Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem (iPS) Cells Suppress or Activate T Cells via Costimulatory Signals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
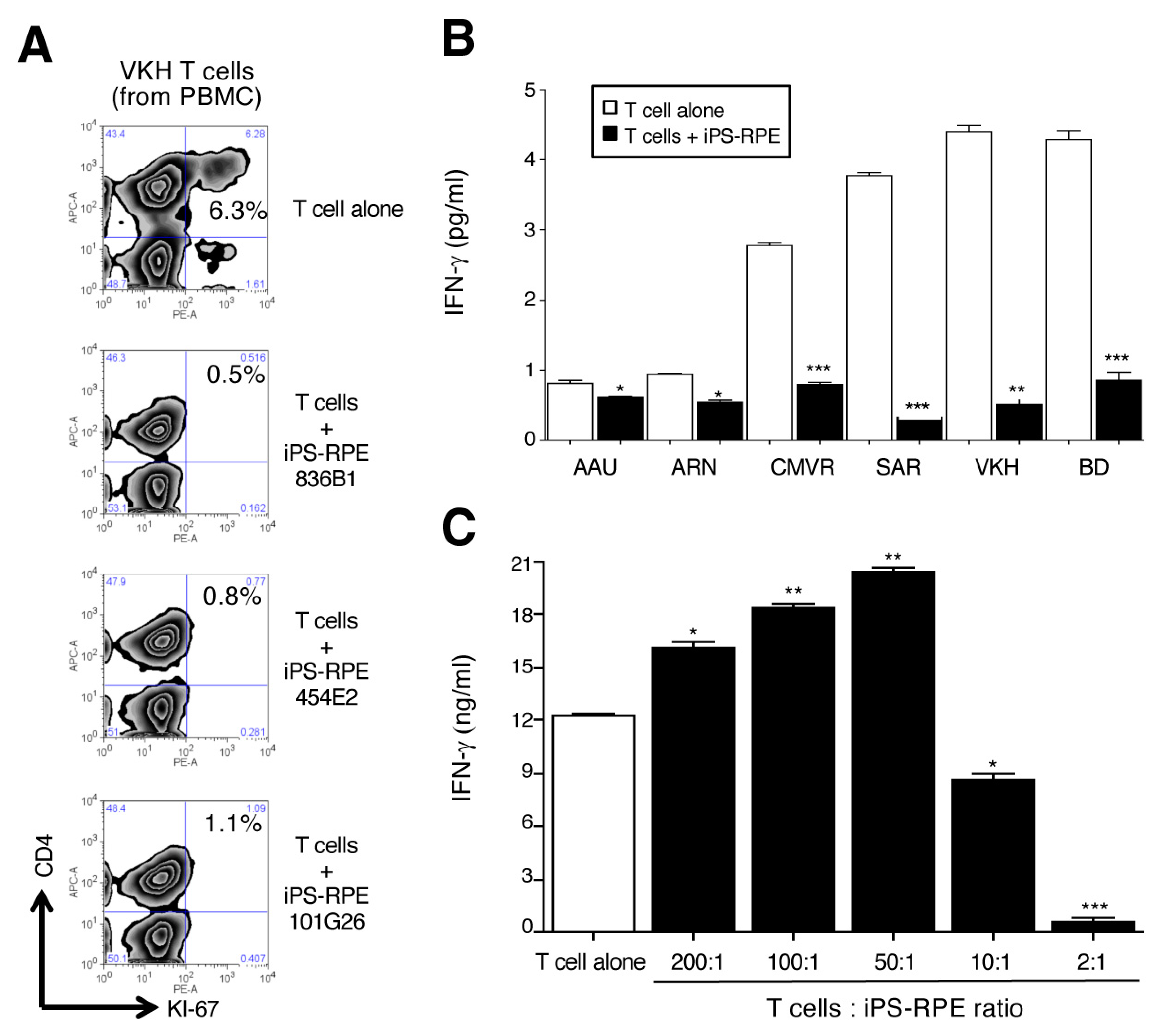
2.1. Ability of IPS-Derived RPE Cells to Suppress or Activate T Cells from Active Uveitis Patients
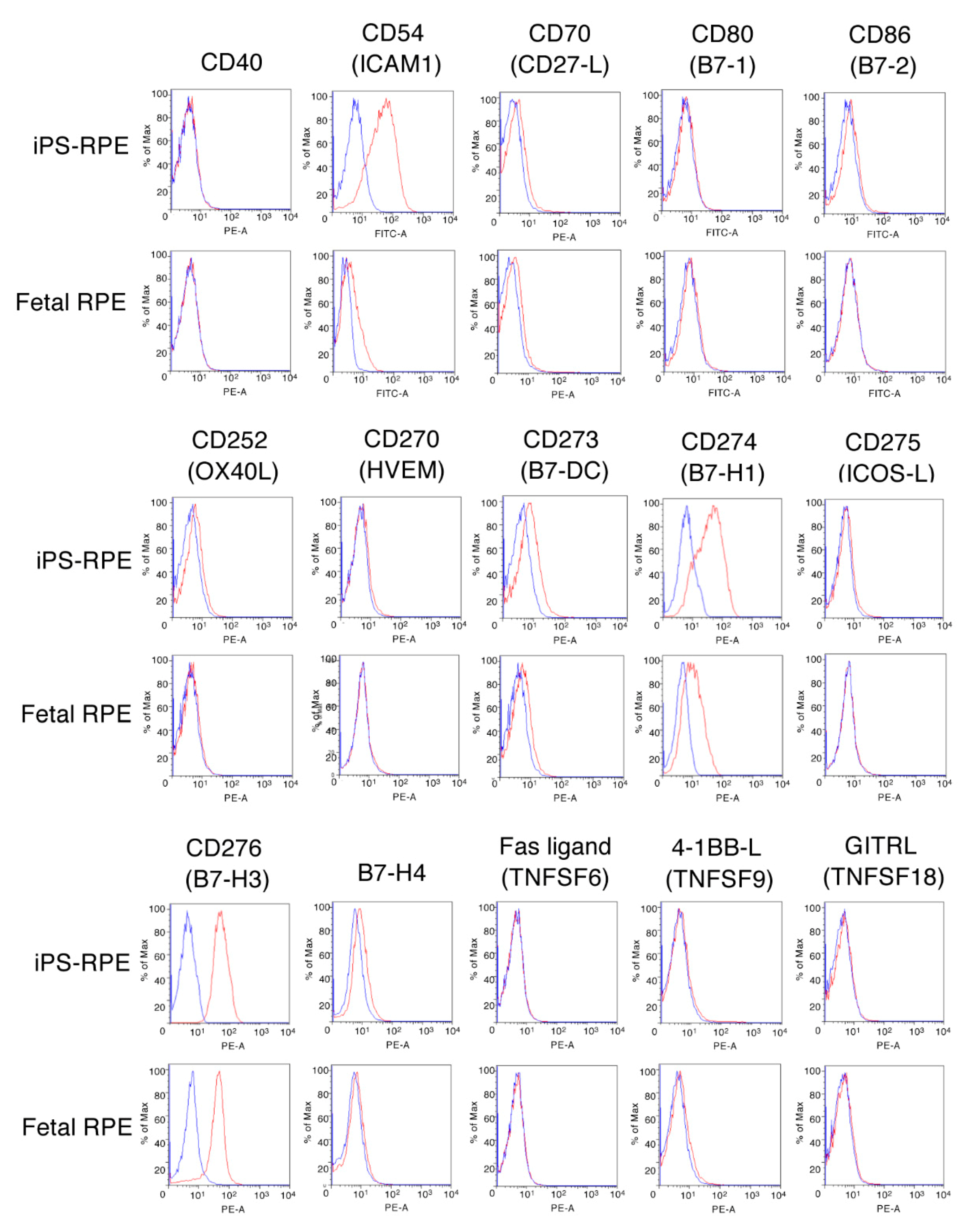
2.2. Survey of Candidate Immunosuppressive Molecules on IPS-RPE Cells
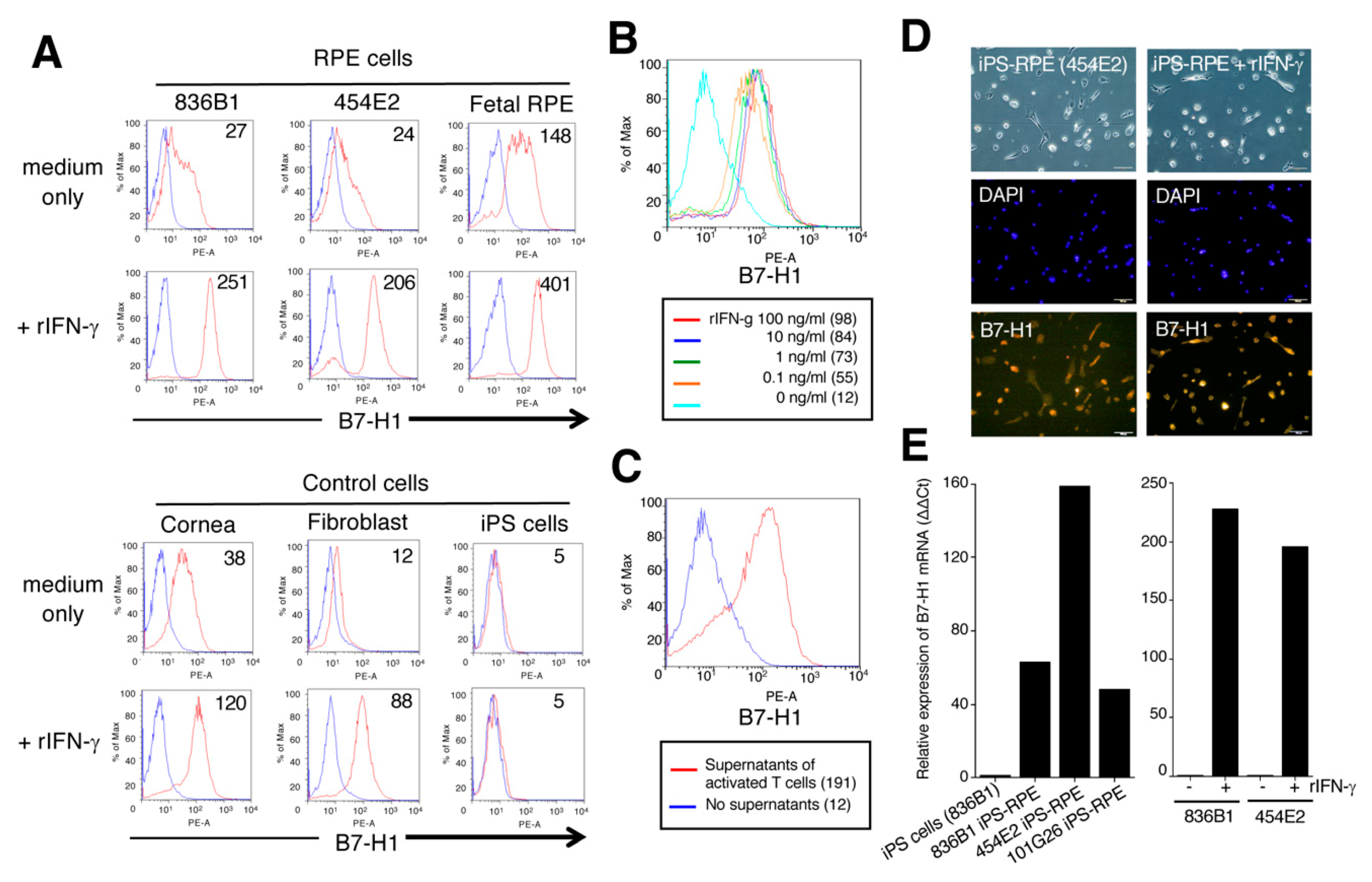
2.3. Expression of B7-H1/CD274 and B7-DC/CD273 on Human IPS-RPE Cells
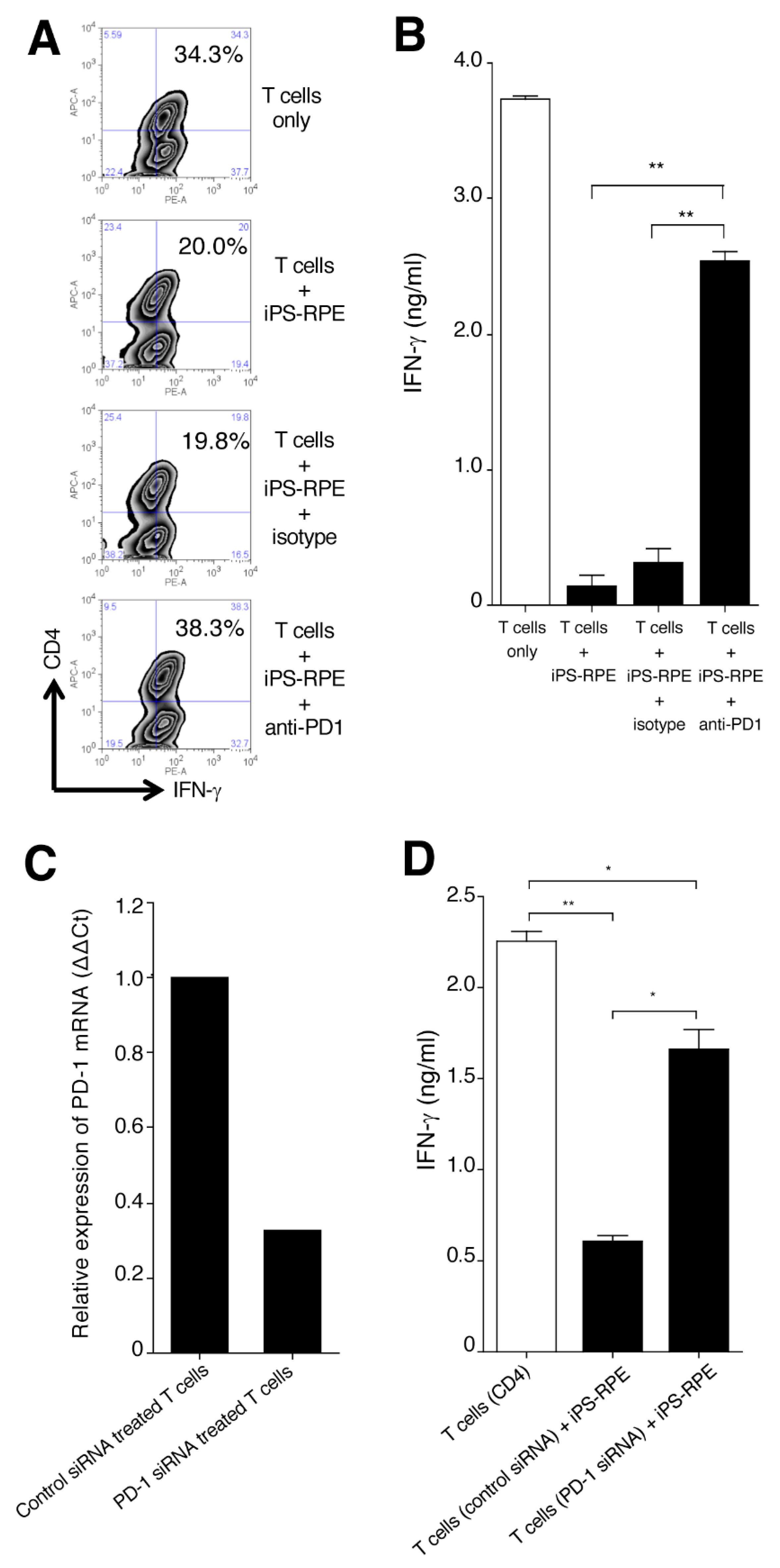
2.4. Expression of PD-1 on Uveitis T Cells
2.5. Capacity of T-cell Suppression by Cocultures with Anti-PD-1 Antibody or PD-1 SiRNA
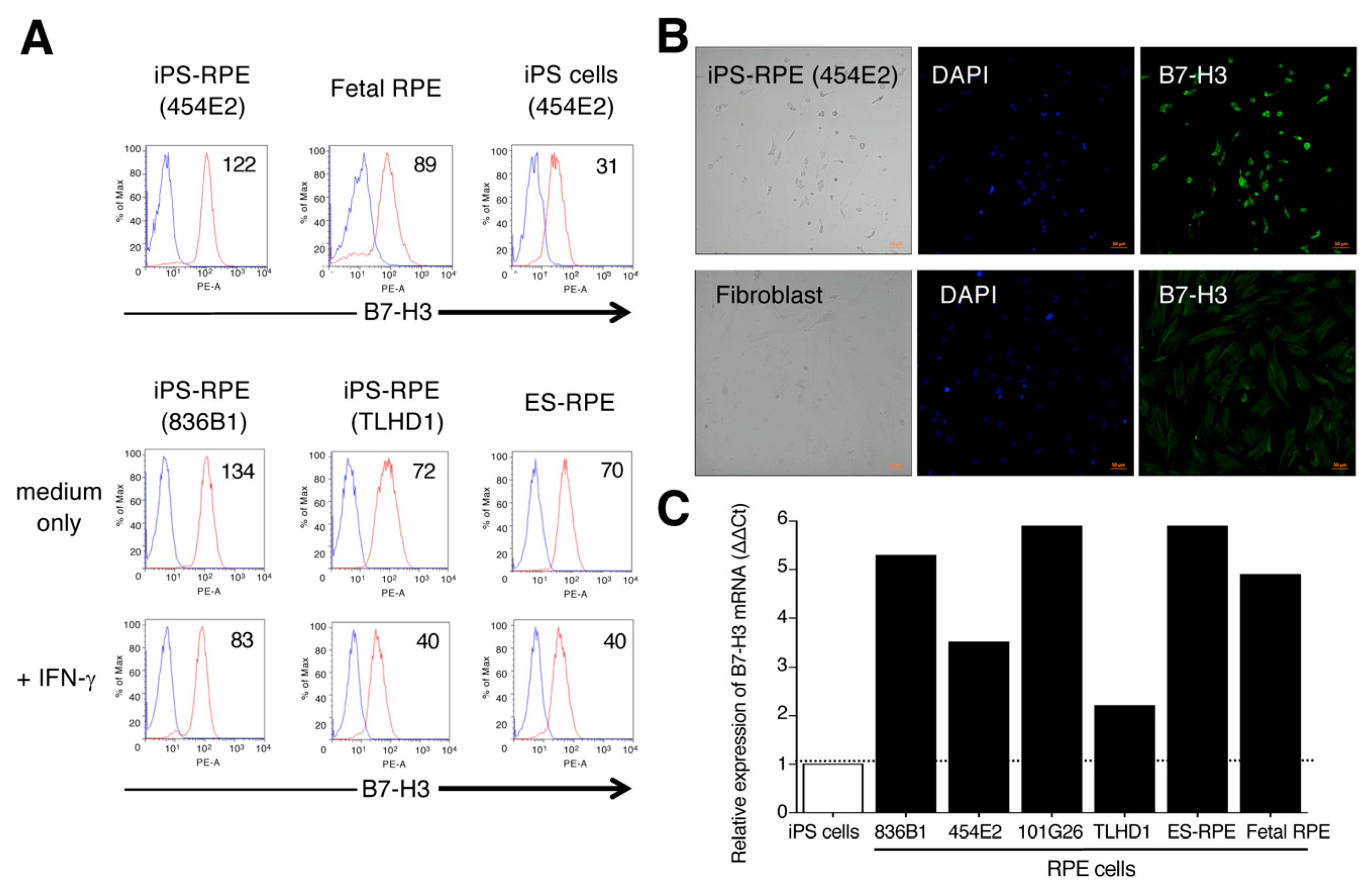
2.6. Expression of B7-H3/CD276 on Human IPS-RPE Cells
2.7. Expression of TLT-2 on Uveitis T Cells
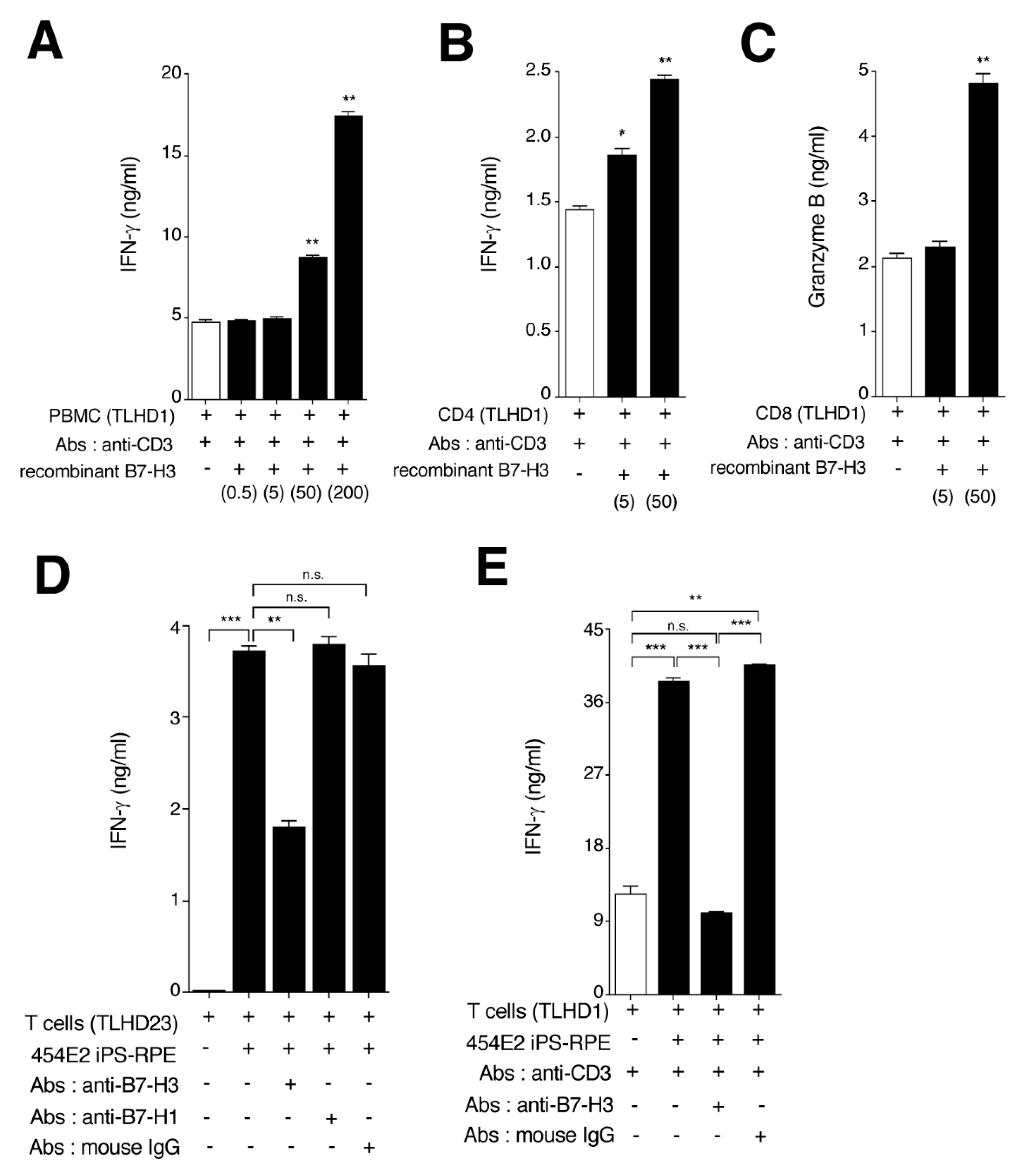
2.8. RPE Cells Derived from IPS Cells Can Activate Bystander T Cells via B7-H3 Costimulatory Signals
3. Discussion
4. Material and Methods
4.1. Establishment of IPS-Derived RPE (iPS-RPE) Cells
4.2. Establishment of T Cells from Uveitis Patients
4.3. Flow Cytometry
4.4. Quantitative RT-PCR
4.5. Transfection of SiRNA
4.6. Immunohistochemistry
4.7. Recombinant Proteins
4.8. Statistical Evaluation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FACS | fluorescence-activated cell sorting |
| IHC | immunohistochemistry |
| iPSC | induced pluripotent stem cells |
| PBMC | peripheral blood mononuclear cells |
| PD-1 | programmed death 1 |
| PD-L1 | programmed death-ligand 1 |
| RPE | retinal pigment epithelial cells |
| TLT-2 | Trem-like transcript 2 |
| VKH | Vogt-Koyanagi-Harada |
References
- Niederkorn, J.Y. Immune privilege in the anterior chamber of the eye. Crit. Rev. Immunol. 2002, 22, 13–46. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S. Role of ocular pigment epithelial cells in immune privilege. Arch. Immunol. Ther. Exp. 2009, 57, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W. Ocular immune privilege: Therapeutic opportunities from an experiment of nature. Nat. Rev. Immunol. 2003, 3, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Algvere, P.V. Clinical possibilities in retinal pigment epithelial transplantations. Acta Ophthalmol. Scand. 1997, 75, 1. [Google Scholar] [CrossRef]
- Peyman, G.A.; Blinder, K.J.; Paris, C.L.; Alturki, W.; Nelson, N.C., Jr.; Desai, U. A technique for retinal pigment epithelium transplantation for age-related macular degeneration secondary to extensive subfoveal scarring. Ophthalmic Surg. 1991, 22, 102–108. [Google Scholar] [CrossRef]
- Weisz, J.M.; Humayun, M.S.; De Juan, E.J.; Del Cerro, M.; Sunness, J.S.; Dagnelie, G.; Soylu, M.; Rizzo, L.; Nussenblatt, R.B. Allogenic fetal retinal pigment epithelial cell transplant in a patient with geographic atrophy. Retina 1999, 19, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Fujii, S.; Sugita, S.; Futatsugi, Y.; Ishida, M.; Edo, A.; Makabe, K.; Kamao, H.; Iwasaki, Y.; Sakaguchi, H.; Hirami, Y.; et al. A strategy for personalized treatment of iPS-retinal immune rejections assessed in cynomolgus monkey models. Int. J. Mol. Sci. 2020, 21, 3077. [Google Scholar] [CrossRef]
- Sugita, S.; Mandai, M.; Hirami, Y.; Takagi, S.; Maeda, T.; Fujihara, M.; Matsuzaki, M.; Yamamoto, M.; Iseki, K.; Hayashi, N.; et al. HLA-matched allogeneic iPS cells-derived RPE Transplantation for macular degeneration. J. Clin. Med. 2020, 9, 2217. [Google Scholar] [CrossRef]
- Sugita, S.; Futagami, Y.; Smith, S.B.; Naggar, H.; Mochizuki, M. Retinal and ciliary body pigment epithelium suppress activation of T lymphocytes via transforming growth factor beta. Exp. Eye Res. 2006, 83, 1459–1471. [Google Scholar] [CrossRef]
- Sugita, S.; Horie, S.; Nakamura, O.; Futagami, Y.; Takase, H.; Keino, H.; Aburatani, H.; Katunuma, N.; Ishidoh, K.; Yamamoto, Y.; et al. Retinal pigment epithelium-derived CTLA-2alpha induces TGFbeta-producing T regulatory cells. J. Immunol. 2008, 181, 7525–7536. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Kamao, H.; Iwasaki, Y.; Okamoto, S.; Hashiguchi, T.; Iseki, K.; Hayashi, N.; Mandai, M.; Takahashi, M. Inhibition of T-cell activation by retinal pigment epithelial cells derived from induced pluripotent stem cells. Investig. Ophthalmol Vis. Sci. 2015, 56, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Forrester, J.V.; Xu, H. Synthesis of complement factor H by retinal pigment epithelial cells is down-regulated by oxidized photoreceptor outer segments. Exp. Eye Res. 2007, 84, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Juel, H.B.; Kaestel, C.; Folkersen, L.; Faber, C.; Heegaard, N.H.; Borup, R.; Nissen, M.H. Retinal pigment epithelial cells upregulate expression of complement factors after co-culture with activated T cells. Exp. Eye Res. 2011, 92, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Makabe, K.; Fujii, S.; Takahashi, M. Detection of complement activators in immune attack eyes after iPS-derived retinal pigment epithelial cell transplantation. Investig. Ophthalmol Vis. Sci. 2018, 59, 4198–4209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juel, H.B.; Faber, C.; Udsen, M.S.; Folkersen, L.; Nissen, M.H. Chemokine expression in retinal pigment epithelial ARPE-19 cells in response to coculture with activated T cells. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8472–8480. [Google Scholar] [CrossRef] [Green Version]
- Devine, L.; Lightman, S.L.; Greenwood, J. Role of LFA-1, ICAM-1, VLA-4 and VCAM-1 in lymphocyte migration across retinal pigment epithelial monolayers in vitro. Immunology 1996, 88, 456–462. [Google Scholar] [CrossRef]
- Sugita, S.; Iwasaki, Y.; Makabe, K.; Kimura, T.; Futagami, T.; Suegami, S.; Takahashi, M. Lack of T cell response to iPSC-derived retinal pigment epithelial cells from HLA homozygous donors. Stem Cell Rep. 2016, 7, 619–634. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Iwasaki, Y.; Makabe, K.; Kamao, H.; Mandai, M.; Shiina, T.; Ogasawara, K.; Hirami, Y.; Kurimoto, Y.; Takahashi, M. Successful transplantation of retinal pigment epithelial cells from MHC homozygote iPSCs in MHC-matched models. Stem Cell Rep. 2016, 7, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Usui, Y.; Horie, S.; Futagami, Y.; Aburatani, H.; Okazaki, T.; Honjo, T.; Takeuchi, M.; Mochizuki, M. T-cell suppression by programmed cell death 1 ligand 1 on retinal pigment epithelium during inflammatory conditions. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2862–2870. [Google Scholar] [CrossRef]
- Usui, Y.; Okunuki, Y.; Hattori, T.; Kezuka, T.; Keino, H.; Ebihara, N.; Sugita, S.; Usui, M.; Goto, H.; Takeuchi, M. Functional expression of B7H1 on retinal pigment epithelial cells. Exp. Eye Res. 2008, 86, 52–59. [Google Scholar] [CrossRef]
- Owaki, T.; Asakawa, M.; Morishima, N.; Hata, K.; Fukai, F.; Matsui, M.; Mizuguchi, J.; Yoshimoto, T. A role for IL-27 in early regulation of Th1 differentiation. J. Immunol. 2005, 175, 2191–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, S.Y.; Otsuji, M.; Gorski, K.; Huang, X.; Slansky, J.E.; Pai, S.I.; Shalabi, A.; Shin, T.; Pardoll, D.M.; Tsuchiya, H. B7-DC, a new dendritic cell molecule with potent costimulatory properties for T cells. J. Exp. Med. 2001, 193, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A costimulatory molecule for T cell activation and IFN-gamma production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Suh, W.K.; Gajewska, B.U.; Okada, H.; Gronski, M.A.; Bertram, E.M.; Dawicki, W.; Duncan, G.S.; Bukczynski, J.; Plyte, S.; Elia, A.; et al. The B7 family member B7-H3 preferentially down-regulates T helper type 1-mediated immune responses. Nat. Immunol. 2003, 4, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Nurieva, R.I.; Prasad, D.V. Immune regulation by novel costimulatory molecules. Immunol. Res. 2003, 28, 39–48. [Google Scholar] [CrossRef]
- King, R.G.; Herrin, B.R.; Justement, L.B. Trem-like transcript 2 is expressed on cells of the myeloid/granuloid and B lymphoid lineage and is up-regulated in response to inflammation. J. Immunol. 2006, 176, 6012–6021. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.W.; McVicar, D.W. TREM and TREM-like receptors in inflammation and disease. Curr. Opin. Immunol. 2009, 21, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Leitner, J.; Klauser, C.; Pickl, W.F.; Stöckl, J.; Majdic, O.; Bardet, A.F.; Kreil, D.P.; Dong, C.; Yamazaki, T.; Zlabinger, G.; et al. B7-H3 is a potent inhibitor of human T-cell activation: No evidence for B7-H3 and TREML2 interaction. Eur. J. Immunol. 2009, 39, 1754–1764. [Google Scholar] [CrossRef]
- Takase, H.; Sugita, S.; Taguchi, C.; Imai, Y.; Mochizuki, M. Capacity of ocular infiltrating T helper type 1 cells of patients with non-infectious uveitis to produce chemokines. Br. J. Ophthalmol. 2006, 90, 765–768. [Google Scholar] [CrossRef] [Green Version]
- Takase, H.; Futagami, Y.; Yoshida, T.; Kamoi, K.; Sugita, S.; Imai, Y.; Mochizuki, M. Cytokine profile in aqueous humor and sera of patients with infectious or noninfectious uveitis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1557–1561. [Google Scholar] [CrossRef] [Green Version]
- Carreno, B.M.; Collins, M. The B7 family of ligands and its receptors: New pathways for costimulation and inhibition of immune responses. Annu. Rev. Immunol. 2002, 20, 29–53. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhu, G.; Tamada, K.; Chen, L. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 1999, 5, 1365–1369. [Google Scholar] [CrossRef] [PubMed]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Usui, Y.; Horie, S.; Futagami, Y.; Yamada, Y.; Ma, J.; Kezuka, T.; Hamada, H.; Usui, T.; Mochizuki, M.; et al. Human corneal endothelial cells expressing programmed death-ligand 1 (PD-L1) suppress PD-1 + T helper 1 cells by a contact-dependent mechanism. Investig. Ophthalmol. Vis. Sci. 2009, 50, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Ng, T.F.; Schwartzkopff, J.; Streilein, J.W. CTLA-4+CD8+ T cells that encounter B7-2+ iris pigment epithelial cells express their own B7-2 to achieve global suppression of T cell activation. J. Immunol. 2004, 172, 4184–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugita, S.; Ng, T.F.; Lucas, P.J.; Gress, R.E.; Streilein, J.W. B7+ iris pigment epithelium induce CD8+ T regulatory cells; both suppress CTLA-4+ T cells. J. Immunol. 2006, 176, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugita, S.; Keino, H.; Futagami, Y.; Takase, H.; Mochizuki, M.; Stein-Streilein, J.; Streilein, J.W. B7+ iris pigment epithelial cells convert T cells into CTLA-4+, B7-expressing CD8+ regulatory T cells. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5376–5384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, J.; Wang, M.; Miyashita, M.; Tanemoto, K.; Takahashi, H.; Takemori, T.; Okumura, K.; Yagita, H.; Azuma, M. B7-H1-induced apoptosis as a mechanism of immune privilege of corneal allografts. J. Immunol. 2006, 177, 5928–5935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Fraser, C.C.; Kikly, K.; Wells, A.D.; Han, R.; Coyle, A.J.; Chen, L.; Hancock, W.W. B7-H3 promotes acute and chronic allograft rejection. Eur. J. Immunol. 2005, 35, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Theillac, C.; Straub, M.; Breton, A.L.; Thomas, L.; Dalle, S. Bilateral uveitis and macular edema induced by Nivolumab: A case report. BMC Ophthalmol. 2017, 17, 227. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Lam, W.C.; Chen, L. Recurrent grade 4 panuveitis with serous retinal detachment related to nivolumab treatment in a patient with metastatic renal cell carcinoma. Cancer Immunol. Immunother. 2019, 68, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Obata, S.; Saishin, Y.; Teramura, K.; Ohji, M. Vogt-Koyanagi-Harada Disease-Like Uveitis during Nivolumab (Anti-PD-1 Antibody) Treatment for Metastatic Cutaneous Malignant Melanoma. Case Rep. Ophthalmol. 2019, 10, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Hanna, K.S. A rare case of pembrolizumab-induced uveitis in a patient with metastatic melanoma. Pharmacotherapy 2016, 36, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Yamasaki, O. Vogt-Koyanagi-Harada disease-like posterior uveitis in the course of nivolumab (anti-PD-1 antibody), interposed by vemurafenib (BRAF inhibitor), for metastatic cutaneous malignant melanoma. Clin. Case Rep. 2017, 5, 694–700. [Google Scholar] [CrossRef]
- Osakada, F.; Jin, Z.B.; Hirami, Y.; Ikeda, H.; Danjyo, T.; Watanabe, K.; Sasai, Y.; Takahashi, M. In vitro differentiation of retinal cells from human pluripotent stem cells by small-molecule induction. J. Cell Sci. 2009, 122, 3169–3179. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Takase, H.; Taguchi, C.; Imai, Y.; Kamoi, K.; Kawaguchi, T.; Sugamoto, Y.; Futagami, Y.; Itoh, K.; Mochizuki, M. Ocular infiltrating CD4+ T cells from patients with Vogt-Koyanagi-Harada disease recognize human melanocyte antigens. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2547–2554. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugita, S.; Futatsugi, Y.; Ishida, M.; Edo, A.; Takahashi, M. Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem (iPS) Cells Suppress or Activate T Cells via Costimulatory Signals. Int. J. Mol. Sci. 2020, 21, 6507. https://doi.org/10.3390/ijms21186507
Sugita S, Futatsugi Y, Ishida M, Edo A, Takahashi M. Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem (iPS) Cells Suppress or Activate T Cells via Costimulatory Signals. International Journal of Molecular Sciences. 2020; 21(18):6507. https://doi.org/10.3390/ijms21186507
Chicago/Turabian StyleSugita, Sunao, Yoko Futatsugi, Masaaki Ishida, Ayaka Edo, and Masayo Takahashi. 2020. "Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem (iPS) Cells Suppress or Activate T Cells via Costimulatory Signals" International Journal of Molecular Sciences 21, no. 18: 6507. https://doi.org/10.3390/ijms21186507
APA StyleSugita, S., Futatsugi, Y., Ishida, M., Edo, A., & Takahashi, M. (2020). Retinal Pigment Epithelial Cells Derived from Induced Pluripotent Stem (iPS) Cells Suppress or Activate T Cells via Costimulatory Signals. International Journal of Molecular Sciences, 21(18), 6507. https://doi.org/10.3390/ijms21186507