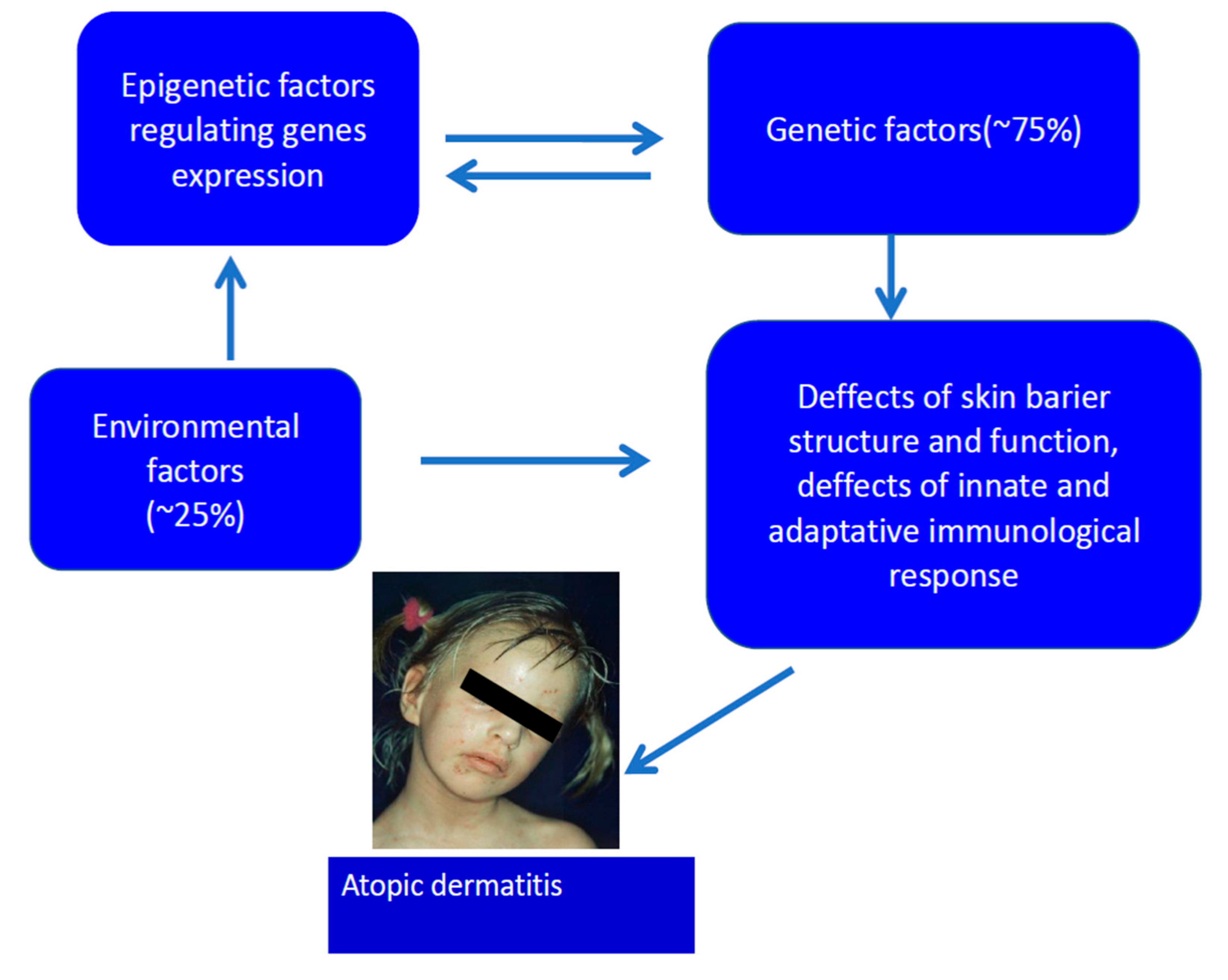
Genetic and Epigenetic Aspects of Atopic Dermatitis

 ,
,  ,
,  , , , , ,
, , , , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Genetics of AD
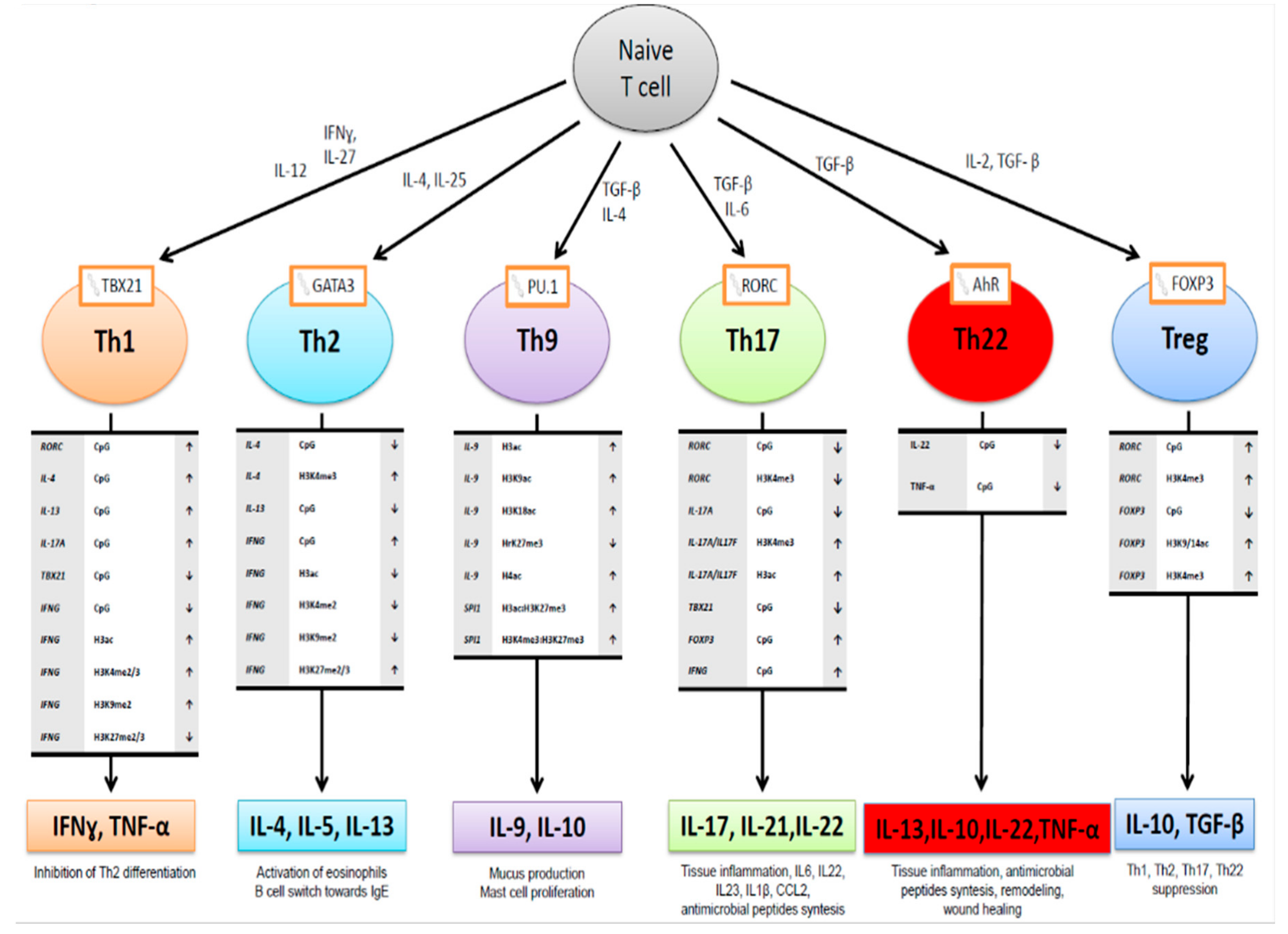
3. Epigenetic Regulation of Gene Expression
- a.
- Post-translational alterations of the histone proteins impacting chromatin architecture, affecting its density and availability for enzyme complexes (Table 1);
- b.
- Methylation, hydroxymethylation, or the demethylation of cytosine in the regulatory gene sequences (promoter or enhancer) altering the transcription of a gene. When the DNA bases cytosine and guanine are next to each other, the silencing of particular genes may occur as a result of adding a methyl group to the DNA molecule, which results in their inactivation and prevention of transcription. On the contrary, the demethylation of the promoter triggers the transcription process and induces the expression of a particular gene (Table 1);
- c.
- Non-coding RNA, including micro-RNAs (miRNAs), small interfering RNAs (siRNAs), long non-coding RNAs (ln-RNAs) and Pivi-interacting RNAs (piRNAs) also comprise an important signaling and regulatory tool. This affects the process of transcription, and may also alter gene expression at the post-transcriptional levels [7,11,12,16,43,44,45,46,47].
3.1. Post-Translational Histone Alterations
3.2. Methylation, Hydroxymethylation or Demethylation of Cytosine
3.3. Micro-RNAs
3.4. Non-Mendelian, “Heretic” Transgeneration Inheritance of Epigenetic Changes
4. Epigenetic Changes in Atopic Dermatitis
5. Epigenetic Regulation of Gene Expression in Pregnancy and Early Childhood
6. Therapies of AD
7. Therapies of AD with Drugs Interfering Epigenetic Processes
8. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Prim. 2018, 4, 1. [Google Scholar] [CrossRef]
- Silverberg, J.I. Atopic Dermatitis in Adults. Med. Clin. 2020, 104, 157–176. [Google Scholar] [CrossRef]
- Schmid, P.; Simon, D.; Simon, H.; Akdis, C.A.; Wüthrich, B. Epidemiology, clinical features, and immunology of the “intrinsic”(non-IgE-mediated) type of atopic dermatitis (constitutional dermatitis). Allergy 2001, 56, 841–849. [Google Scholar]
- Flohr, C.; Johansson, S.G.O.; Wahlgren, C.-F.; Williams, H. How atopic is atopic dermatitis? J. Allergy Clin. Immunol. 2004, 114, 150–158. [Google Scholar] [CrossRef]
- Gutowska-Owsiak, D.; Salimi, M.; Selvakumar, T.A.; Wang, X.; Taylor, S.; Ogg, G.S. Histamine exerts multiple effects on expression of genes associated with epidermal barrier function. J. Investig. Allergol. Clin. Immunol. 2014, 24, 231–239. [Google Scholar]
- Gutowska-Owsiak, D.; Schaupp, A.L.; Salimi, M.; Selvakumar, T.A.; McPherson, T.; Taylor, S.; Ogg, G.S. IL-17 downregulates filaggrin and affects keratinocyte expression of genes associated with cellular adhesion. Exp. Dermatol. 2012, 21, 104–110. [Google Scholar] [CrossRef]
- Martin, M.J.; Estravís, M.; García-Sánchez, A.; Dávila, I.; Isidoro-García, M.; Sanz, C. Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review. Genes 2020, 11, 442. [Google Scholar] [CrossRef] [Green Version]
- Løset, M.; Brown, S.J.; Saunes, M.; Hveem, K. Genetics of atopic dermatitis: From DNA sequence to clinical relevance. Dermatology 2019, 235, 355–364. [Google Scholar] [CrossRef]
- Gutowska-Owsiak, D.; Schaupp, A.L.; Salimi, M.; Taylor, S.; Ogg, G.S. Interleukin-22 downregulates filaggrin expression and affects expression of profilaggrin processing enzymes. Br. J. Dermatol. 2011, 165, 492–498. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Mildner, M.; Mlitz, V.; Gruber, F.; Eckhart, L.; Werfel, T.; Gutzmer, R.; Elias, P.M.; Tschachler, E. Histamine suppresses epidermal keratinocyte differentiation and impairs skin barrier function in a human skin model. Allergy 2013, 68, 37–47. [Google Scholar] [CrossRef]
- Liang, Y.; Chang, C.; Lu, Q. The Genetics and Epigenetics of Atopic Dermatitis-Filaggrin and Other Polymorphisms. Clin. Rev. Allergy Immunol. 2016, 51, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Bin, L.; Leung, D.Y.M. Genetic and epigenetic studies of atopic dermatitis. Allergy Asthma Clin. Immunol. Off. J. Can. Soc. Allergy Clin. Immunol. 2016, 12, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stemmler, S.; Hoffjan, S. Trying to understand the genetics of atopic dermatitis. Mol. Cell. Probes 2016, 30, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Bonamonte, D.; Filoni, A.; Vestita, M.; Romita, P.; Foti, C.; Angelini, G. The role of the environmental risk factors in the pathogenesis and clinical outcome of atopic dermatitis. Biomed Res. Int. 2019, 2019, 2450605. [Google Scholar] [CrossRef] [PubMed]
- Hoffjan, S.; Stemmler, S. Unravelling the complex genetic background of atopic dermatitis: From genetic association results towards novel therapeutic strategies. Arch. Dermatol. Res. 2015, 307, 659–670. [Google Scholar] [CrossRef]
- Mu, Z.; Zhang, J. The Role of Genetics, the Environment, and Epigenetics in Atopic Dermatitis. Adv. Exp. Med. Biol. 2020, 1253, 107–140. [Google Scholar]
- Totté, J.E.E.; van der Feltz, W.T.; Hennekam, M.; van Belkum, A.; van Zuuren, E.J.; Pasmans, S.G.M.A. Prevalence and odds of Staphylococcus aureus carriage in atopic dermatitis: A systematic review and meta-analysis. Br. J. Dermatol. 2016, 175, 687–695. [Google Scholar] [CrossRef]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet (London England) 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Rebane, A.; Akdis, C.A. MicroRNAs: Essential players in the regulation of inflammation. J. Allergy Clin. Immunol. 2013, 132, 15–26. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Ruckemann-Dziurdzińska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Postep. Dermatologii I Alergol. 2014, 31, 84–91. [Google Scholar] [CrossRef]
- Samotij, D.; Nedoszytko, B.; Bartosińska, J.; Batycka-Baran, A.; Czajkowski, R.; Dobrucki, I.T.; Dobrucki, L.W.; Górecka-Sokołowska, M.; Janaszak-Jasienicka, A.; Krasowska, D.; et al. Pathogenesis of psoriasis in the “omic” era. Part I. Epidemiology, clinical manifestation, immunological and neuroendocrine disturbances. Postep. Dermatologii I Alergol. 2020, 37, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Nedoszytko, B.; Szczerkowska-Dobosz, A.; Stawczyk-Maciej, M.; Owczarczyk-Saczone, A. Pathogenesis of psoriasis in the “omics” era. Part II - Genetic, genomic and epigenetic changes in psoriasis. Adv. Dermatol. Allergol. 2020, 37, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gimalova, G.F.; Karunas, A.S.; Fedorova, Y.Y.; Gumennaya, E.R.; Levashova, S.V.; Khismatullina, Z.R.; Prans, E.; Kõks, S.; Etkina, E.I.; Khusnutdinova, E.K. Association of polymorphisms in the toll-like receptor genes with atopic dermatitis in the Republic of Bashkortostan. Mol. Biol. 2014, 48, 227–237. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.-C.; Feng, C.; Yan, M. Analysis of the Association of Polymorphisms rs5743708 in TLR2 and rs4986790 in TLR4 with Atopic Dermatitis Risk. Immunol. Invest. 2019, 48, 169–180. [Google Scholar] [CrossRef]
- Trzeciak, M.; Wesserling, M.; Bandurski, T.; Glen, J.; Nowicki, R.; Pawelczyk, T. Association of a Single Nucleotide Polymorphism in a Late Cornified Envelope-like Proline-rich 1 Gene (LELP1) with Atopic Dermatitis. Acta Derm. Venereol. 2016, 96, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Sokołowska-Wojdyło, M.; Gleń, J.; Zabłotna, M.; Rębała, K.; Trzeciak, M.; Sikorska, M.; Ruckemann-Dziurdzińska, K.; Nedoszytko, B.; Florek, A.; Nowicki, R. The frequencies of haplotypes defined by three polymorphisms of the IL-31 gene: −1066, −2057, and IVS2+12 in Polish patients with atopic dermatitis. Int. J. Dermatol. 2015, 54, 62–67. [Google Scholar] [CrossRef]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [Green Version]
- Zablotna, M.; Sobjanek, M.; Glen, J.; Niedoszytko, M.; Wilkowska, A.; Roszkiewicz, J.; Nedoszytko, B. Association between the -1154 G/A promoter polymorphism of the vascular endothelial growth factor gene and atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 91–92. [Google Scholar] [CrossRef]
- Trzeciak, M.; Gleń, J.; Roszkiewicz, J.; Nedoszytko, B. Association of single nucleotide polymorphism of interleukin-18 with atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 78–79. [Google Scholar] [CrossRef]
- Trzeciak, M.; Gleń, J.; Bandurski, T.; Sokołowska-Wojdyło, M.; Wilkowska, A.; Roszkiewicz, J. Relationship between serum levels of interleukin-18, IgE and disease severity in patients with atopic dermatitis. Clin. Exp. Dermatol. 2011, 36, 728–732. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Niedoszytko, M.; Lange, M.; Van Doormaal, J.; Gleń, J.; Zabłotna, M.; Renke, J.; Vales, A.; Buljubasic, F.; Jassem, E.; et al. Interleukin-13 promoter gene polymorphism -1112C/T is associated with the systemic form of mastocytosis. Allergy 2009, 64, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Wilkowska, A.; Gleń, J.; Zabłotna, M.; Trzeciak, M.; Ryduchowska, M.; Sobjanek, M.; Nedoszytko, B.; Nowicki, R.; Sokołowska-Wojdyło, M. The association of GM-CSF -677A/C promoter gene polymorphism with the occurrence and severity of atopic dermatitis in a Polish population. Int. J. Dermatol. 2014, 53, e172–e174. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Illig, T.; Baurecht, H.; Irvine, A.D.; Rodriguez, E.; Diaz-Lacava, A.; Klopp, N.; Wagenpfeil, S.; Zhao, Y.; Liao, H. Loss-of-function variations within the filaggrin gene predispose for atopic dermatitis with allergic sensitizations. J. Allergy Clin. Immunol. 2006, 118, 214–219. [Google Scholar] [CrossRef]
- McLean, W.H.I. Filaggrin failure-from ichthyosis vulgaris to atopic eczema and beyond. Br. J. Dermatol. 2016, 175 (Suppl. 4–7). [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; Kroboth, K.; Sandilands, A.; Campbell, L.E.; Pohler, E.; Kezic, S.; Cordell, H.J.; McLean, W.H.I.; Irvine, A.D. Intragenic copy number variation within filaggrin contributes to the risk of atopic dermatitis with a dose-dependent effect. J. Invest. Dermatol. 2012, 132, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, H.; Nagao, K.; Kubo, A.; Hata, T.; Shimizu, A.; Mizuno, H.; Yamada, T.; Amagai, M. Altered stratum corneum barrier and enhanced percutaneous immune responses in filaggrin-null mice. J. Allergy Clin. Immunol. 2012, 129, 1538–1546. [Google Scholar] [CrossRef] [Green Version]
- Marwah, I.; Wang, X.; Chan, H.; Ogg, G.S.; Gutowska-Owsiak, D. Filaggrin-insufficiency in keratinocytes influences responsiveness of allergen-specific T cells to cognate antigen and compounds barrier function deficiency. Clin. Immunol. 2014, 153, 153–155. [Google Scholar] [CrossRef]
- Jarrett, R.; Salio, M.; Lloyd-Lavery, A.; Subramaniam, S.; Bourgeois, E.; Archer, C.; Cheung, K.L.; Hardman, C.; Chandler, D.; Salimi, M.; et al. Filaggrin inhibits generation of CD1a neolipid antigens by house dust mite-derived phospholipase. Sci. Transl. Med. 2016, 8, 325. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Cho, K.-A.; Kim, J.-Y.; Kim, J.-Y.; Baek, J.-H.; Woo, S.-Y.; Kim, J.-W. Filaggrin knockdown and Toll-like receptor 3 (TLR3) stimulation enhanced the production of thymic stromal lymphopoietin (TSLP) from epidermal layers. Exp. Dermatol. 2011, 20, 149–151. [Google Scholar] [CrossRef]
- Leitch, C.S.; Natafji, E.; Yu, C.; Abdul-Ghaffar, S.; Madarasingha, N.; Venables, Z.C.; Chu, R.; Fitch, P.M.; Muinonen-Martin, A.J.; Campbell, L.E.; et al. Filaggrin-null mutations are associated with increased maturation markers on Langerhans cells. J. Allergy Clin. Immunol. 2016, 138, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziyab, A.H.; Karmaus, W.; Holloway, J.W.; Zhang, H.; Ewart, S.; Arshad, S.H. DNA methylation of the filaggrin gene adds to the risk of eczema associated with loss-of-function variants. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e420–e423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harb, H.; Renz, H. Update on epigenetics in allergic disease. J. Allergy Clin. Immunol. 2015, 135, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, B.E.; Meissner, A.; Lander, E.S. The mammalian epigenome. Cell 2007, 128, 669–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botchkarev, V.A.; Gdula, M.R.; Mardaryev, A.N.; Sharov, A.A.; Fessing, M.Y. Epigenetic regulation of gene expression in keratinocytes. J. Invest. Dermatol. 2012, 132, 2505–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makeyev, E.V.; Maniatis, T. Multilevel regulation of gene expression by microRNAs. Science 2008, 319, 1789–1790. [Google Scholar] [CrossRef] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Jones, B.; Chen, J. Inhibition of IFN-gamma transcription by site-specific methylation during T helper cell development. EMBO J. 2006, 25, 2443–2452. [Google Scholar] [CrossRef]
- Lee, D.U.; Agarwal, S.; Rao, A. Th2 lineage commitment and efficient IL-4 production involves extended demethylation of the IL-4 gene. Immunity 2002, 16, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and allergy: From basic mechanisms to clinical applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Renke, J.; Lange, M.; Trzonkowski, P.; Sobjanek, M.; Szczerkowska-Dobosz, A.; Niedoszytko, M.; Górska, A.; Romantowski, J.; et al. The role of regulatory T cells and genes involved in their differentiation in pathogenesis of selected inflammatory and neoplastic skin diseases. Part III: Polymorphisms of genes involved in Tregs’ activation and function. Postep. Dermatologii I Alergol. 2017, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrmann, J.; Tatura, R.; Zeschnigk, M.; Probst-Kepper, M.; Geffers, R.; Steinmann, J.; Buer, J. Impact of 5-aza-2′-deoxycytidine and epigallocatechin-3-gallate for induction of human regulatory T cells. Immunology 2014, 142, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs and immunity: Novel players in the regulation of normal immune function and inflammation. Semin. Cancer Biol. 2008, 18, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.R.; Brede, G.; Johansen, J.; Johnsen, R.; Storrø, O.; Sætrom, P.; Øien, T. Human breast milk miRNA, maternal probiotic supplementation and atopic dermatitis in offspring. PLoS ONE 2015, 10, e0143496. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs: Novel regulators in skin inflammation. Clin. Exp. Dermatol. 2008, 33, 312–315. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. Diagnostic, functional, and therapeutic roles of microRNA in allergic diseases. J. Allergy Clin. Immunol. 2013, 132, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B.C.; John, S.M.; Schmitz, G. Milk: An exosomal microRNA transmitter promoting thymic regulatory T cell maturation preventing the development of atopy? J. Transl. Med. 2014, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Ma, Y.-L.; Liang, W.; Li, H.-H.; Ma, Z.-J.; Yu, X.; Liao, Y.-H. MicroRNA-155 modulates Treg and Th17 cells differentiation and Th17 cell function by targeting SOCS1. PLoS ONE 2012, 7, e46082. [Google Scholar] [CrossRef] [Green Version]
- Batagov, A.O.; Kurochkin, I. V Exosomes secreted by human cells transport largely mRNA fragments that are enriched in the 3′-untranslated regions. Biol. Direct 2013, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- van Heyningen, V.; Yeyati, P.L. Mechanisms of non-Mendelian inheritance in genetic disease. Hum. Mol. Genet. 2004, 13, R225–R233. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, T.M.; Rezwan, F.I.; Jiang, Y.; Karmaus, W.; Svanes, C.; Holloway, J.W. Transgenerational and intergenerational epigenetic inheritance in allergic diseases. J. Allergy Clin. Immunol. 2018, 142, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Legoff, L.; D’Cruz, S.C.; Tevosian, S.; Primig, M.; Smagulova, F. Transgenerational inheritance of environmentally induced epigenetic alterations during mammalian development. Cells 2019, 8, 1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.M.; Liao, W. Genomic imprinting in psoriasis and atopic dermatitis: A review. J. Dermatol. Sci. 2015, 80, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, H.E.; Moffatt, M.F.; Faux, J.A.; Walley, A.J.; Coleman, R.; Trembath, R.C.; Cookson, W.O.; Harper, J.I. Association of atopic dermatitis to the beta subunit of the high affinity immunoglobulin E receptor. Br. J. Dermatol. 1998, 138, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.A.; Wahn, U.; Kehrt, R.; Tarani, L.; Businco, L.; Gustafsson, D.; Andersson, F.; Oranje, A.P.; Wolkertstorfer, A.; v Berg, A.; et al. A major susceptibility locus for atopic dermatitis maps to chromosome 3q21. Nat. Genet. 2000, 26, 470–473. [Google Scholar] [CrossRef]
- Kaiser, J. The epigenetics heretic. Science 2014, 343, 361–363. [Google Scholar] [CrossRef]
- Rodríguez, E.; Baurecht, H.; Wahn, A.F.; Kretschmer, A.; Hotze, M.; Zeilinger, S.; Klopp, N.; Illig, T.; Schramm, K.; Prokisch, H.; et al. An integrated epigenetic and transcriptomic analysis reveals distinct tissue-specific patterns of DNA methylation associated with atopic dermatitis. J. Investig. Dermatol. 2014, 134, 1873–1883. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Zhou, B.; Zhao, M.; Tang, J.; Lu, Q. Promoter demethylation contributes to TSLP overexpression in skin lesions of patients with atopic dermatitis. Clin. Exp. Dermatol. 2014, 39, 48–53. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, P.; Zhao, M.; Liang, G.; Yin, H.; Zhang, G.; Wen, H.; Lu, Q. Demethylation of the FCER1G promoter leads to FcεRI overexpression on monocytes of patients with atopic dermatitis. Allergy 2012, 67, 424–430. [Google Scholar] [CrossRef]
- Sonkoly, E.; Janson, P.; Majuri, M.-L.; Savinko, T.; Fyhrquist, N.; Eidsmo, L.; Xu, N.; Meisgen, F.; Wei, T.; Bradley, M.; et al. MiR-155 is overexpressed in patients with atopic dermatitis and modulates T-cell proliferative responses by targeting cytotoxic T lymphocyte-associated antigen 4. J. Allergy Clin. Immunol. 2010, 126, 520–581. [Google Scholar] [CrossRef]
- Lu, T.X.; Hartner, J.; Lim, E.-J.; Fabry, V.; Mingler, M.K.; Cole, E.T.; Orkin, S.H.; Aronow, B.J.; Rothenberg, M.E. MicroRNA-21 limits in vivo immune response-mediated activation of the IL-12/IFN-gamma pathway, Th1 polarization, and the severity of delayed-type hypersensitivity. J. Immunol. 2011, 187, 3362–3373. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a alleviates chronic skin inflammation in atopic dermatitis through suppression of innate immune responses in keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.L.; Zhang, Z.; Johansson, E.; Ray, S.; Jagpal, A.; Ruff, B.P.; Kothari, A.; He, H.; Martin, L.J.; Ji, H.; et al. Disease-associated KIF3A variants alter gene methylation and expression impacting skin barrier and atopic dermatitis risk. Nat. Commun. 2020, 11, 4092. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.; Myers, J.M.B.; Martin, L.J.; He, H.; Pilipenko, V.; Mersha, T.; Weirauch, M.; Salomonis, N.; Ryan, P.; LeMasters, G.K. KIF3A genetic variation is associated with pediatric asthma in the presence of eczema independent of allergic rhinitis. J. Allergy Clin. Immunol. 2017, 140, 595–598. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, M.; Li, L.; Zhang, L.; Chan, M.T.V.; Wu, W.K.K. MicroRNAs in atopic dermatitis: A systematic review. J. Cell. Mol. Med. 2020, 24, 5966–5972. [Google Scholar] [CrossRef]
- Li, H.M.; Xiao, Y.J.; Min, Z.S.; Tan, C. Identification and interaction analysis of key genes and micro RNA s in atopic dermatitis by bioinformatics analysis. Clin. Exp. Dermatol. 2019, 44, 257–264. [Google Scholar] [CrossRef]
- Yan, F.; Meng, W.; Ye, S.; Zhang, X.; Mo, X.; Liu, J.; Chen, D.; Lin, Y. MicroRNA-146a as a potential regulator involved in the pathogenesis of atopic dermatitis. Mol. Med. Rep. 2019, 20, 4645–4653. [Google Scholar] [CrossRef]
- Sonkoly, E.; Wei, T.; Janson, P.C.J.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A. MicroRNAs: Novel regulators involved in the pathogenesis of psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Xue, H.; Wang, F.; Shu, C.; Zhang, J. MicroRNA-155 may be involved in the pathogenesis of atopic dermatitis by modulating the differentiation and function of T helper type 17 (Th17) cells. Clin. Exp. Immunol. 2015, 181, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Yoon, W.S.; Lee, S.S.; Chae, Y.S.; Park, Y.K. Therapeutic effects of recombinant Salmonella typhimurium harboring CCL22 miRNA on atopic dermatitis-like skin in mice. Exp. Mol. Med. 2011, 43, 63–70. [Google Scholar] [CrossRef]
- Dissanayake, E.; Inoue, Y.; Ochiai, S.; Eguchi, A.; Nakano, T.; Yamaide, F.; Hasegawa, S.; Kojima, H.; Suzuki, H.; Mori, C. Hsa-mir-144-3p expression is increased in umbilical cord serum of infants with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 447–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zhang, L.; Zhang, J.; Dou, X.; Shao, Y.; Jia, X.; Zhang, W.; Yu, B. MiR-151a is involved in the pathogenesis of atopic dermatitis by regulating interleukin-12 receptor β2. Exp. Dermatol. 2018, 27, 427–432. [Google Scholar] [CrossRef] [PubMed]
- White, G.P.; Watt, P.M.; Holt, B.J.; Holt, P.G. Differential patterns of methylation of the IFN-gamma promoter at CpG and non-CpG sites underlie differences in IFN-gamma gene expression between human neonatal and adult CD45RO-T cells. J. Immunol. 2002, 168, 2820–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminuma, O.; Kitamura, F.; Miyatake, S.; Yamaoka, K.; Miyoshi, H.; Inokuma, S.; Tatsumi, H.; Nemoto, S.; Kitamura, N.; Mori, A.; et al. T-box 21 transcription factor is responsible for distorted T(H)2 differentiation in human peripheral CD4+ T cells. J. Allergy Clin. Immunol. 2009, 123, 813–823.e3. [Google Scholar] [CrossRef] [PubMed]
- Fujimaki, W.; Takahashi, N.; Ohnuma, K.; Nagatsu, M.; Kurosawa, H.; Yoshida, S.; Dang, N.H.; Uchiyama, T.; Morimoto, C. Comparative study of regulatory T cell function of human CD25CD4 T cells from thymocytes, cord blood, and adult peripheral blood. Clin. Dev. Immunol. 2008, 2008, 305859. [Google Scholar] [CrossRef]
- Schaub, B.; Liu, J.; Schleich, I.; Höppler, S.; Sattler, C.; von Mutius, E. Impairment of T helper and T regulatory cell responses at birth. Allergy 2008, 63, 1438–1447. [Google Scholar] [CrossRef]
- Cao, S.; Feehley, T.J.; Nagler, C.R. The role of commensal bacteria in the regulation of sensitization to food allergens. FEBS Lett. 2014, 588, 4258–4266. [Google Scholar] [CrossRef] [Green Version]
- Amberbir, A.; Medhin, G.; Erku, W.; Alem, A.; Simms, R.; Robinson, K.; Fogarty, A.; Britton, J.; Venn, A.; Davey, G. Effects of Helicobacter pylori, geohelminth infection and selected commensal bacteria on the risk of allergic disease and sensitization in 3-year-old Ethiopian children. Clin. Exp. Allergy 2011, 41, 1422–1430. [Google Scholar] [CrossRef]
- Sharma, M.; Li, Y.; Stoll, M.L.; Tollefsbol, T.O. The Epigenetic Connection Between the Gut Microbiome in Obesity and Diabetes. Front. Genet. 2019, 10, 1329. [Google Scholar] [CrossRef] [Green Version]
- Schaub, B.; Liu, J.; Höppler, S.; Schleich, I.; Huehn, J.; Olek, S.; Wieczorek, G.; Illi, S.; von Mutius, E. Maternal farm exposure modulates neonatal immune mechanisms through regulatory T cells. J. Allergy Clin. Immunol. 2009, 123, 774–782. [Google Scholar] [CrossRef]
- Lluis, A.; Depner, M.; Gaugler, B.; Saas, P.; Casaca, V.I.; Raedler, D.; Michel, S.; Tost, J.; Liu, J.; Genuneit, J.; et al. Increased regulatory T-cell numbers are associated with farm milk exposure and lower atopic sensitization and asthma in childhood. J. Allergy Clin. Immunol. 2014, 133, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Michel, S.; Busato, F.; Genuneit, J.; Pekkanen, J.; Dalphin, J.-C.; Riedler, J.; Mazaleyrat, N.; Weber, J.; Karvonen, A.M.; Hirvonen, M.-R.; et al. Farm exposure and time trends in early childhood may influence DNA methylation in genes related to asthma and allergy. Allergy 2013, 68, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Schieck, M.; Sharma, V.; Michel, S.; Toncheva, A.A.; Worth, L.; Potaczek, D.P.; Genuneit, J.; Kretschmer, A.; Depner, M.; Dalphin, J.-C.; et al. A polymorphism in the TH 2 locus control region is associated with changes in DNA methylation and gene expression. Allergy 2014, 69, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Brand, S.; Teich, R.; Dicke, T.; Harb, H.; Yildirim, A.Ö.; Tost, J.; Schneider-Stock, R.; Waterland, R.A.; Bauer, U.-M.; von Mutius, E.; et al. Epigenetic regulation in murine offspring as a novel mechanism for transmaternal asthma protection induced by microbes. J. Allergy Clin. Immunol. 2011, 128, 617–618. [Google Scholar] [CrossRef]
- Breton, C.V.; Byun, H.-M.; Wenten, M.; Pan, F.; Yang, A.; Gilliland, F.D. Prenatal tobacco smoke exposure affects global and gene-specific DNA methylation. Am. J. Respir. Crit. Care Med. 2009, 180, 462–467. [Google Scholar] [CrossRef]
- Wang, I.-J.; Chen, S.-L.; Lu, T.-P.; Chuang, E.Y.; Chen, P.-C. Prenatal smoke exposure, DNA methylation, and childhood atopic dermatitis. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2013, 43, 535–543. [Google Scholar] [CrossRef]
- Tang, W.; Levin, L.; Talaska, G.; Cheung, Y.Y.; Herbstman, J.; Tang, D.; Miller, R.L.; Perera, F.; Ho, S.-M. Maternal exposure to polycyclic aromatic hydrocarbons and 5′-CpG methylation of interferon-γ in cord white blood cells. Environ. Health Perspect. 2012, 120, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- Hinz, D.; Bauer, M.; Röder, S.; Olek, S.; Huehn, J.; Sack, U.; Borte, M.; Simon, J.C.; Lehmann, I.; Herberth, G. Cord blood Tregs with stable FOXP3 expression are influenced by prenatal environment and associated with atopic dermatitis at the age of one year. Allergy 2012, 67, 380–389. [Google Scholar] [CrossRef]
- Herberth, G.; Bauer, M.; Gasch, M.; Hinz, D.; Röder, S.; Olek, S.; Kohajda, T.; Rolle-Kampczyk, U.; von Bergen, M.; Sack, U.; et al. Maternal and cord blood miR-223 expression associates with prenatal tobacco smoke exposure and low regulatory T-cell numbers. J. Allergy Clin. Immunol. 2014, 133, 543–550. [Google Scholar] [CrossRef]
- Wollenberg, A.; Szepietowski, J.; Taieb, A.; Ring, J. Corrigendum: Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults and children: Part I. J. Eur. Acad. Dermatology Venereol. 2019, 33, 1436. [Google Scholar] [CrossRef] [Green Version]
- Wollenberg, A.; Oranje, A.; Deleuran, M.; Simon, D.; Szalai, Z.; Kunz, B.; Svensson, A.; Barbarot, S.; von Kobyletzki, L.; Taieb, A.; et al. ETFAD/EADV Eczema task force 2015 position paper on diagnosis and treatment of atopic dermatitis in adult and paediatric patients. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 729–747. [Google Scholar] [CrossRef] [Green Version]
- Reitamo, S.; Wollenberg, A.; Schöpf, E.; Perrot, J.L.; Marks, R.; Ruzicka, T.; Christophers, E.; Kapp, A.; Lahfa, M.; Rubins, A.; et al. Safety and efficacy of 1 year of tacrolimus ointment monotherapy in adults with atopic dermatitis. The European Tacrolimus Ointment Study Group. Arch. Dermatol. 2000, 136, 999–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devillers, A.C.A.; Oranje, A.P. Wet-wrap treatment in children with atopic dermatitis: A practical guideline. Pediatr. Dermatol. 2012, 29, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; Gieler, U.; Girolomoni, G.; Lau, S.; Muraro, A. Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults and children: Part II. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 850–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altucci, L.; Clarke, N.; Nebbioso, A.; Scognamiglio, A.; Gronemeyer, H. Acute myeloid leukemia: Therapeutic impact of epigenetic drugs. Int. J. Biochem. Cell Biol. 2005, 37, 1752–1762. [Google Scholar] [CrossRef]
- Blum, W.; Marcucci, G. Targeting epigenetic changes in acute myeloid leukemia. Clin. Adv. Hematol. Oncol. 2005, 3, 855–865. [Google Scholar]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone Deacetylase Inhibitors as Anticancer Drugs. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Steelant, B.; Wawrzyniak, P.; Martens, K.; Jonckheere, A.-C.; Pugin, B.; Schrijvers, R.; Bullens, D.M.; Vanoirbeek, J.A.; Krawczyk, K.; Dreher, A.; et al. Blocking histone deacetylase activity as a novel target for epithelial barrier defects in patients with allergic rhinitis. J. Allergy Clin. Immunol. 2019, 144, 1242–1253. [Google Scholar] [CrossRef] [Green Version]
- Oh, K.-S.; Patel, H.; Gottschalk, R.A.; Lee, W.S.; Baek, S.; Fraser, I.D.C.; Hager, G.L.; Sung, M.-H. Anti-Inflammatory Chromatinscape Suggests Alternative Mechanisms of Glucocorticoid Receptor Action. Immunity 2017, 47, 298–309. [Google Scholar] [CrossRef]
- Wu, C.-J.; Yang, C.-Y.; Chen, Y.-H.; Chen, C.-M.; Chen, L.-C.; Kuo, M.-L. The DNA methylation inhibitor 5-azacytidine increases regulatory T cells and alleviates airway inflammation in ovalbumin-sensitized mice. Int. Arch. Allergy Immunol. 2013, 160, 356–364. [Google Scholar] [CrossRef]
- Liew, W.C.; Sundaram, G.M.; Quah, S.; Lum, G.G.; Tan, J.S.L.; Ramalingam, R.; Common, J.E.A.; Tang, M.B.Y.; Lane, E.B.; Thng, S.T.G. Belinostat resolves skin barrier defects in atopic dermatitis by targeting the dysregulated miR-335: SOX6 axis. J. Allergy Clin. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Pathological Process in AD | Example of Genes Involved |
|---|---|
| Epidermal barrier genes | Filaggrin, filaggrin 2, hornerin Corneodesmosomal genes (desmoglein, desmocollin) and tight junction genes (claudins, ocludins) Epidermal protease genes (kallikreins, cathepsins, caspase 14), and their inhibitors (SPINK5, Cystatin A) OVOL1 (ovo like transcriptional repressor)—transcription factor that regulates FLG expression |
| Genes of the innate immune mechanisms | TLR1, TLR2, TLR4, TLR6, TLR9, TLR10, CD14, NOD1 and defensins (DEFB1) |
| Genes of the adaptive immune mechanism | Genes of receptor subunits for IgE (FcεRI α i FCεRI-¥) Genes of Th2 response: IL-4, IL-5, IL-13, IL2RA, IL-13RA IL-5RA, TSLPR, IL-4R, IL-18, IL-31 Other genes of Th bias IL17A, TNFα, IL-22 Treg genes: STAT-6, FOXP3, LRRC32 |
| Genes encoding alarmins produced by keratinocytes | IL-25, TSLP, IL-33 |
| Genes regulating DNA methylation | KIF3A |
| Genes regulating vitamin D pathways | CYP27A1, CYP2R1, VDR |
| Chromatin Modification | Activation of Gene Transcription | Deactivation of Gene Transcription | ||
|---|---|---|---|---|
| Promoter methylation | ||||
| Methylation of cytosine in gene promoter | no | yes | ||
| Demethylation of cytosine in the promoter | yes | no | ||
| Histone modification | ||||
| Acetyla-tion of histone amino acids residues | H2A | K5* | yes | no |
| H2B | K5,K12,K15,K20 | yes | no | |
| H3 | K4,K14,K18,K23,K27 | yes | no | |
| H4 | K8, K16 | yes | no | |
| Methy-lation of histone amino acid residues | H3 | K4, K79, R17 | yes | no |
| H3 | K9, K27 | no | yes | |
| H4 | R3 | yes | no | |
| H4 | K20 | no | yes | |
| Ubiquiti-nation of histones | H2A | K119 | no | yes |
| H2B | K120 | yes | no | |
| miRNA Expression Change in AD Lesions or Serum | Associated Effect |
|---|---|
| ↓Let-7 a-d | ↑IL-13↑CCR7 |
| ↓miR-375 | ↑TSLP (Thymic stromal lymphopoietin) |
| ↓hsa-miR-26a-5a | ↑HAS3 (hyaluronian 3 synthase) |
| ↑miR-21 | ↓IL12 |
| ↑miR-29b | Promotion of INF-γ-induced keratinocyte apoptosis |
| ↑miR-146a | ↓STAT1 and decrease in Treg activation ↓NFκB–pro-inflammatory transcription factor |
| ↑miR-155 | ↓CTLA-4 and decrease in Treg proliferation ↑Promotion of Th17 differentiation ↓Inhibition of tight junction formation |
| ↑miR-223 in umbilical cord blood | Decrease in Treg activation |
| ↑MiR-151a in serum | Inhibition of IL-12 signaling |
| Other miRNA expression changes in atopic skin: ↑ miR-17-5p, ↑ miR142-3p/5p, ↓ miR-122a, ↓miR-326, ↓miR-133b, ↓miR-125b, ↓miR375, ↓ miR193c, ↓miR365 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedoszytko, B.; Reszka, E.; Gutowska-Owsiak, D.; Trzeciak, M.; Lange, M.; Jarczak, J.; Niedoszytko, M.; Jablonska, E.; Romantowski, J.; Strapagiel, D.; et al. Genetic and Epigenetic Aspects of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 6484. https://doi.org/10.3390/ijms21186484
Nedoszytko B, Reszka E, Gutowska-Owsiak D, Trzeciak M, Lange M, Jarczak J, Niedoszytko M, Jablonska E, Romantowski J, Strapagiel D, et al. Genetic and Epigenetic Aspects of Atopic Dermatitis. International Journal of Molecular Sciences. 2020; 21(18):6484. https://doi.org/10.3390/ijms21186484
Chicago/Turabian StyleNedoszytko, Bogusław, Edyta Reszka, Danuta Gutowska-Owsiak, Magdalena Trzeciak, Magdalena Lange, Justyna Jarczak, Marek Niedoszytko, Ewa Jablonska, Jan Romantowski, Dominik Strapagiel, and et al. 2020. "Genetic and Epigenetic Aspects of Atopic Dermatitis" International Journal of Molecular Sciences 21, no. 18: 6484. https://doi.org/10.3390/ijms21186484
APA StyleNedoszytko, B., Reszka, E., Gutowska-Owsiak, D., Trzeciak, M., Lange, M., Jarczak, J., Niedoszytko, M., Jablonska, E., Romantowski, J., Strapagiel, D., Skokowski, J., Siekierzycka, A., Nowicki, R. J., Dobrucki, I. T., Zaryczańska, A., & Kalinowski, L. (2020). Genetic and Epigenetic Aspects of Atopic Dermatitis. International Journal of Molecular Sciences, 21(18), 6484. https://doi.org/10.3390/ijms21186484