]
- Pitchford, S.; Pan, D.; Welch, H.C. Platelets in neutrophil recruitment to sites of inflammation. Curr. Opin. Hematol. 2017, 24, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuchtriegel, G.; Uhl, B.; Puhr-Westerheide, D.; Pörnbacher, M.; Lauber, K.; Krombach, F.; Reichel, C.A. Platelets guide leukocytes to their sites of extravasation. PLoS Biol. 2016, 14, e1002459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, G.S.; Yang, Z.; Carman, C.V.; Karp, J.M.; Lin, C.P. Intravital imaging of mesenchymal stem cell trafficking and association with platelets and neutrophils. Stem. Cells 2015, 33, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, M.; Wang, X.; Lim, B.; Leitner, E.; Klingberg, F.; Ching, V.; Yao, Y.; Huang, D.; Gao, X.M.; Kiriazis, H.; et al. Platelet-targeted delivery of peripheral blood mononuclear cells to the ischemic heart restores cardiac function after ischemia-reperfusion injury. Theranostics 2017, 7, 3192–3206. [Google Scholar] [CrossRef]
- Alvarez-Dolado, M.; Pardal, R.; Garcia-Verdugo, J.M.; Fike, J.R.; Lee, H.O.; Pfeffer, K.; Lois, C.; Morrison, S.J.; Alvarez-Buylla, A. Fusion of bone-marrow-derived cells with Purkinje neurons.; cardiomyocytes and hepatocytes. Nature 2003, 425, 968–973. [Google Scholar] [CrossRef]
- Wagers, A.J.; Weissman, I.L. Plasticity of adult stem cells. Cell 2004, 116, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, T.; Nakanuma, S.; Ahmed, A.K.; Makino, I.; Hayashi, H.; Oyama, K.; Nakagawara, H.; Tajima, H.; Takamura, H.; Ninomiya, I.; et al. Ischemia reperfusion-facilitated sinusoidal endothelial cell injury in liver transplantation and the resulting impact of extravasated platelet aggregation. Eur. Surg. 2016, 48, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Nakanuma, S.; Miyashita, T.; Hayashi, H.; Tajima, H.; Tsukada, T.; Okamoto, K.; Sakai, S.; Makino, I.; Kinoshita, J.; Nakamura, K.; et al. Extravasated platelet aggregation in liver zone 3 may correlate with the progression of sinusoidal obstruction syndrome following living donor liver transplantation: A case report. Exp. Ther. Med. 2015, 9, 1119–1124. [Google Scholar] [CrossRef] [Green Version]
- Ohta, S.; Inujima, Y.; Abe, M.; Uosaki, Y.; Sato, S.; Miki, I. Inhibition of P-selectin specific cell adhesion by a low molecular weight, non-carbohydrate compound, KF38789. Inflamm. Res. 2001, 50, 544–551. [Google Scholar] [CrossRef]
- Duijvestijn, A.M.; van Goor, H.; Klatter, F.; Majoor, G.D.; van Bussel, E.; van Breda, V.P.J. Antibodies defining rat endothelial cells: RECA-1, a pan-endothelial cell-specific monoclonal antibody. Lab. Investig. 1992, 66, 459–466. [Google Scholar]
- Stieger, B.; Hagenbuch, B.; Landmann, L.; Hochli, M.; Schroeder, A.; Meier, P.J. In situ localization of the hepatocytic Na+/Taurocholate cotransporting polypeptide in rat liver. Gastroenterology 1994, 107, 1781–1787. [Google Scholar] [CrossRef]
- Gebhardt, R.; Mecke, D. Heterogeneous distribution of glutamine synthetase among rat liver parenchymal cells in situ and in primary culture. EMBO J. 1983, 2, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Wendel, C.; Hemping-Bovenkerk, A.; Krasnyanska, J.; Mees, S.T.; Kochetkova, M.; Stoeppeler, S.; Haier, J. CXCR4/CXCL12 participate in extravasation of metastasizing breast cancer cells within the liver in a rat model. PLoS ONE 2012, 7, e30046. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehwald, N.; Duhme, C.; Pinchuk, I.; Kirchner, J.; Wieferich, K.; Schmelzle, M.; Jurk, K.; Windmöller, B.A.; Hübner, W.; Homey, B.; et al. Platelets Boost Recruitment of CD133+ Bone Marrow Stem Cells to Endothelium and the Rodent Liver—The Role of P-Selectin/PSGL-1 Interactions. Int. J. Mol. Sci. 2020, 21, 6431. https://doi.org/10.3390/ijms21176431
Lehwald N, Duhme C, Pinchuk I, Kirchner J, Wieferich K, Schmelzle M, Jurk K, Windmöller BA, Hübner W, Homey B, et al. Platelets Boost Recruitment of CD133+ Bone Marrow Stem Cells to Endothelium and the Rodent Liver—The Role of P-Selectin/PSGL-1 Interactions. International Journal of Molecular Sciences. 2020; 21(17):6431. https://doi.org/10.3390/ijms21176431
Chicago/Turabian StyleLehwald, Nadja, Constanze Duhme, Iryna Pinchuk, Julian Kirchner, Kristina Wieferich, Moritz Schmelzle, Kerstin Jurk, Beatrice A. Windmöller, Wolfgang Hübner, Bernhard Homey, and et al. 2020. "Platelets Boost Recruitment of CD133+ Bone Marrow Stem Cells to Endothelium and the Rodent Liver—The Role of P-Selectin/PSGL-1 Interactions" International Journal of Molecular Sciences 21, no. 17: 6431. https://doi.org/10.3390/ijms21176431
APA StyleLehwald, N., Duhme, C., Pinchuk, I., Kirchner, J., Wieferich, K., Schmelzle, M., Jurk, K., Windmöller, B. A., Hübner, W., Homey, B., Bode, J., Kubitz, R., Benhidjeb, T., Krüger, M., Robson, S. C., Knoefel, W. T., Kehrel, B. E., & Schulte am Esch, J. (2020). Platelets Boost Recruitment of CD133+ Bone Marrow Stem Cells to Endothelium and the Rodent Liver—The Role of P-Selectin/PSGL-1 Interactions. International Journal of Molecular Sciences, 21(17), 6431. https://doi.org/10.3390/ijms21176431