A Single-Tube HNB-Based Loop-Mediated Isothermal Amplification for the Robust Detection of the Ostreid herpesvirus 1


 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
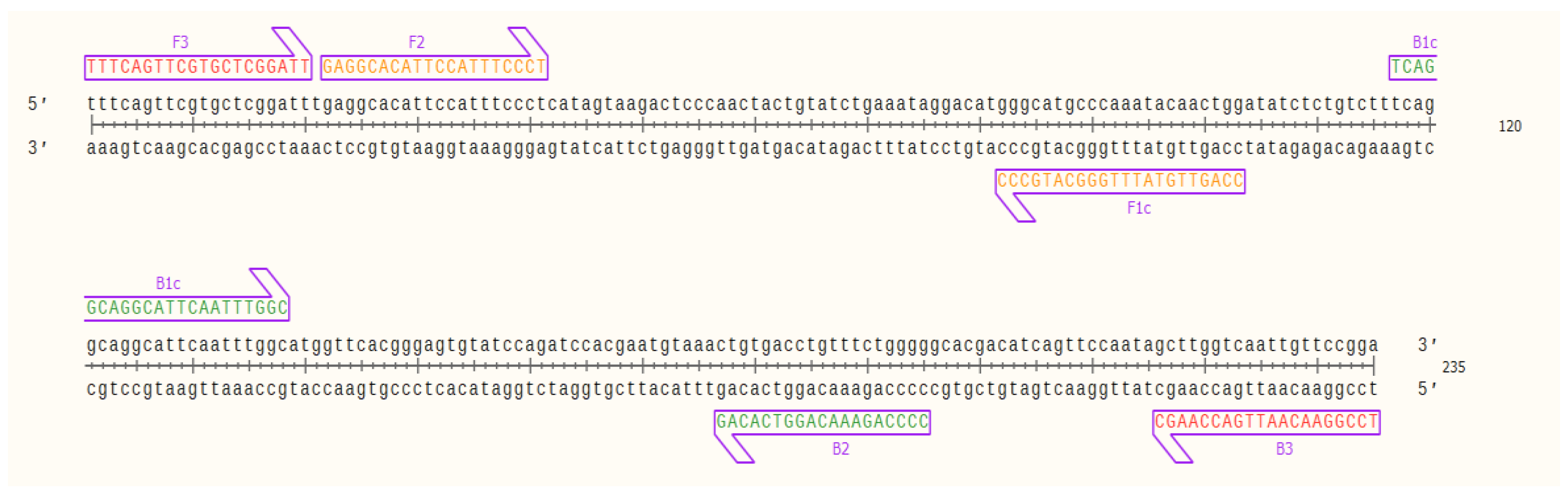
2.1. OsHV-1 Specific LAMP Primer Design
2.2. Bst DNA Polymerase LAMP Assay Optimization
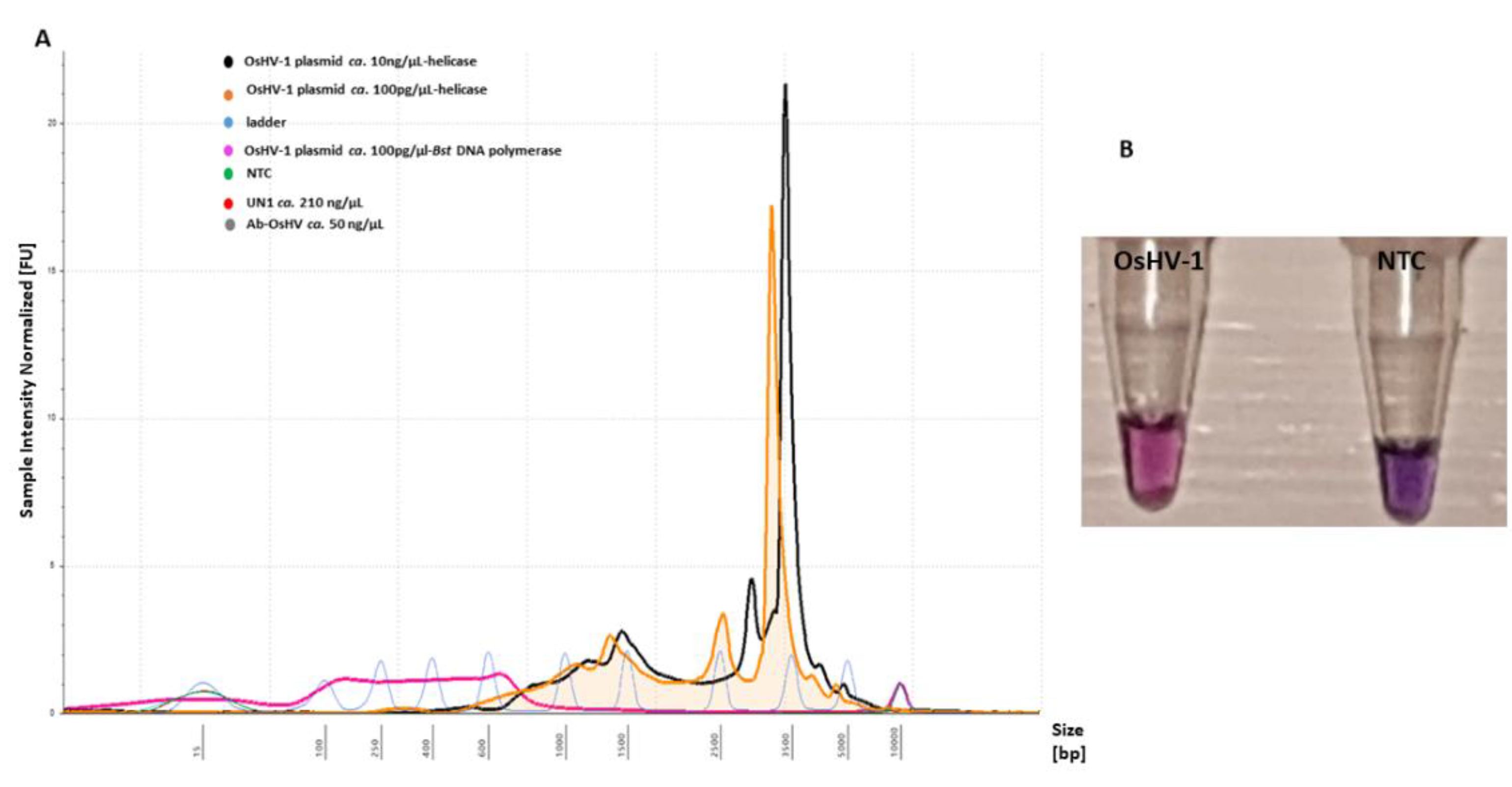
2.3. Sensitivity and Applicability of LAMP Assay
2.4. Tte UvrD Helicase LAMP Assay
3. Discussion
4. Materials and Methods
4.1. Samples Preparation, DNA Extraction and qPCR
4.2. DNA Sequencing Using MinION Device and Data Analysis
4.3. Designing and Assessment of Synthetic DNA of OsHV-1 and AbHV
4.4. LAMP Primers Design and in Silico Specificity
4.5. Optimization of LAMP Conditions
4.5.1. Bst DNA Polymerase LAMP Assay
4.5.2. Tte UvrD Helicase Lamp Assay
4.6. Sensitivity and Applicability of LAMP Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LAMP | Loop-mediated isothermal amplification |
| HDA | Helicase-dependent isothermal amplification |
| HNB | Hydroxynaphtol blue |
| OsHV-1 | Ostreid herpesvirus 1 |
| AbHV | Abalone herpesvirus 1 |
| PCR | Polymerase chain reaction |
| ORF | Open reading frame |
References
- Aiewsakun, P.; Simmonds, P. The genomic underpinnings of eukaryotic virus taxonomy: Creating a sequence-based framework for family-level virus classification. Microbiome 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Pellet, P.E.; Roizman, B. The family Herpesviridae: A brief introduction. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 2479–2499. [Google Scholar]
- Hellberg, T.; Paßvogel, L.; Schulz, K.S.; Klupp, B.G.; Mettenleiter, T.C. Nuclear egress of Herpesviruses: The prototypic vesicular nucleocytoplasmic transport. Adv. Virus Res. 2016, 94, 81–140. [Google Scholar] [PubMed]
- Lionel, D.; Guyader, T.; Tourbiez, D.; Pépin, J.F. Is horizontal transmission of the Ostreid herpesvirus OsHV-1 in Crassostrea gigas affected by unselected or selected survival status in adults to juveniles? Aquaculture 2013, 408–409, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Rosani, U.; Venier, P. Oyster RNA-seq data support the development of Malacoherpesviridae genomics. Front. Microbiol. 2017, 8, 1515. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Segarra, A.; Pépin, J.F.; Arzul, I.; Morga, B.; Faury, N.; Renault, T. Detection and description of a particular Ostreid herpesvirus 1 genotype associated with massive mortality outbreaks of Pacific oysters, Crassostrea gigas, in France in 2008. Virus Res. 2010, 153, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Chen, H.; Renault, T.; Cai, Y.; Bai, C.; Wang, C.; Huang, J. Complete genome sequence of Acute viral necrosis virus associated with massive mortality outbreaks in the Chinese scallop, Chlamys farreri. Virol. J. 2013, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Qi, R.J.; Yang, R.; Han, T.; Jiang, J.Z.; Wang, J.Y. The prevalence of Abalone herpesvirus in two Haliotis species in South China during 2002–2013. Aquaculture 2019, 505, 18–26. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Bean, T.P.; Hooper, C.H.; Stenton, C.A.; Sanders, M.B.; Paley, R.K.; Rastas, P.; Bryrom, M.; Matika, O.; Houston, D. A Genome-Wide Association Study for Host Resistance to Ostreid Herpesvirus in Pacific Oysters (Crassostrea gigas). Genes gEnom. Genet. 2018, 8, 1273–1280. [Google Scholar] [CrossRef] [Green Version]
- Carnegie, R.B.; Arzul, I.; Bushek, D. Managing marine mollusc diseases in the context of regional and international commerce: Policy issues and emerging concerns. Phil. Trans. R. Soc. B 2016, 371, 20150215. [Google Scholar] [CrossRef] [Green Version]
- Hambrey, J.; Evans, S. SR694 Aquaculture in England, Wales and Northern Ireland: An Analysis of the Economic Contribution and Value of the Major Sub-Sectors and the Most Important Farmed Species. Final Report on Seafish. Available online: https://www.seafish.org/media/publications/FINALISED_Aquaculture_in_EWNI_FINALISED__-_Sept_2016.pdf (accessed on 1 September 2016).
- OIE. Manual of Diagnostic Tests for Aquatic Animals. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/aahm/current/chapitre_ostreid_herpesvirus_1.pdf (accessed on 14 November 2019).
- Gao, F.; Jiang, J.Z.; Wang, J.Y.; Wei, H.Y. Real-time quantitative isothermal detection of Ostreid herpesvirus-1 DNA in Scapharca subcrenata using recombinase polymerase amplification. J. Virol. Methods 2018, 255, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Arzul, I.; Nicolas, J.L.; Davison, A.J.; Renault, T. French scallops: A new host for Ostreid herpesvirus 1. Virology 2001, 290, 342–349. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Oyster Mortality. EFSA Panel on Animal Health and Welfare. Available online: https://efsa.onlinelibrary.wiley.com/doi/pdf/10.2903/j.efsa.2015.4122 (accessed on 4 June 2015).
- Pepin, J.F.; Riou, A.; Renault, T. Rapid and sensitive detection of Ostreid herpesvirus 1 in oyster samples by real-time PCR. J. Virol. Methods 2008, 149, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martenot, C.; Faury, N.; Morga, B.; Degremont, L.; Lamy, J.B.; Houssin, M.; Renault, T. Exploring first interactions between Ostreid Herpesvirus 1 (OsHV-1) and its host, Crassostrea gigas: Effects of specific antiviral antibodies and dextran sulfate. Front. Microbiol. 2019, 10, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadkar, V.J.; Goldfarb, D.M.; Gantt, S.; Tilley, P.A.G. Real-time detection and monitoring of loop mediated amplification (LAMP) reaction using self-quenching and de-quenching fluorogenic probes. Sci. Rep. 2018, 8, 5548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, M.; Honda, E.; Ogura, A.; Nomoto, A.; Hanaki, K.I. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. Biotechniques 2009, 46, 167–172. [Google Scholar] [CrossRef]
- Hill, J.; Beriwal, S.; Chandra, I.; Paul, V.K.; Kapil, A.; Singh, T.; Wadowsky, R.M.; Singh, V.; Goyal, A.; Jahnukainen, T.; et al. Loop-mediated isothermal amplification assay for rapid detection of common strains of Escherichia coli. J. Clin. Microbiol. 2008, 46, 2800–2804. [Google Scholar] [CrossRef] [Green Version]
- Zhao, V.X.T.; Wong, T.I.; Zheng, X.T.; Tan, Y.N.; Zhou, H. Colorimetric biosensors for point-of-care virus detection. Mater. Sci. Energy Technol. 2020, 3, 237–249. [Google Scholar] [CrossRef]
- Ren, W.; Renault, T.; Cai, Y.; Wang, C. Development of a loop-mediated isothermal amplification assay for rapid and sensitive detection of Ostreid herpesvirus 1 DNA. J. Virol. Methods 2010, 170, 30–36. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Abbadi, M.; Zamperin, G.; Gastaldelli, M.; Pascoli, F.; Rosani, U.; Milani, A.; Schivo, A.; Rossetti, E.; Turolla, E.; Gennari, L.; et al. Identification of a newly described OsHV-1 µvar from the North Adriatic Sea (Italy). J. Gen. Virol. 2018, 99, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.; Duran, C.; Berkman, P.J.; Lorenc, M.T.; Stiller, J.; Manoli, S.; Hayden, M.; Forrest, K.; Fleury, D.; Baumann, U.; et al. Single nucleotide polymorphism discovery from wheat next-generation sequence data. Plant Biotechnol. J. 2012, 10, 743–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockweiler, T. Loop-Mediated Isothermal Amplification (LAMP) and Primer Optimization. Available online: https://www.lucigen.com/docs/slide-decks/Loop-Mediated-Isothermal-Amplification-Design-Primers-Webinar-March-2018.pdf (accessed on 18 March 2018).
- Meagher, R.J.; Priye, A.; Light, Y.K.; Huang, C.; Wang, E. Impact of primer dimers and self-amplifying hairpins on reverse transcription loop-mediated isothermal amplification detection of viral RNA. Analyst 2018, 143, 1924–1933. [Google Scholar] [CrossRef] [PubMed]
- Jansson, L.; Hedman, J. Challenging the proposed causes of the PCR plateau phase. Biomol. Detect. Quantif. 2019, 17, 100082. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Suh, S.-S.; Park, M.; Oh, M.J.; Kim, J.O.; Lee, S.; Lee, T.-K. Detection of coat protein gene of nervous necrosis virus using loop-mediated isothermal amplification. Asian Pac. J. Trop. Med. 2016, 9, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Fishbach, J.; Xander, N.C.; Frohme, M.; Glökler, J.F. Shining a light on LAMP assays’ A comparison of LAMP visualization methods including the novel use of berberine. BioTechniques 2018, 58, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, K.; Zhao, X.; Ding, X.; Li, X.D.; Zou, S.M.; Guo, J.F.; Wang, D.Y.; Gao, R.B.; Li, X.Y.; Huang, W.J.; et al. Visual detection of human infection with influenza A (H7N9) virus by subtype-specific reverse transcription loop-mediated isothermal amplification with hydroxynaphthol blue dye. Clin. Microbiol. Infect. 2013, 19, 372–375. [Google Scholar] [CrossRef] [Green Version]
- Chahar, M.; Anvikar, A.; Valecha, N. Development and evaluation of a novel HNB based isothermal amplification assay for fast detection of pyrimethamine resistance (S108N) in Plasmodium falciparum. Int. J. Environ. Res. Public Health 2019, 16, 1635. [Google Scholar] [CrossRef] [Green Version]
- Teng, P.H.; Chen, C.L.; Sung, P.F.; Lee, F.C.; Ou, B.R.; Lee, P.Y. Specific detection of reverse transcription-loop-mediated isothermal amplification amplicons for Taura syndrome virus by colorimetric dot-blot hybridization. J. Virol. Mthods 2007, 146, 317–326. [Google Scholar] [CrossRef]
- Diessi, N.; Ernst, U.; Schultz, A.; Wolf, S.; Graf, E. Quality Control in Illumina Sequencing Workflows Using the TapeStation System. Available online: https://www.agilent.com/cs/library/applications/application-Illumina-workflow-qulality-control-5994-0327EN-agilent.pdf (accessed on 23 October 2018).
- Barreda-García, S.; Miranda-Castro, R.; de-Los-Santos-Álvarez, N.; Miranda-Ordieres, A.J.; Lobo-Castañón, M.J. Helicase-dependent isothermal amplification: A novel tool in the development of molecular-based analytical systems for rapid pathogen detection. Anal. Bioanal. Chem. 2018, 410, 679–693. [Google Scholar] [CrossRef]
- New England Biolabs Inc. Available online: https://international.neb.com/protocols/2019/01/16/protocol-for-unwinding-double-stranded-dna-with-tte-uvrd-helicase-m1202 (accessed on 16 January 2019).
- Toldrà, A.; Andree, K.B.; Bertomeu, E.; Roque, A.; Carrasco, N.; Gairín, I.; Furones, M.D.; Campàs, M. Rapid capture and detection of Ostreid herpesvirus-1 from Pacific oyster Crassostrea gigas and seawater using magnetic beads. PLoS ONE 2018, 13, e0205207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, S.C.; Fidler, A.; Renault, T. Primers for PCR-based detection of Ostreid herpesvirus-1 (OsHV-1): Application in a survey of New Zealand mollusks. Aquaculture 2007, 272, 126–139. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Designed Primer/Sequence (5′–3′) | Length (bp) | GC (%) | GC Clamp |
|---|---|---|---|---|
| OsHV-LP-F | F3/TTTCAGTTCGTGCTCGGATT | 20 | 45.0 | 2 |
| OsHV-LP-B | B3/TCCGGAACAATTGACCAAGC | 20 | 50.0 | 3 |
| OsHV-LP-FIP 1 | F1c/CCAGTTGTATTTGGGCATGCCC | 46 | 54.5 | 3 |
| F2/GAGGCACATTCCATTTCCCT | ||||
| OsHV-LP-BIP 2 | B1c/TCAGGCAGGCATTCAATTTGGC | 45 | 48.9 | 3 |
| B2/CCCCAGAAACAGGTCACAG |
| No. | Reagent | Stock Concentration | Concentration of Tested Reagents/ 1 µL Reaction Volume | ||
|---|---|---|---|---|---|
| 1. | MgSO4 (New England BioLabs) | 100 mM | 4 mM | 6 mM | 8 mM * |
| 2. | dNTP Mix (Thermo Fisher) | 10 mM | n/a | 1.4 mM | 1.6 mM |
| 3. | FIP/BIP Primers (Thermo Fisher) | 40 µM | 0.4 µM | 0.8 µM | 1.6 µM |
| 4. | F3/B3 Primers (Thermo Fisher) | 5 µM | 0.05 µM | 0.1 µM | 0.2 µM |
| 5. | HNB (Merck) | 20 mM | 120 µM | 148 µM | 400 µM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaczek-Moczydłowska, M.A.; Mohamed-Smith, L.; Toldrà, A.; Hooper, C.; Campàs, M.; Furones, M.D.; Bean, T.P.; Campbell, K. A Single-Tube HNB-Based Loop-Mediated Isothermal Amplification for the Robust Detection of the Ostreid herpesvirus 1. Int. J. Mol. Sci. 2020, 21, 6605. https://doi.org/10.3390/ijms21186605
Zaczek-Moczydłowska MA, Mohamed-Smith L, Toldrà A, Hooper C, Campàs M, Furones MD, Bean TP, Campbell K. A Single-Tube HNB-Based Loop-Mediated Isothermal Amplification for the Robust Detection of the Ostreid herpesvirus 1. International Journal of Molecular Sciences. 2020; 21(18):6605. https://doi.org/10.3390/ijms21186605
Chicago/Turabian StyleZaczek-Moczydłowska, Maja A., Letitia Mohamed-Smith, Anna Toldrà, Chantelle Hooper, Mònica Campàs, M. Dolors Furones, Tim P. Bean, and Katrina Campbell. 2020. "A Single-Tube HNB-Based Loop-Mediated Isothermal Amplification for the Robust Detection of the Ostreid herpesvirus 1" International Journal of Molecular Sciences 21, no. 18: 6605. https://doi.org/10.3390/ijms21186605
APA StyleZaczek-Moczydłowska, M. A., Mohamed-Smith, L., Toldrà, A., Hooper, C., Campàs, M., Furones, M. D., Bean, T. P., & Campbell, K. (2020). A Single-Tube HNB-Based Loop-Mediated Isothermal Amplification for the Robust Detection of the Ostreid herpesvirus 1. International Journal of Molecular Sciences, 21(18), 6605. https://doi.org/10.3390/ijms21186605