Integrated Analysis to Study the Relationship between Tumor-Associated Selenoproteins: Focus on Prostate Cancer


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
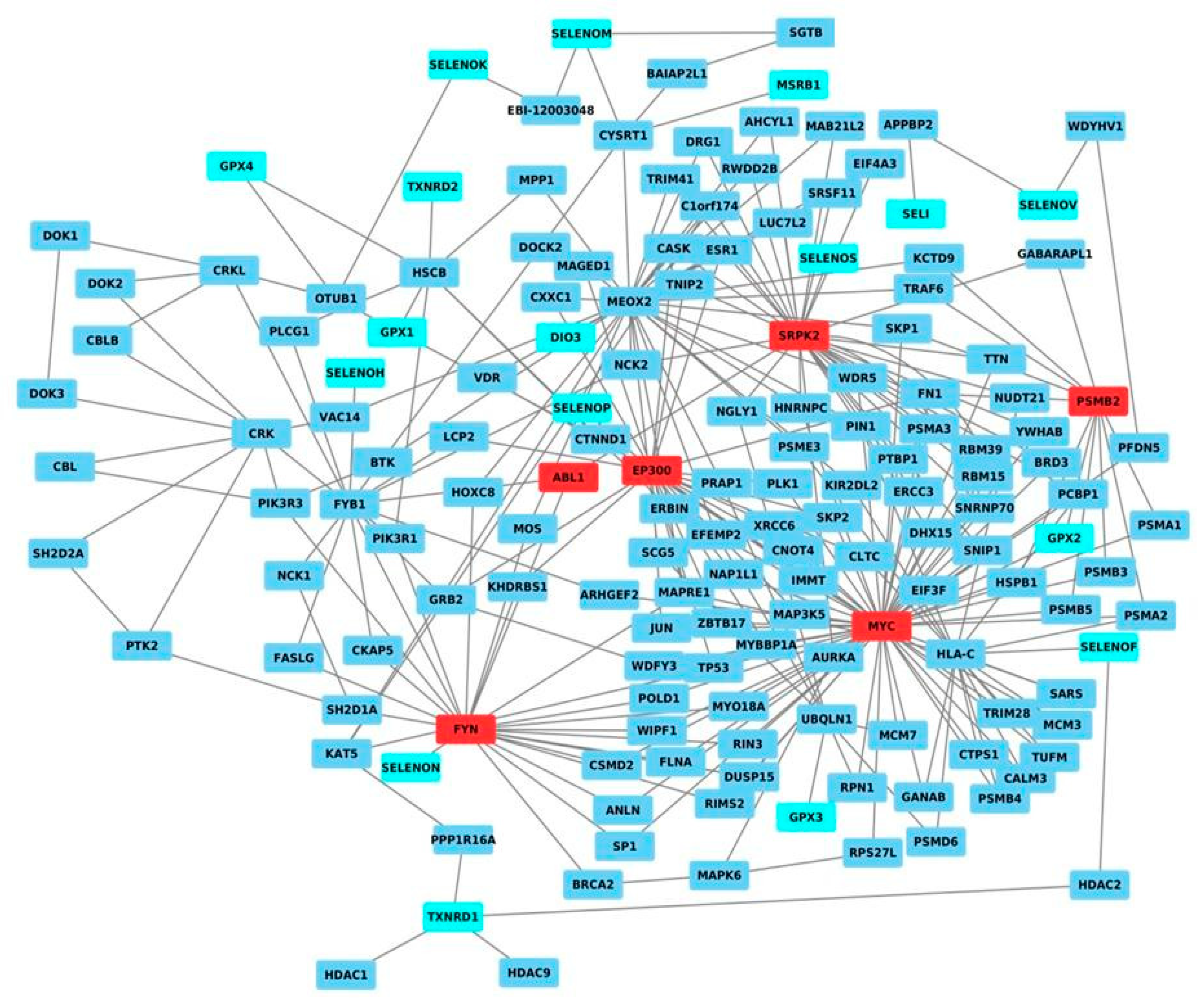
2.1. Network Analysis
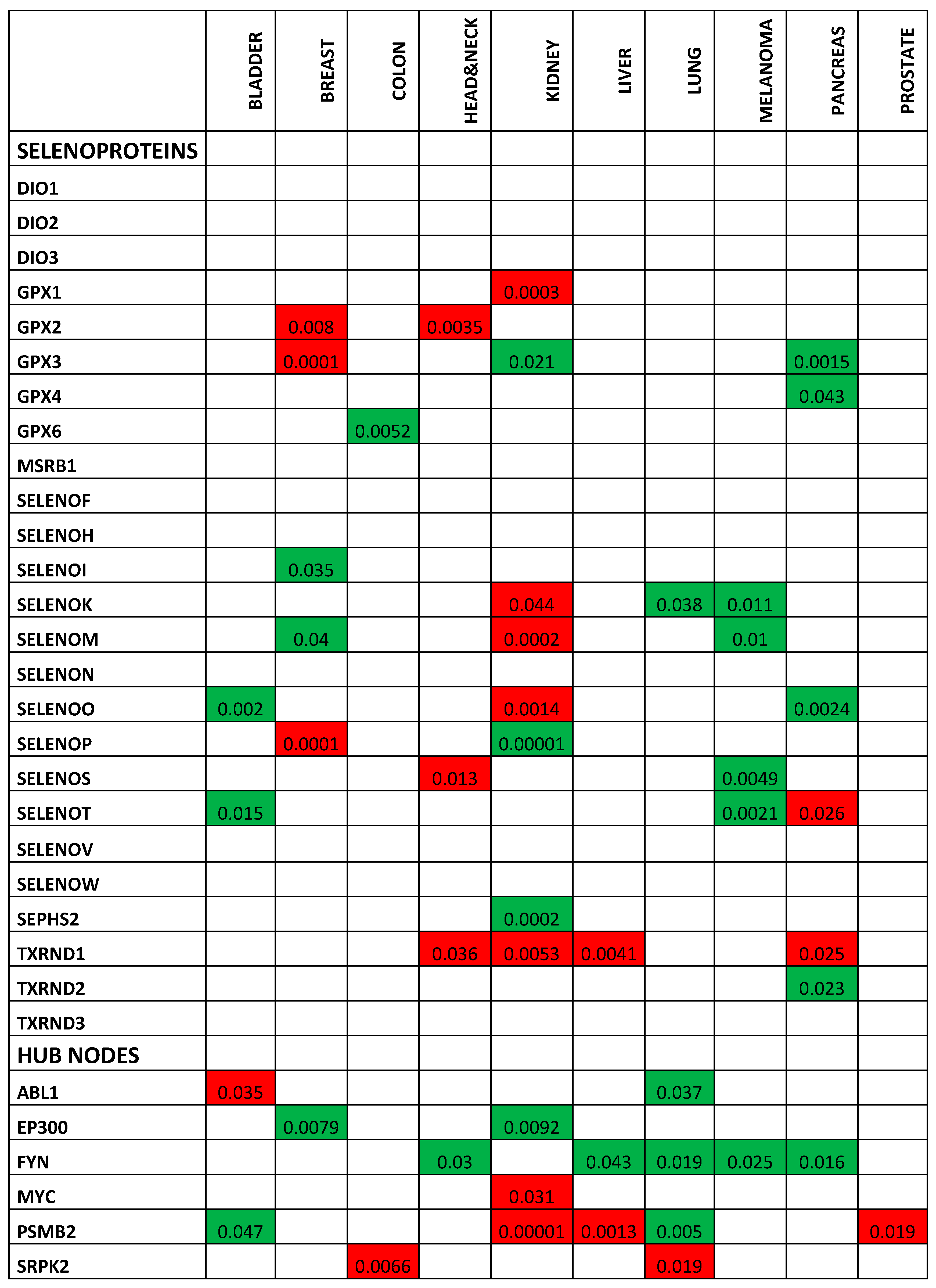
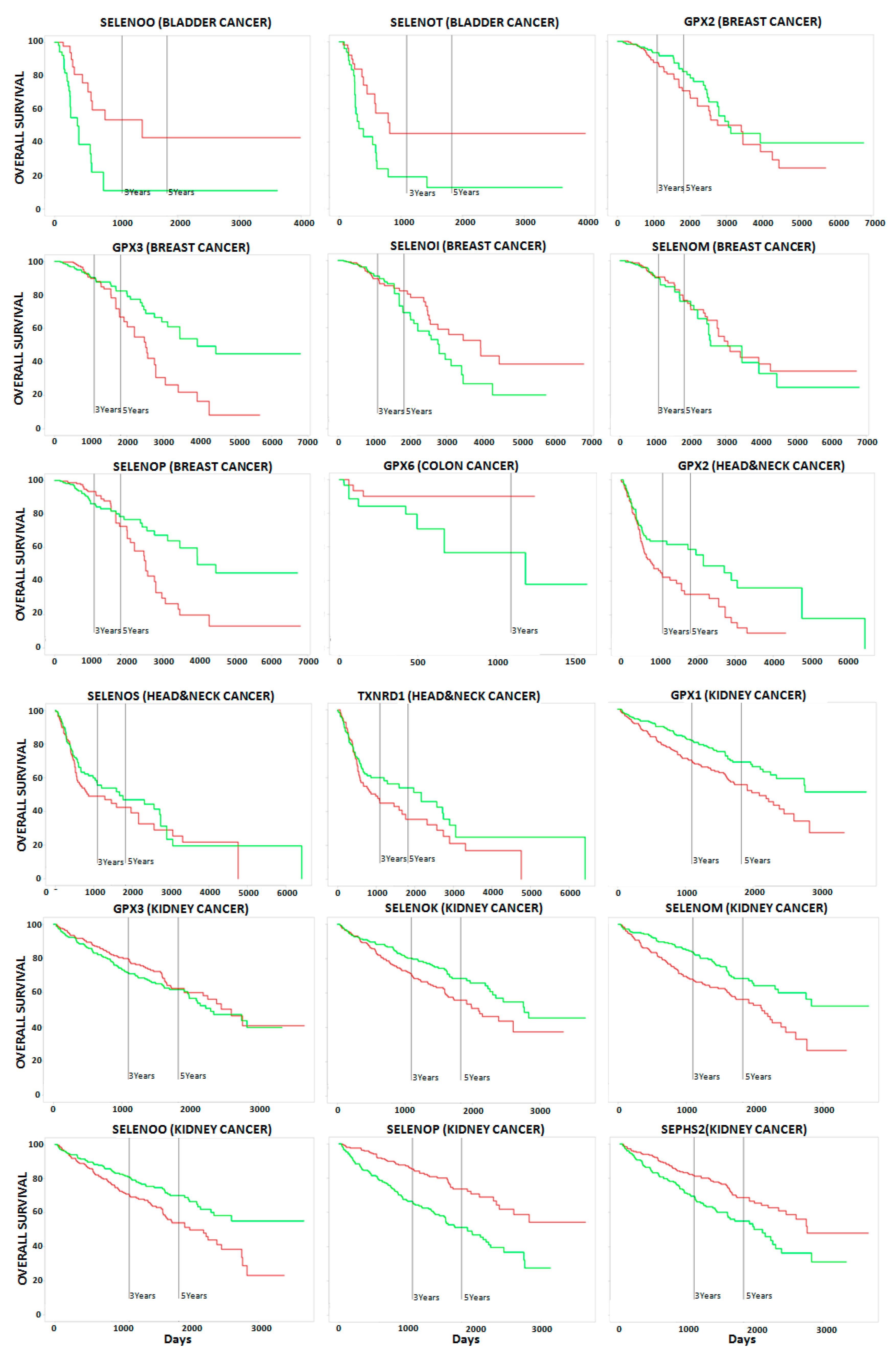
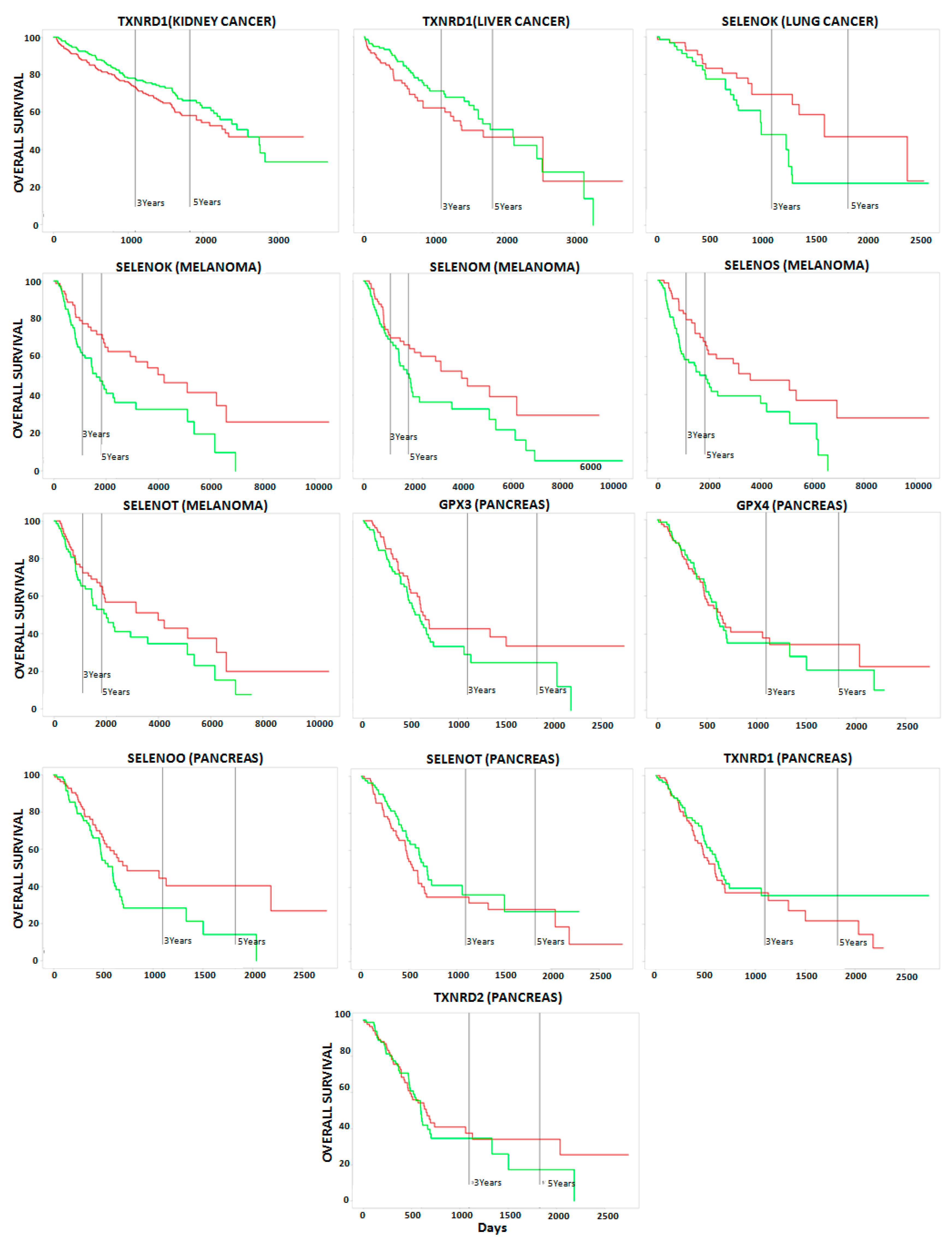
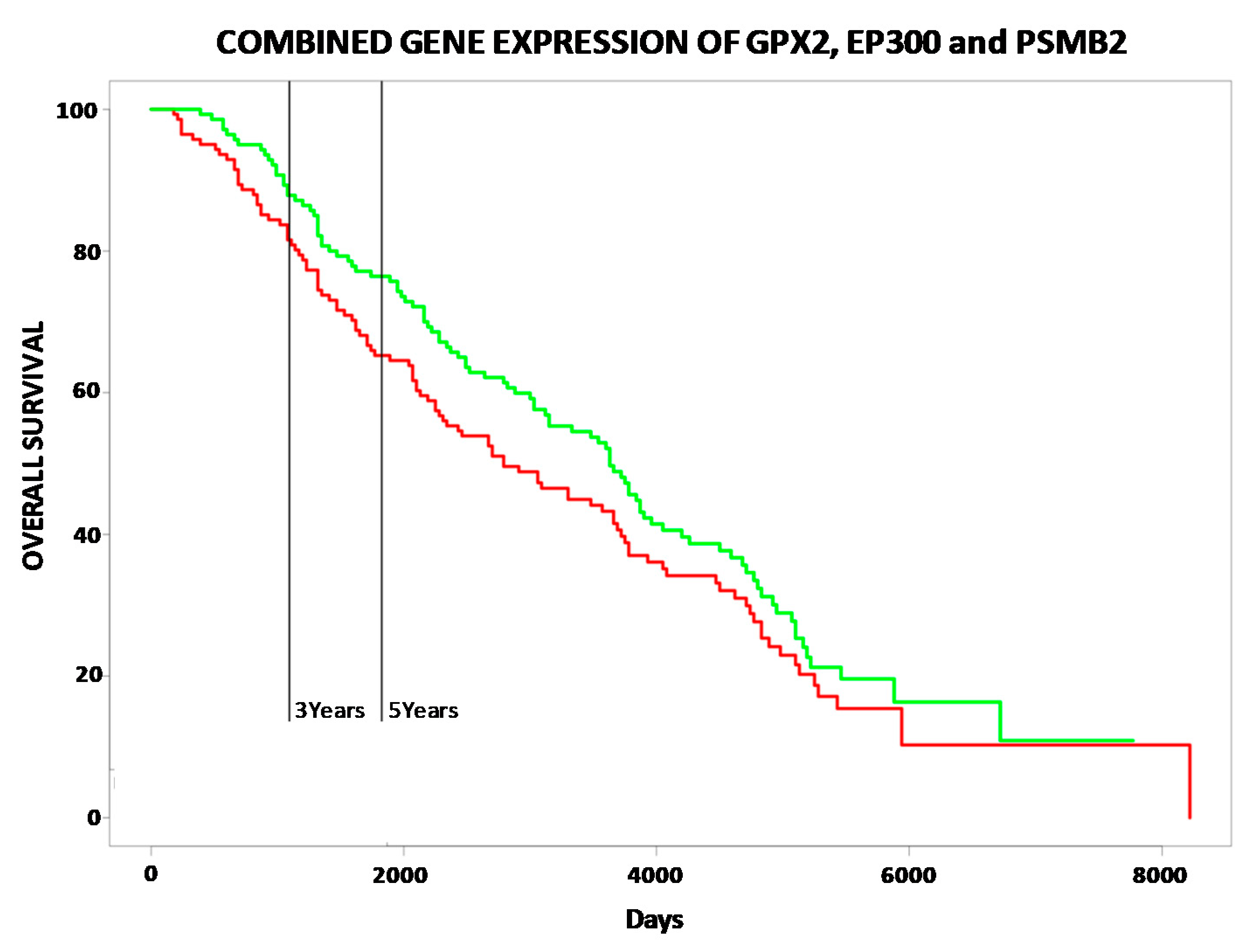
2.2. Correlation between Selenoproteins/HUB Nodes and Cancer Patient Overall Survival
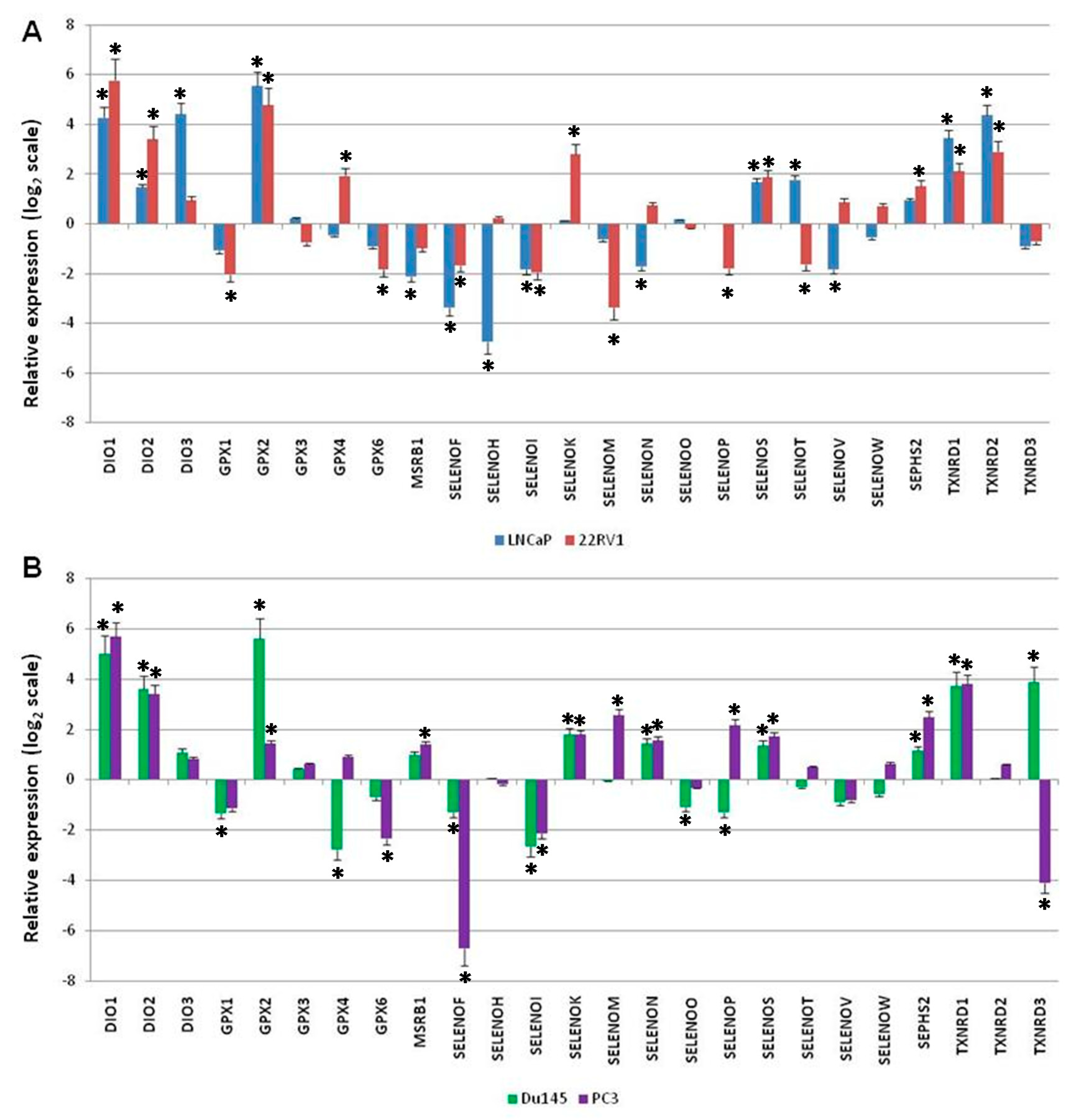
2.3. Gene Expression Levels of Twenty-Five Selenoproteins in Prostate Cancer Cells Compared to Normal Prostate Cells
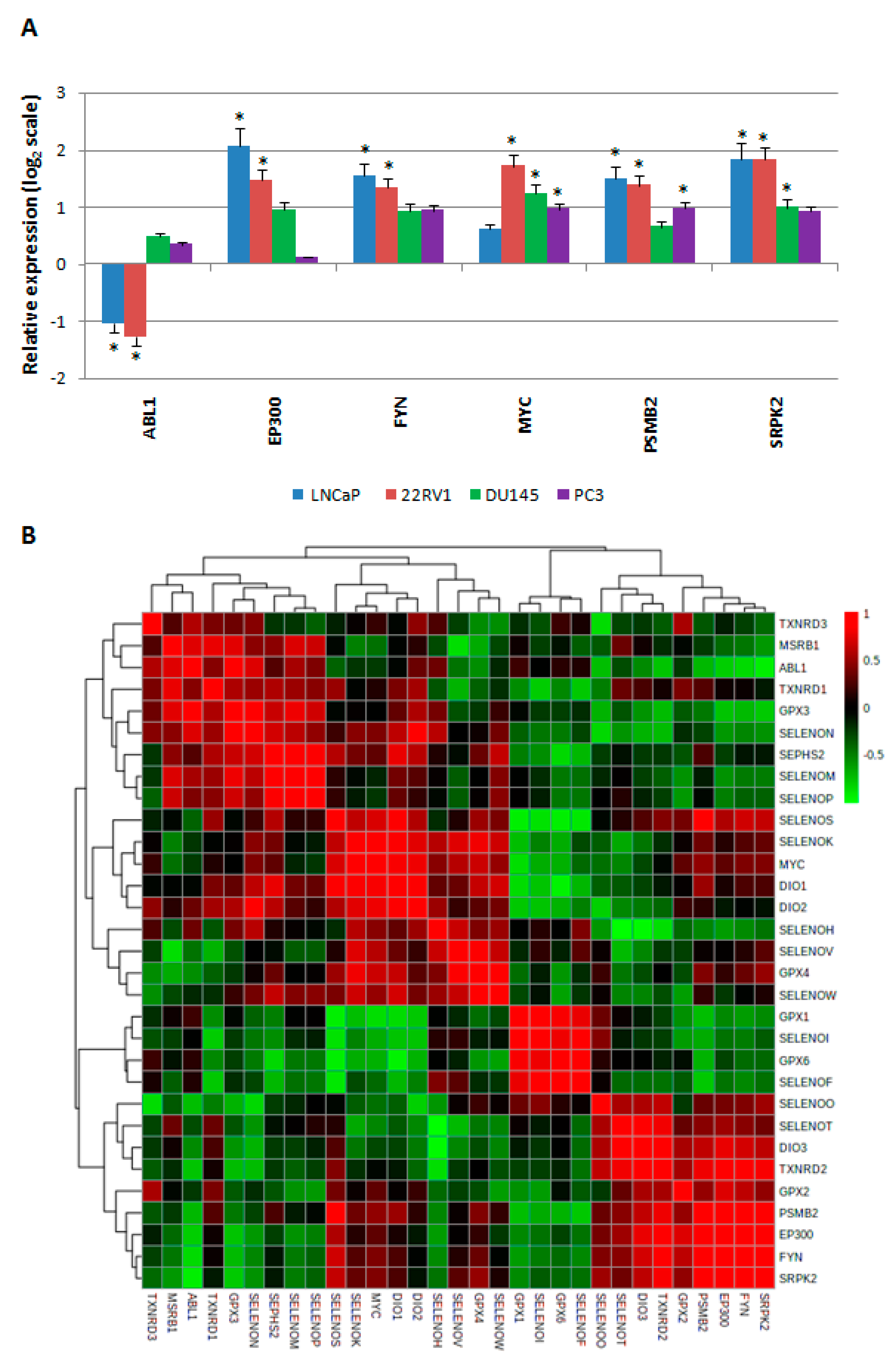
2.4. Gene Expression Levels of HUB Nodes in Prostate Cancer Cells, and Their Correlation with Selenoprotein Expression
3. Discussion
4. Materials and Methods
4.1. Network Analysis
4.2. Survival Gene Analysis
4.3. Prostate Cancer Cell Lines
4.4. RNA Preparation and Reverse Transcription-qPCR (RT-qPCR)
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diamond, A.M. The subcellular location of selenoproteins and the impact on their function. Nutrients 2015, 7, 3938–3948. [Google Scholar] [CrossRef] [Green Version]
- Reszka, E.; Jablonska, E.; Gromadzinska, J.; Wasowicz, W. Relevance of selenoprotein transcripts for selenium status in humans. Genes Nutr. 2012, 7, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Chen, Q.; Zeng, F.; Zeng, L.; Yang, H.; Xiong, Y.; Zhou, C.; Liu, L.; Jiang, W.; Yang, N.; et al. The serum activity of thioredoxin reductases 1 (TrxR1) is correlated with the poor prognosis in EGFR wild-type and ALK negative non-small cell lung cancer. Oncotarget 2017, 8, 115270–115279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlamova, E.G. Participation of selenoproteins localized in the ER in the processes occurring in this organelle and in the regulation of carcinogenesis-associated processes. J. Trace Elem. Med. Biol. 2018, 48, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Castets, P.; Lescure, A.; Guicheney, P.; Allamand, V. Selenoprotein N in skeletal muscle: From diseases to function. J. Mol. Med. 2012, 90, 1095–1107. [Google Scholar] [CrossRef]
- Short, S.P.; Williams, C.S. Selenoproteins in tumorigenesis and cancer progression. Adv. Cancer Res. 2017, 136, 49–83. [Google Scholar]
- Cheng, W.H.; Prabhu, K.S. Special Issue of “Optimal Selenium Status and Selenoproteins in Health”. Biol. Trace Elem. Res. 2019, 192, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Lubiński, J.; Marciniak, W.; Muszynska, M.; Jaworowska, E.; Sulikowski, M.; Jakubowska, A.; Kaczmarek, K.; Sukiennicki, G.; Falco, M.; Baszuk, P.; et al. Serum selenium levels and the risk of progression of laryngeal cancer. PLoS ONE 2018, 13, e0184873. [Google Scholar] [CrossRef] [Green Version]
- Lubinski, J.; Marciniak, W.; Muszynska, M.; Huzarski, T.; Gronwald, J.; Cybulski, C.; Jakubowska, A.; Debniak, T.; Falco, M.; Kladny, J.; et al. Serum selenium levels predict survival after breast cancer. Breast Cancer Res. Treat. 2018, 167, 591–598. [Google Scholar] [CrossRef]
- Peters, K.M.; Carlson, B.A.; Gladyshev, V.N.; Tsuji, P.A. Selenoproteins in colon cancer. Free Radic. Biol. Med. 2018, 127, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.M. Selenoproteins of the Human Prostate: Unusual Properties and Role in Cancer Etiology. Biol. Trace Elem. Res. 2019, 192, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Shi, Q.Q.; Ge, L.Y.; Zhuang, Q.F.; Xue, D.; Xu, H.Y.; He, X.Z. Selenoprotein M stimulates the proliferative and metastatic capacities of renal cell carcinoma through activating the PI3K/AKT/mTOR pathway. Cancer Med. 2019, 8, 4836–4844. [Google Scholar] [CrossRef] [PubMed]
- Marciel, M.P.; Hoffmann, P.R. Molecular Mechanisms by Which Selenoprotein K Regulates Immunity and Cancer. Biol. Trace Elem. Res. 2019, 192, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Men, L.; Sun, J.; Ren, D. Deficiency of VCP-Interacting Membrane Selenoprotein (VIMP) Leads to G1 Cell Cycle Arrest and Cell Death in MIN6 Insulinoma Cells. Cell. Physiol. Biochem. 2018, 51, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, E.; Accardo, M.; Capone, F.; Colonna, G.; Castello, G.; Costantini, S. Assessment of the Selenoprotein M (SELM) over-expression on human hepatocellular carcinoma tissues by immunohistochemistry. Eur. J. Histochem. 2014, 58, 2433. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, E.; Capone, F.; Accardo, M.; Sorice, A.; Costantini, M.; Colonna, G.; Castello, G.; Costantini, S. GPX4 and GPX7 over-expression in human hepatocellular carcinoma tissues. Eur. J. Histochem. 2015, 59, 2540. [Google Scholar] [CrossRef]
- Guariniello, S.; Di Bernardo, G.; Colonna, G.; Cammarota, M.; Castello, G.; Costantini, S. Evaluation of the selenotranscriptomeexpression in twohepatocellular carcinoma celllines. Anal. Cell. Pathol. 2015, 2015, 419561. [Google Scholar] [CrossRef] [Green Version]
- Potenza, N.; Castiello, F.; Panella, M.; Colonna, G.; Ciliberto, G.; Russo, A.; Costantini, S. Human MiR-544a Modulates SELK Expression in Hepatocarcinoma Cell Lines. PLoS ONE 2016, 11, e0156908. [Google Scholar] [CrossRef]
- Rusolo, F.; Capone, F.; Pasquale, R.; Angiolillo, A.; Colonna, G.; Castello, G.; Costantini, M.; Costantini, S. Comparison of the seleno-transcriptome expression between human non-cancerous mammary epithelial cells and two human breast cancer cell lines. Oncol. Lett. 2017, 13, 2411–2417. [Google Scholar] [CrossRef]
- Nunziata, C.; Polo, A.; Sorice, A.; Capone, F.; Accardo, M.; Guerriero, E.; Marino, F.Z.; Orditura, M.; Budillon, A.; Costantini, S. Structural analysis of human SEPHS2 protein, a selenocysteine machinery component, over-expressed in triple negative breast cancer. Sci. Rep. 2019, 9, 16131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Costantini, S.; Colonna, G. The protein-protein interaction network of the human Sirtuin family. Biochim. Biophys. Acta 2013, 1834, 1998–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Acharya, A.; Mattheos, N.; Li, S.; Ziebolz, D.; Schmalz, G.; Haak, R.; Schmidt, J.; Sun, Y. Molecular mechanisms linking peri-implantitis and type 2 diabetes mellitus revealed by transcriptomic analysis. Peer J. 2019, 7, e7124. [Google Scholar] [CrossRef]
- Covani, U.; Marconcini, S.; Derchi, G.; Barone, A.; Giacomelli, L. Relationship between human periodontitis and type 2 diabetes at a genomic level: A data-mining study. J. Periodontol. 2009, 80, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, L.; Nicolini, C. Gene expression of human T lymphocytes cell cycle: Experimental and bioinformatic analysis. J. Cell. Biochem. 2006, 99, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, V.; Giacomelli, L.; Sivozhelezov, V.; Degauque, N.; Lair, D.; Soulillou, J.-P.; Pechkova, E.; Nicolini, C.; Brouard, S. AKT1 leader gene and downstream targets are involved in a rat model of kidney allograft tolerance. J. Cell. Biochem. 2010, 111, 709–719. [Google Scholar] [CrossRef]
- Barone, A.; Toti, P.; Giuca, M.R.; Derchi, G.; Covani, U. A gene network bioinformatics analysis for pemphigoid autoimmune blistering diseases. Clin. Oral Investig. 2015, 19, 1207–1222. [Google Scholar] [CrossRef]
- Milone, M.R.; Pucci, B.; Bruzzese, F.; Carbone, C.; Piro, G.; Costantini, S.; Capone, F.; Leone, A.; Di Gennaro, E.; Caraglia, M.; et al. Acquired resistance to zoledronic acid and the parallel acquisition of an aggressive phenotype are mediated by p38-MAP kinase activation in prostate cancer cells. Cell Death Dis. 2013, 4, e641. [Google Scholar] [CrossRef] [Green Version]
- Bizzarro, V.; Belvedere, R.; Milone, M.R.; Pucci, B.; Lombardi, R.; Bruzzese, F.; Popolo, A.; Parente, L.; Budillon, A.; Petrella, A. Annexin A1 is involved in the acquisition and maintenance of a stem cell-like/aggressive phenotype in prostate cancer cells with acquired resistance to zoledronic acid. Oncotarget 2015, 6, 25076. [Google Scholar] [CrossRef] [Green Version]
- Ciardiello, C.; Leone, A.; Lanuti, P.; Roca, M.S.; Moccia, T.; Minciacchi, V.R.; Minopoli, M.; Gigantino, V.; De Cecio, R.; Rippa, M.; et al. Large oncosome soverexpressing integrin alpha-V promote prostate canceradhesion and invasion via AKT activation. J. Exp. Clin. Cancer. Res. 2019, 38, 317. [Google Scholar] [CrossRef] [Green Version]
- Kalkat, M.; De Melo, J.; Hickman, K.A.; Lourenco, C.; Redel, C.; Resetca, D.; Tamachi, A.; Tu, W.B.; Penn, L.Z. MYC deregulation in primary human cancers. Genes 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabay, M.; Yulin, L.; Felsher, D.W. MYC Activation Is a Hallmark of Cancer Initiation and Maintenance. Cold Spring Harb. Perspect. Med. 2014, 4, a014241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.; Zhao, X.; Wang, J.; Liu, X.; Yan, Y.; Liu, L.; Cai, H.; Qu, H.; Lu, N.; Sun, Y.; et al. MYC overexpression with its prognostic and clinicopathological significance in breast cancer. Oncotarget 2017, 8, 93998. [Google Scholar] [CrossRef] [PubMed]
- Nikolaos, F.; Triggiani, E.; Teodossiu, G.; Renda, A. C-myc Overexpression in Preneoplastic Conditions and in Colorectal Cancer as Decisional Factor in Medical and Surgical Treatment. Eur. J. Surg. Oncol. 2011, 37, S4. [Google Scholar]
- Attar, N.; Kurdistani, S.K. Exploitation of EP300 and CREBBP Lysine Acetyltransferases by Cancer. Cold Spring Harb. Perspect. Med. 2017, 7, a026534. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Eller, M.S.; Elm, C.; Larocca, C.A.; Ryu, B.; Panova, I.P.; Dancy, B.M.; Bowers, E.M.; Meyers, D.; Lareau, L.; et al. Selective inhibition of p300 HAT blocks cell cycle progression, induces cellular senescence, and inhibits the DNA damage response in melanoma cells. J. Investig. Dermatol. 2015, 133, 2444–2452. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, A.E.; Krazinski, B.E.; Godlewski, J.; Kiewisz, J.; Kwiatkowski, P.; Sliwinska-Jewsiewicka, A.; Kiezun, J.; Sulik, M.; Kmiec, Z. Expression of the EP300, TP53 and BAX genes in colorectal cancer: Correlations with clinicopathological parameters and survival. Oncol. Rep. 2017, 38, 201–210. [Google Scholar] [CrossRef]
- Goel, R.K.; Lukong, E.K. Understanding the cellular roles of Fyn-related kinase (FRK): Implications in cancer biology. Cancer Metastasis Rev. 2016, 35, 179–199. [Google Scholar] [CrossRef]
- Xie, Y.G.; Yu, Y.; Hou, L.K.; Wang, X.; Zhang, B.; Cao, X.C. FYN promotes breast cancer progression through epithelial-mesenchymal transition. Oncol. Rep. 2016, 36, 1000–1006. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, H.F.; Shen, W.; Xu, D.Y.; Ruan, T.Y.; Tao, G.Q.; Lu, P.H. SRPK2 promotes the growth and migration of the colon cancer cells. Gene 2016, 586, 41–47. [Google Scholar] [CrossRef]
- Lee, G.; Zheng, Y.; Cho, S.; Jang, C.; England, C.; Dempsey, J.M.; Yu, Y.; Liu, X.; He, L.; Cavaliere, P.M.; et al. Post-transcriptional Regulation of De Novo Lipogenesis by mTORC1-S6K1-SRPK2 Signaling. Cell 2017, 171, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Greuber, E.K.; Smith-Pearson, P.; Wang, J.; Pendergast, A.M. Role of ABL family kinases in cancer: From leukaemia to solid tumours. Nat. Rev. Cancer 2013, 13, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, M.; Koumoto, Y.; Kanesaki, Y.; Wu, X.; Kido, H. 20S proteasome prevents aggregation of heat-denatured proteins without PA700 regulatory subcomplex like a molecular chaperone. Biomacromolecules 2004, 5, 1465. [Google Scholar] [CrossRef] [PubMed]
- Bruzzoni-Giovanelli, H.; Gonzalez, J.R.; Sigaux, F.; Villoutreix, B.O.; Cayuela, J.M.; Guilhot, J.; Preudhomme, C.; Guilhot, F.; Poyet, J.L.; Rousselot, P. Genetic polymorphisms associated with increased risk of developing chronic myelogenous leukemia. Oncotarget 2015, 6, 36269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.; Li, H.; Zhang, W.; Shao, Y.; Liu, Y.; Guan, H.; Wu, J.; Kang, Y.; Zhao, J.; Yu, Q.; et al. NUDT21 negatively regulates PSMB2 and CXXC5 by alternative polyadenylation and contributes to hepatocellular carcinoma suppression. Oncogene 2018, 37, 4887. [Google Scholar] [CrossRef] [PubMed]
- Polo, A.; Crispo, A.; Cerino, P.; Falzone, L.; Candido, S.; Giudice, A.; De Petro, G.; Ciliberto, G.; Montella, M.; Budillon, A.; et al. Environment and bladdercancer: Molecularanalysis by interaction networks. Oncotarget 2017, 8, 65240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, C.P.; Nakshatri, H. PROGgeneV2: Enhancements on the existing database. BMC Cancer 2014, 14, 970. [Google Scholar] [CrossRef] [Green Version]
- Rocca, C.; Pasqua, T.; Boukhzar, L.; Anouar, Y.; Angelone, T. Progress in the emerging role of selenoproteins in cardiovascular disease: Focus on endoplasmic reticulum-resident selenoproteins. Cell Mol. Life Sci. 2019, 76, 1–17. [Google Scholar] [CrossRef]
- Rebsch, C.M.; Penna, F.J.; Copeland, P.R. Selenoprotein Expression Is Regulated at Multiple Levels in Prostate Cells. Cell. Res. 2006, 16, 940. [Google Scholar] [CrossRef] [Green Version]
- Ekoue, D.N.; Ansong, E.; Liu, L.; Macias, V.; Deaton, R.; Lacher, C.; Picklo, M.; Nonn, L.; Gann, P.H.; Kajdacsy-Balla, A.; et al. Correlations of SELENOF and SELENOP Genotypes With Serum Selenium Levels and Prostate Cancer. Prostate 2018, 78, 279. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Moreno, O.; Boque, N.; Redrado, M.; Milagro, F.; Campion, J.; Endermann, T.; Takahashi, K.; Saito, Y.; Catena, R.; Schomburg, L.; et al. Selenoprotein-P Is Down-Regulated in Prostate Cancer, Which Results in Lack of Protection Against Oxidative Damage. Prostate 2011, 71, 824. [Google Scholar] [CrossRef] [PubMed]
- Naiki, T.; Naiki-Ito, A.; Asamoto, M.; Kawai, N.; Tozawa, K.; Etani, T.; Sato, S.; Suzuki, S.; Shirai, T.; Kohri, K.; et al. GPX2 Overexpression Is Involved in Cell Proliferation and Prognosis of Castration-Resistant Prostate Cancer. Carcinogenesis 2014, 35, 1962. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Zhong, W.; Zhao, R.; Oberley, T.D. Thioredoxin 1 as a subcellular biomarker of redox imbalance in human prostate cancer progression. Free Radic. Biol. Med. 2010, 49, 2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaranayake, G.J.; Troccoli, C.I.; Huynh, M.; Lyles, R.D.Z.; Kage, K.; Win, A.; Lakshmanan, V.; Kwon, D.; Ban, Y.; Chen, S.X.; et al. Thioredoxin-1 protects against androgen receptor-induced redox vulnerability in castration-resistant prostate cancer. Nat. Commun. 2017, 8, 1204. [Google Scholar] [CrossRef]
- Cunningham, D.; You, Z. In Vitro and in Vivo Model Systems Used in Prostate Cancer Research. J. Biol. Methods 2015, 2, e17. [Google Scholar] [CrossRef] [Green Version]
- Sramkoski, R.M.; Pretlow, T.G.; Giaconia, J.M.; Pretlow, T.P.; Schwartz, S.; Sy, M.S.; Marengo, S.R.; Rhim, J.S.; Zhang, D.; Jacobberger, J.W. A new human prostate carcinoma cell line, 22Rv1. In Vitro Cell. Dev. Biol. Anim. 1999, 35, 403. [Google Scholar] [CrossRef]
- van Bokhoven, A.; Varella-Garcia, M.; Korch, C.; Johannes, W.U.; Smith, E.E.; Miller, H.L.; Nordeen, S.K.; Miller, G.J.; Lucia, M.S. Molecular characterization of human prostate carcinoma cell lines. Prostate 2003, 57, 205. [Google Scholar] [CrossRef]
- Horibata, Y.; Elpeleg, O.; Eran, A.; Hirabayashi, Y.; Savitzki, D.; Tal, G.; Mandel, H.; Sugimoto, H. EPT1 (selenoprotein I) is critical for the neural development and maintenance of plasmalogen in humans. J. Lipid. Res. 2018, 59, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Cvetković, B.; Vučić, V.; Cvetković, Z.; Popović, T.; Glibetić, M. Systemic alterations in concentrations and distribution of plasma phospholipids in prostate cancer patients. Med. Oncol. 2012, 29, 809. [Google Scholar] [CrossRef]
- Lee, J.H.; Jang, J.K.; Ko, K.Y.; Jin, Y.; Ham, M.; Kang, H.; Kim, I.Y. Degradation of selenoprotein S and selenoprotein K through PPARγ-mediated ubiquitination is required for adipocyte differentiation. Cell Death Differ. 2019, 26, 1007. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kwon, J.H.; Jeon, Y.H.; Ko, K.Y.; Lee, S.R.; Kim, I.Y. Pro178 and Pro183 of selenoprotein S are essential residues for interaction withp97(VCP) during endoplasmic reticulum-associated degradation. J. Biol. Chem. 2014, 289, 13758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Park, K.J.; Jang, J.K.; Jeon, Y.H.; Ko, K.Y.; Kwon, J.H.; Lee, S.-R.; Kim, I.Y. Selenoprotein S-dependent Selenoprotein K binding top97(VCP) protein is essential for endoplasmic reticulum-associated degradation. J. Biol. Chem. 2015, 290, 29941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhang, Z.; Rozovsky, S. Selenoprotein K form an intermolecular diselenide bond with unusually high redox potential. FEBS Lett. 2014, 588, 3311. [Google Scholar] [CrossRef] [Green Version]
- Ben, S.B.; Peng, B.; Wang, G.C.; Li, C.; Gu, H.F.; Jiang, H.; Meng, X.L.; Lee, B.J.; Chen, C.L. Overexpression of SelenoproteinSelK in BGC-823 Cells Inhibits Cell Adhesion and Migration. Biochemistry (Mosc) 2015, 80, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Méplan, C.; Rohrmann, S.; Steinbrecher, A.; Schomburg, L.; Jansen, E.; Linseisen, J.; Hesketh, J. Polymorphisms in thioredoxinreductase and selenoprotein K genes and selenium status modulate risk of prostate cancer. PLoS ONE 2012, 7, e48709. [Google Scholar] [CrossRef]
- Xu, C.H.; Xiao, L.M.; Zeng, E.M.; Chen, L.-K.; Zheng, S.-Y.; Li, D.-H.; Liu, Y. MicroRNA-181 inhibits the proliferation, drug sensitivity and invasion of human glioma cells by targeting Selenoprotein K (SELK). Am. J. Transl. Res. 2019, 11, 6632. [Google Scholar]
- Sutherland, A.; Kim, D.H.; Relton, C.; Ahn, Y.O.; Hesketh, J. Polymorphisms in the selenoprotein S and 15-kDa selenoprotein genes are associated with altered susceptibility to colorectal cancer. Genes Nutr. 2010, 5, 215. [Google Scholar] [CrossRef] [Green Version]
- Curran, J.E.; Jowett, J.B.; Elliott, K.S.; Gao, Y.; Gluschenko, K.; Wang, J.; AbelAzim, D.M.; Cai, G.; Mahaney, M.C.; Comuzzie, A.G.; et al. Genetic variation in selenoprotein S influences inflammatory response. Nat Genet. 2005, 37, 1234. [Google Scholar] [CrossRef] [Green Version]
- Curcio, C.; Baqui, M.M.; Salvatore, D.; Rihn, B.H.; Mohr, S.; Harney, J.W.; Larsen, P.R.; Bianco, A.C. The human type 2 iodothyroninedeiodinase is a selenoprotein highly expressed in a mesothelioma cell line. J. Biol. Chem. 2001, 276, 30183. [Google Scholar] [CrossRef] [Green Version]
- Hepburn, A.C.; Steele, R.E.; Veeratterapillay, R.; Wilson, L.; Kounatidou, E.E.; Barnard, A.; Berry, P.; Cassidy, J.R.; Moad, M.; El-Sherif, A.; et al. The induction of core pluripotency master regulators in cancers defines poor clinical outcomes and treatment resistance. Oncogene 2019, 38, 4412. [Google Scholar] [CrossRef] [PubMed]
- Rebello, R.J.; Pearson, R.B.; Hannan, R.D.; Furic, L. Therapeutic Approaches Targeting MYC-Driven Prostate Cancer. Genes 2017, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drazic, A.; Miura, H.; Peschek, J.; Le, Y.; Bach, N.C.; Kriehuber, T.; Winter, J. Methionine oxidation activates a transcription factor in response to oxidative stress. Proc. Natl. Acad. Sci. USA 2013, 110, 9493. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Péterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol. Cell. 2013, 51, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.C.; Lee, S.G.; Choo, M.K.; Kim, J.H.; Lee, H.M.; Kim, S.; Fomenko, D.E.; Kim, H.-Y.; Park, J.M.; Gladyshev, V.N. Selenoprotein MsrB1 promotes anti-inflammatory cytokine gene expression in macrophages and controls immune response in vivo. Sci. Rep. 2017, 7, 5119. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Li, H.; Meng, F.; Sun, X.; Feng, X.; Chen, J.; Li, L.; Liu, J. MethionineSulfoxideReductase B1 Regulates Hepatocellular Carcinoma Cell Proliferation and Invasion via the Mitogen-Activated Protein Kinase Pathway and Epithelial-Mesenchymal Transition. Oxid. Med. Cell. Longev. 2018, 2018, 5287971. [Google Scholar] [CrossRef]
- De Luca, A.; Sacchetta, P.; Nieddu, M.; Di Ilio, C.; Favaloro, B. Important roles of multiple Sp1 binding sites and epigenetic modifications in the regulation of the methionine sulfoxidereductase B1 (MsrB1) promoter. BMC Mol. Biol. 2007, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, R.; Liu, Z.; Plattner, R. EnABLing Tumor Growth and Progression: Recent progress in unraveling the functions of ABL kinases in solid tumor cells. Curr. Pharmacol. Rep. 2018, 4, 367. [Google Scholar] [CrossRef]
- Gruber, M.; Ferrone, L.; Puhr, M.; Santer, F.R.; Furlan, T.; Eder, I.E.; Sampson, N.; Schäfer, G.; Handle, F.; Culig, Z. p300 is up-regulated by docetaxel and is a target in chemoresistant prostate cancer. Endocr. Relat. Cancer 2020, 27, 187. [Google Scholar] [CrossRef] [Green Version]
- Posadas, E.M.; Al-Ahmadie, H.; Robinson, V.L.; Jagadeeswaran, R.; Otto, K.; Kasza, K.E.; Tretiakov, M.; Siddiqui, J.; Pienta, K.J.; Stadler, W.M.; et al. FYN is overexpressed in human prostate cancer. BJU Int. 2009, 103, 171. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, Y.J.; Liu, Z.Z.; Wan, S.; Cai, Z.D.; Xie, J.J.; Cai, Z.D.; Song, S.D.; Wan, Y.P.; Hua, W.; Zhong, W.; et al. Enhanced expression of SRPK2 contributes to aggressive progression and metastasis in prostate cancer. Biomed. Pharmacother. 2018, 102, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Snider, J.; Kotlyar, M.; Saraon, P.; Yao, Z.; Jurisica, I.; Stagljar, I. Fundamentals of protein interaction network mapping. Mol. Syst. Biol. 2015, 11, 848. [Google Scholar] [CrossRef] [PubMed]
- Sinisi, A.A.; Chieffi, P.; Pasquali, D.; Kisslinger, A.; Staibano, S.; Bellastella, A.; Tramontano, D. EPN: A novel epithelial cell line derived from human prostate tissue. Cell. Dev. Biol. Anim. 2002, 38, 165. [Google Scholar] [CrossRef]
- Zhao, S.G.; Jackson, W.C.; Kothari, V.; Schipper, M.J.; Erho, N.; Evans, J.R.; Speers, C.; Hamstra, D.A.; Niknafs, Y.S.; Nguyen, P.L.; et al. High-throughput transcriptomic analysis nominates proteasomal genes as age-specific biomarkers and therapeutic targets in prostate cancer. Prostate Cancer Prostatic Dis. 2015, 18, 229. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capone, F.; Polo, A.; Sorice, A.; Budillon, A.; Costantini, S. Integrated Analysis to Study the Relationship between Tumor-Associated Selenoproteins: Focus on Prostate Cancer. Int. J. Mol. Sci. 2020, 21, 6694. https://doi.org/10.3390/ijms21186694
Capone F, Polo A, Sorice A, Budillon A, Costantini S. Integrated Analysis to Study the Relationship between Tumor-Associated Selenoproteins: Focus on Prostate Cancer. International Journal of Molecular Sciences. 2020; 21(18):6694. https://doi.org/10.3390/ijms21186694
Chicago/Turabian StyleCapone, Francesca, Andrea Polo, Angela Sorice, Alfredo Budillon, and Susan Costantini. 2020. "Integrated Analysis to Study the Relationship between Tumor-Associated Selenoproteins: Focus on Prostate Cancer" International Journal of Molecular Sciences 21, no. 18: 6694. https://doi.org/10.3390/ijms21186694
APA StyleCapone, F., Polo, A., Sorice, A., Budillon, A., & Costantini, S. (2020). Integrated Analysis to Study the Relationship between Tumor-Associated Selenoproteins: Focus on Prostate Cancer. International Journal of Molecular Sciences, 21(18), 6694. https://doi.org/10.3390/ijms21186694