Introducing the Amphibious Mudskipper Goby as a Unique Model to Evaluate Neuro/Endocrine Regulation of Behaviors Mediated by Buccal Sensation and Corticosteroids

Abstract
:1. Introduction
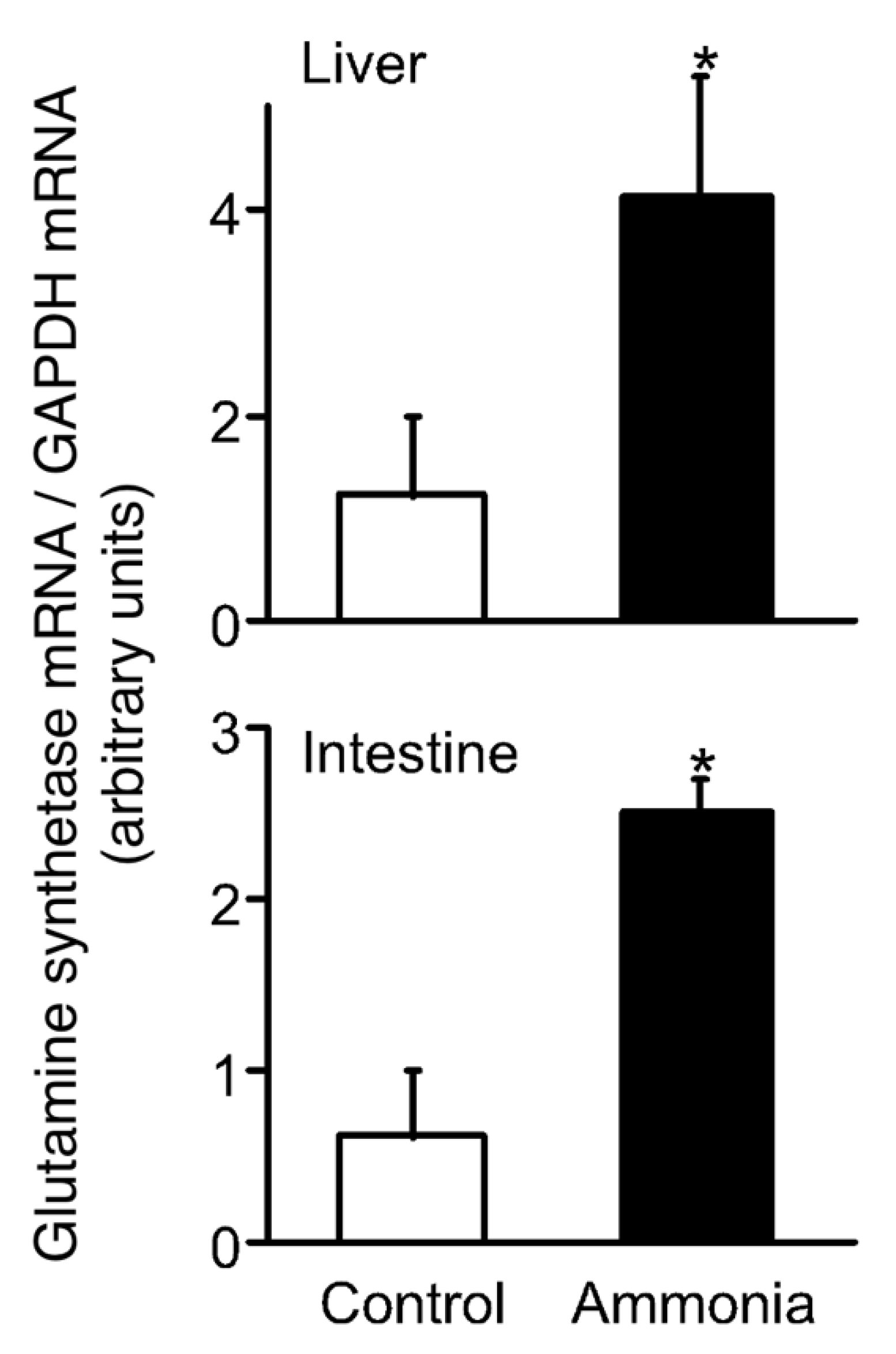
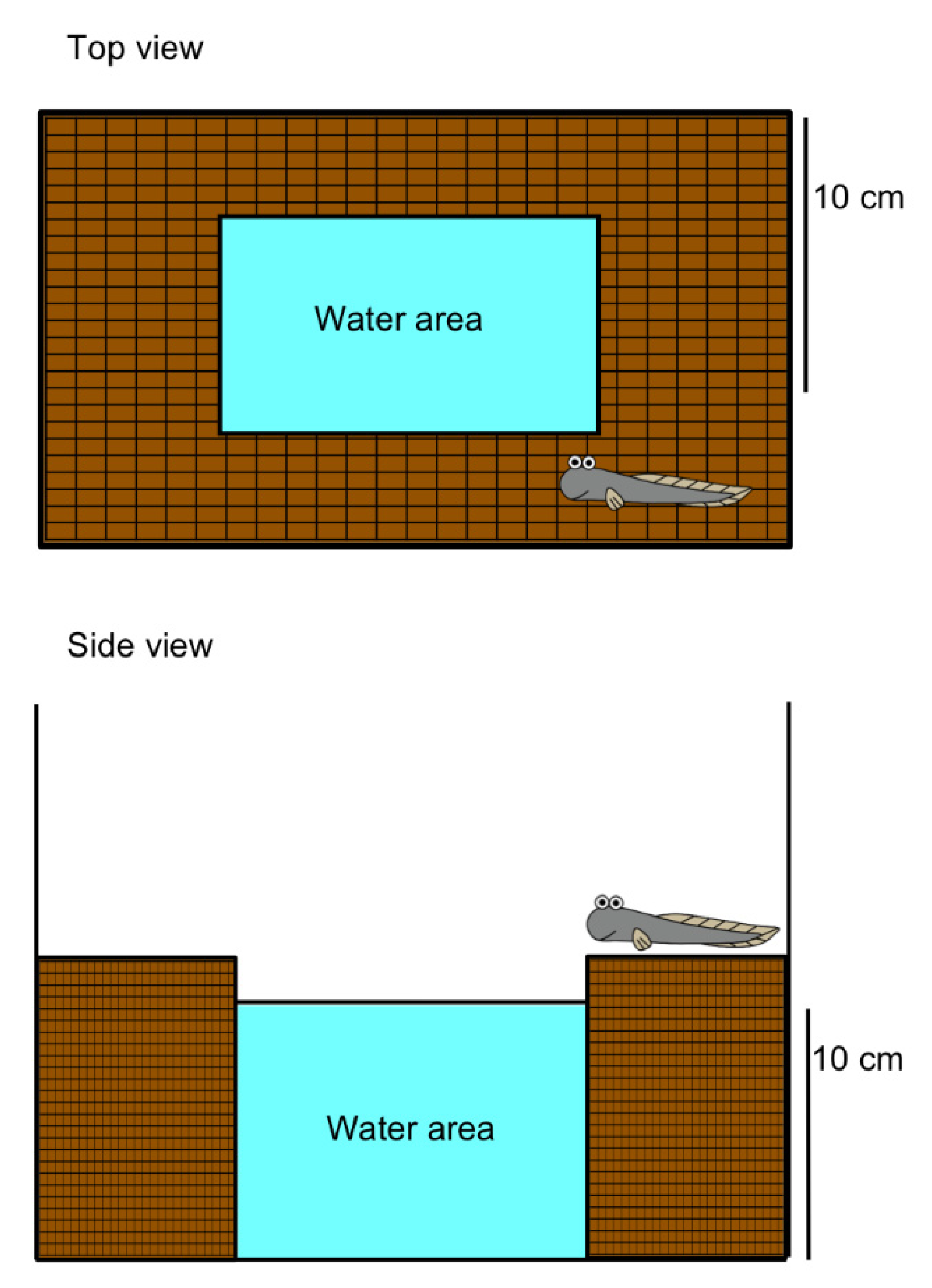
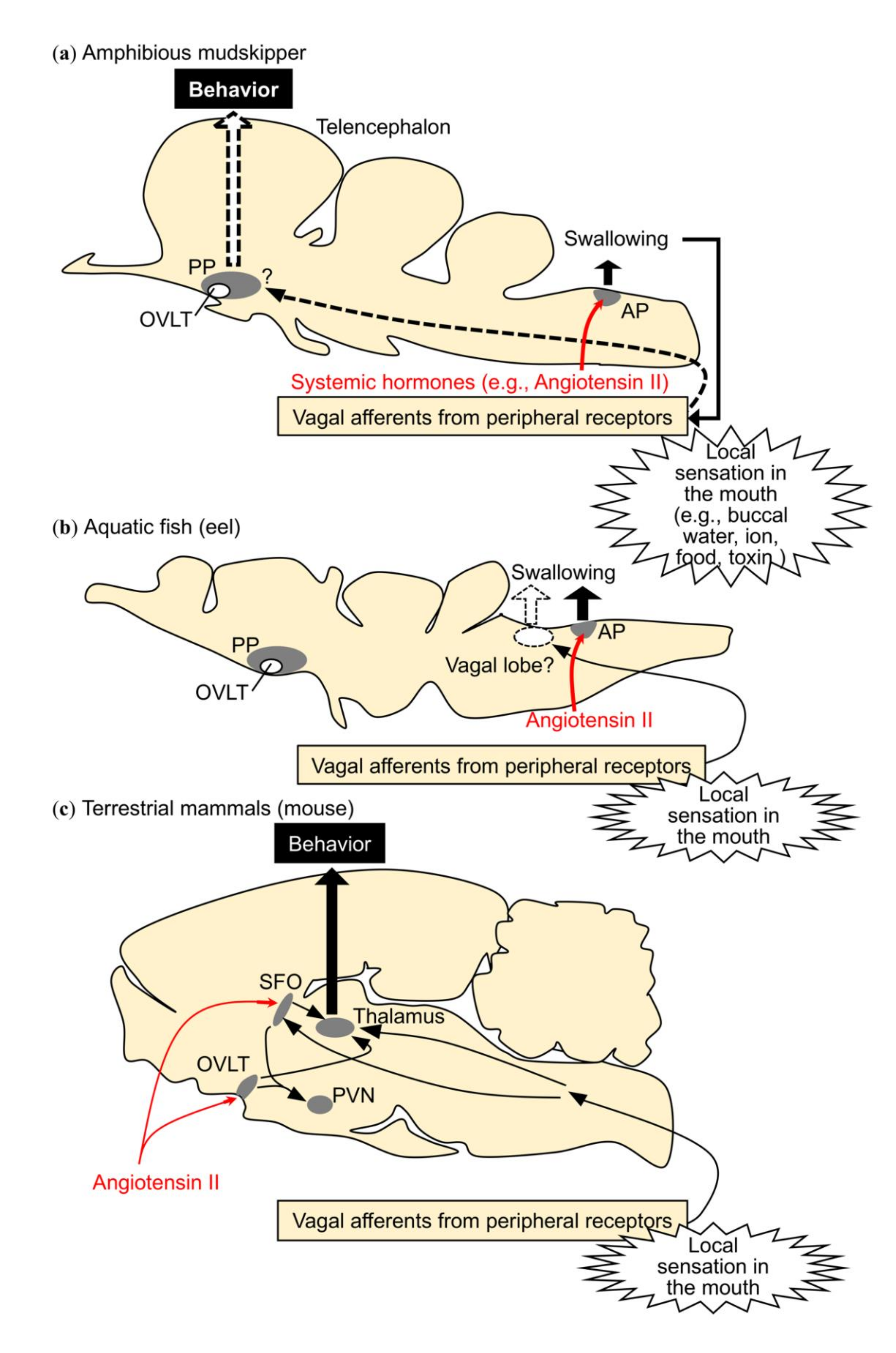
2. Migration to Water Motivated by Local Sensation in the Buccal/Opercular Cavity
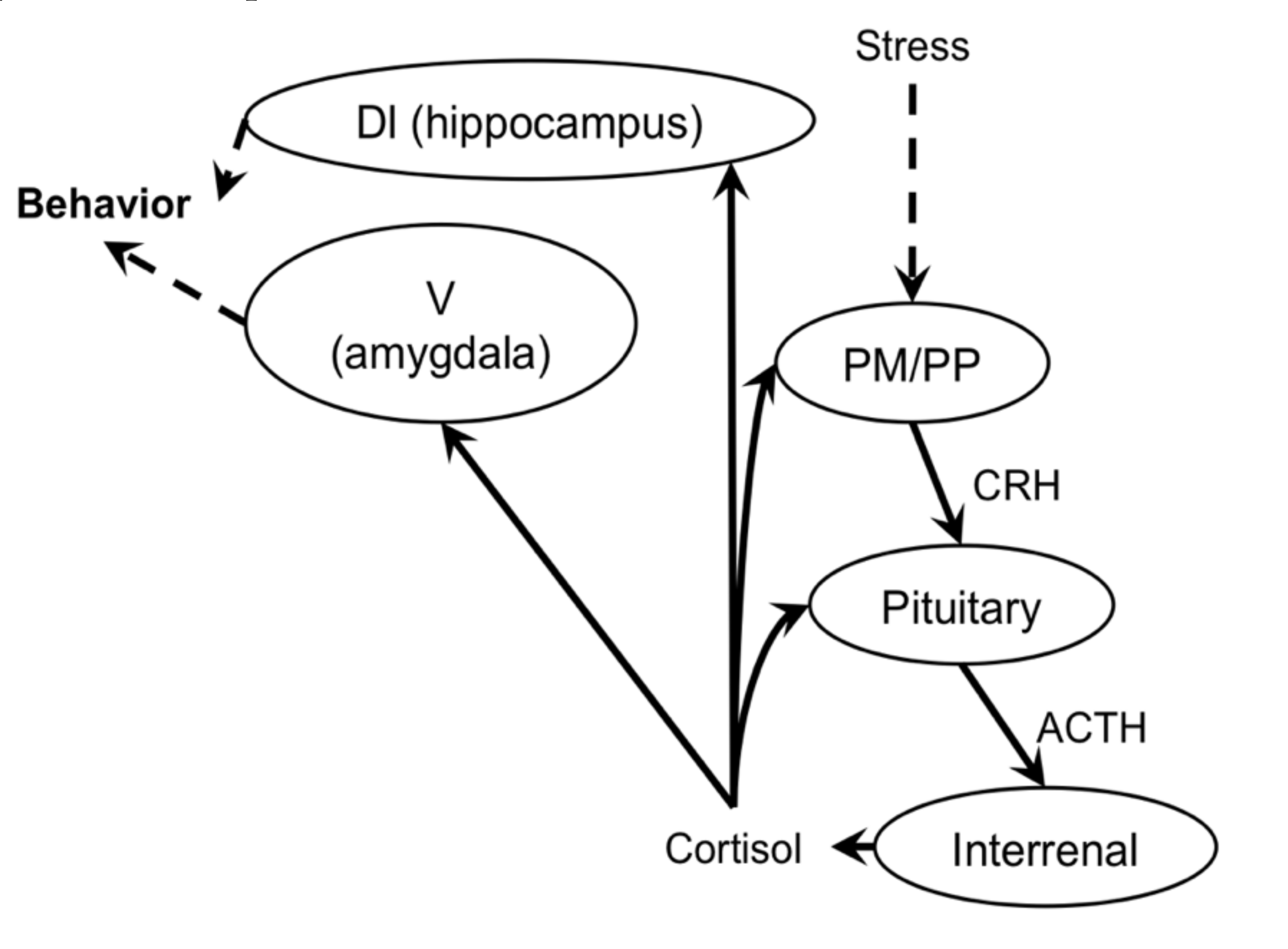
3. Corticosteroid-Regulated Amphibious Behavior
4. Summary and Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ord, T.J.; Cooke, G.M. Repeated evolution of amphibious behavior in fish and its implications for the colonisation of novel environments. Evolution 2016, 70, 1747–1759. [Google Scholar] [CrossRef] [PubMed]
- Barimo, J.F.; Steele, S.L.; Wright, P.A.; Walsh, P.J. Dogmas and controversies in the handling of nitrogenous wastes: Ureotely and ammonia tolerance in early life stages of the gulf toadfish, Opsanus Beta. J. Exp. Biol. 2004, 207, 2011–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ip, A.Y.; Chew, S.F. Ammonia production, excretion, toxicity, and defense in fish: A review. Front. Physiol. 2010, 1, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkie, M.P. Ammonia excretion and urea handling by fish gills: Present understanding and future research challenges. J. Exp. Zool. 2002, 293, 284–301. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Kajimura, M.; Sakamoto, T. Functional ureogenesis in the gobiid fish Mugilogobius abei. J. Exp. Biol. 2000, 203, 3703–3715. [Google Scholar]
- Ip, Y.K.; Chew, S.F.; Wilson, J.M.; Randall, D.J. Defences against ammonia toxicity in tropical air-breathing fishes exposed to high concentrations of environmental ammonia: A review. J. Comp. Physiol. B 2004, 174, 565–575. [Google Scholar] [CrossRef]
- You, X.; Sun, M.; Li, J.; Bian, C.; Chen, J.; Yi, Y.; Yu, H.; Shi, Q. Mudskippers and Their Genetic Adaptations to an Amphibious Lifestyle. Animals 2018, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Ip, Y.K.; Chew, S.F. Air-breathing and excretory nitrogen metabolism in fishes. Acta. Histochem. 2018, 120, 680–690. [Google Scholar] [CrossRef]
- Sayer, M.D.J. Adaptations of amphibious fish for surviving life out of water. Fish Fish. 2005, 6, 186–211. [Google Scholar] [CrossRef]
- Rossi, G.S.; Tunnah, L.; Martin, K.E.; Turko, A.J.; Taylor, D.S.; Currie, S.; Wright, P.A. Mangrove Fishes Rely on Emersion Behavior and Physiological Tolerance to Persist in Sulfidic Environments. Physiol. Biochem. Zool. 2019, 92, 316–325. [Google Scholar] [CrossRef]
- Cochrane, P.V.; Rossi, G.S.; Tunnah, L.; Jonz, M.G.; Wright, P.A. Hydrogen sulphide toxicity and the importance of amphibious behaviour in a mangrove fish inhabiting sulphide-rich habitats. J. Comp. Physiol. B 2019, 189, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Livingston, M.D.; Bhargav, V.V.; Turko, A.J.; Wilson, J.M.; Wright, P.A. Widespread use of emersion and cutaneous ammonia excretion in Aplocheiloid killifishes. Proc. Biol. Sci. 2018, 285, 20181496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blewett, T.A.; Simon, R.A.; Turko, A.J.; Wright, P.A. Copper alters hypoxia sensitivity and the behavioural emersion response in the amphibious fish Kryptolebias Marmoratus. Aquat. Toxicol. 2017, 189, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Randall, D.; Wood, C.; Perry, S.; Bergman, H.; Maloiy, G.; Mommsen, T.; Wright, P. Urea excretion as a strategy for survival in a fish living in a very alkaline environment. Nature 1989, 337, 165–166. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.F.; Ip, Y.K. Excretory nitrogen metabolism and defence against ammonia toxicity in air-breathing fishes. J. Fish Biol. 2014, 84, 603–638. [Google Scholar] [CrossRef]
- Katayama, Y.; Sakamoto, T.; Takanami, K.; Takei, Y. The amphibious mudskipper: A unique model bridging the gap of central actions of osmoregulatory hormones between terrestrial and aquatic vertebrates. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Katayama, Y.; Sakamoto, T.; Saito, K.; Tsuchimochi, H.; Kaiya, H.; Watanabe, T.; Pearson, J.T.; Takei, Y. Drinking by amphibious fish: Convergent evolution of thirst mechanisms during vertebrate terrestrialization. Sci. Rep. UK 2018, 8, 625. [Google Scholar] [CrossRef] [Green Version]
- Clayton, D.A. Mudskippers. Oceanogr. Mar. Biol. 1993, 31, 507–577. [Google Scholar]
- Graham, J.B. Air-Breathing Fishes: Evolution, Diversity, and Adaptation; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Sakamoto, T.; Amano, M.; Hyodo, S.; Moriyama, S.; Takahashi, A.; Kawauchi, H.; Ando, M. Expression of prolactin-releasing peptide and prolactin in the euryhaline mudskippers (Periophthalmus modestus): Prolactin-releasing peptide as a primary regulator of prolactin. J. Mol. Endocrinol. 2005, 34, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Brunt, E.M.; Turko, A.J.; Scott, G.R.; Wright, P.A. Amphibious fish jump better on land after acclimation to a terrestrial environment. J. Exp. Biol. 2016, 219, 3204–3207. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Yasunaga, H.; Yokota, S.; Ando, M. Differential display of skin mRNAs regulated under varying environmental conditions in a mudskipper. J. Comp. Physiol. B 2002, 172, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Ip, Y.K.; Chew, S.F.; Tang, P.C. Evaporation and the turning behavior of the mudskipper, Boleophthalmus boddaerti. Zool. Sci. 1991, 8, 621–623. [Google Scholar]
- Katayama, Y.; Takei, Y.; Kusakabe, M.; Sakamoto, T. Hormonal regulation of thirst in the amphibious ray-finned fish suggests the requirement for terrestrialization during evolution. Sci. Rep. 2019, 9, 16347. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Mori, C.; Minami, S.; Takahashi, H.; Abe, T.; Ojima, D.; Ogoshi, M.; Sakamoto, H. Corticosteroids stimulate the amphibious behavior in mudskipper: Potential role of mineralocorticoid receptors in teleost fish. Physiol. Behav. 2011, 104, 923–928. [Google Scholar] [CrossRef]
- Sakamoto, T.; Nishiyama, Y.; Ikeda, A.; Takahashi, H.; Hyodo, S.; Kagawa, N.; Sakamoto, H. Neurohypophysial Hormones Regulate Amphibious Behaviour in the Mudskipper Goby. PLoS ONE 2015, 10, e0134605. [Google Scholar] [CrossRef] [PubMed]
- Pankhurst, N.W. The endocrinology of stress in fish: An environmental perspective. Gen. Comp. Endocrinol. 2011, 170, 265–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marketon, J.I.; Sternberg, E.M. The glucocorticoid receptor: A revisited target for toxins. Toxins (Basel) 2010, 2, 1357–1380. [Google Scholar] [CrossRef] [Green Version]
- Denton, D.; Shade, R.; Zamarippa, F.; Egan, G.; Blair-West, J.; McKinley, M.; Lancaster, J.; Fox, P. Neuroimaging of genesis and satiation of thirst and an interoceptor-driven theory of origins of primary consciousness. Proc. Natl. Acad. Sci. USA 1999, 96, 5304–5309. [Google Scholar] [CrossRef] [Green Version]
- Gizowski, C.; Bourque, C.W. The neural basis of homeostatic and anticipatory thirst. Nat. Rev. Nephrol. 2018, 14, 11–25. [Google Scholar] [CrossRef]
- Fitzsimons, J.T. Angiotensin, thirst, and sodium appetite. Physiol. Rev. 1998, 78, 583–686. [Google Scholar] [CrossRef]
- Takei, Y. Comparative physiology of body fluid regulation in vertebrates with special reference to thirst regulation. Jpn. J. Physiol. 2000, 50, 171–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinley, M.J.; Johnson, A.K. The physiological regulation of thirst and fluid intake. Physiology 2004, 19, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, K.C. Motivation concepts in behavioral neuroscience. Physiol. Behav. 2004, 81, 179–209. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, C.A.; Lin, Y.-C.; Leib, D.E.; Guo, L.; Huey, E.L.; Daly, G.E.; Chen, Y.; Knight, Z.A. Thirst neurons anticipate the homeostatic consequences of eating and drinking. Nature 2016, 537, 680–684. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, C.A.; Leib, D.E.; Knight, Z.A. Neural circuits underlying thirst and fluid homeostasis. Nat. Rev. Neurosci. 2017, 18, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Michel, K.B.; Heiss, E.; Aerts, P.; Van Wassenbergh, S. A fish that uses its hydrodynamic tongue to feed on land. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20150057. [Google Scholar] [CrossRef] [Green Version]
- Heiss, E.; Aerts, P.; Van Wassenbergh, S. Aquatic–terrestrial transitions of feeding systems in vertebrates: A mechanical perspective. J. Exp. Biol. 2018, 221, jeb154427. [Google Scholar] [CrossRef] [Green Version]
- Fitzsimons, J.; Simons, B.J. The effect on drinking in the rat of intravenous infusion of angiotensin, given alone or in combination with other stimuli of thirst. J. Physiol. 1969, 203, 45–57. [Google Scholar] [CrossRef]
- Oatley, K.; Toates, F. The passage of food through the gut of rats and its uptake of fluid. Psychon. Sci. 1969, 16, 225–226. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T. Some factors regulating water intake by the eel, Anguilla japonica. J. Exp. Biol. 1974, 61, 737–747. [Google Scholar]
- Rankin, J. Drinking in hagfishes and lampreys. Symp. Soc. Exp. Biol. 2002, 54, 1–17. [Google Scholar]
- Takei, Y.; Hiroi, J.; Takahashi, H.; Sakamoto, T. Diverse mechanisms for body fluid regulation in teleost fishes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R778–R792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inokuchi, M.; Breves, J.P.; Moriyama, S.; Watanabe, S.; Kaneko, T.; Lerner, D.T.; Grau, E.G.; Seale, A.P. Prolactin 177, prolactin 188, and extracellular osmolality independently regulate the gene expression of ion transport effectors in gill of Mozambique tilapia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R1251–R1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kültz, D. Physiological mechanisms used by fish to cope with salinity stress. J. Exp. Biol. 2015, 218, 1907–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.-T.; Chou, M.-Y.; Wu, L.-C.; Horng, J.-L.; Lin, L.-Y. Transient receptor potential vanilloid 4 modulates ion balance through the isotocin pathway in zebrafish (Danio rerio). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R751–R759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, A.; Watanabe, S.; Grau, E. Osmoreception: Perspectives on signal transduction and environmental modulation. Gen. Comp. Endocrinol. 2012, 176, 354–360. [Google Scholar] [CrossRef]
- Katayama, Y.; Wong, M.K.-S.; Kusakabe, M.; Fujio, M.; Takahashi, N.; Yaguchi, M.; Tsukada, T. Seawater transfer down-regulates C-type natriuretic peptide-3 expression in prolactin-producing cells of Japanese eel: Negative correlation with plasma chloride concentration. Mol. Cell. Endocrinol. 2020, 507, 110780. [Google Scholar] [CrossRef]
- Mayer-Gostan, N.; Hirano, T. The effects of transecting the IXth and Xth cranial nerves on hydromineral balance in the eel Anguilla anguilla. J. Exp. Biol. 1976, 64, 461–475. [Google Scholar] [PubMed]
- Dickson, I. A thirst-quenching gut–brain signal. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 327. [Google Scholar] [CrossRef]
- Zimmerman, C.A. The Neural Basis of Thirst; UCSF: San Francisco, CA, USA, 2019. [Google Scholar]
- Zocchi, D.; Wennemuth, G.; Oka, Y. The cellular mechanism for water detection in the mammalian taste system. Nat. Neurosci. 2017, 20, 927–933. [Google Scholar] [CrossRef]
- Simpson, J.B.; Routtenberg, A. Subfornical organ: Site of drinking elicitation by angiotensin II. Science (New York, N.Y.) 1973, 181, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Takei, Y. The Renin-Angiotensin System: Comparative Aspects; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1996; Volume 35. [Google Scholar]
- Johnson, A.K.; Buggy, J. Periventricular preoptic-hypothalamus is vital for thirst and normal water economy. Am. J. Physiol. 1978, 234, R122–R129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinley, M.J. The Sensory Circumventricular Organs of the Mammalian Brain: Subfornical Organ, OVLT and Area Postrema; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003; Volume 172. [Google Scholar]
- Augustine, V.; Ebisu, H.; Zhao, Y.; Lee, S.; Ho, B.; Mizuno, G.O.; Tian, L.; Oka, Y. Temporally and Spatially Distinct Thirst Satiation Signals. Neuron 2019, 103, 242–249. [Google Scholar] [CrossRef] [Green Version]
- Augustine, V.; Gokce, S.K.; Lee, S.; Wang, B.; Davidson, T.J.; Reimann, F.; Gribble, F.; Deisseroth, K.; Lois, C.; Oka, Y. Hierarchical neural architecture underlying thirst regulation. Nature 2018, 555, 204. [Google Scholar] [CrossRef] [Green Version]
- Augustine, V.; Lee, S.; Oka, Y. Neural control and modulation of thirst, sodium appetite, and hunger. Cell 2020, 180, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Ye, M.; Zuker, C.S. Thirst driving and suppressing signals encoded by distinct neural populations in the brain. Nature 2015, 520, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Gizowski, C.; Zaelzer, C.; Bourque, C.W. Clock-driven vasopressin neurotransmission mediates anticipatory thirst prior to sleep. Nature 2016, 537, 685–688. [Google Scholar] [CrossRef]
- Allen, W.E.; Chen, M.Z.; Pichamoorthy, N.; Tien, R.H.; Pachitariu, M.; Luo, L.; Deisseroth, K. Thirst regulates motivated behavior through modulation of brainwide neural population dynamics. Science 2019, 364, 6437. [Google Scholar]
- Baker, M.E. Steroid receptors and vertebrate evolution. Mol. Cell. Endocrinol. 2019, 496, 110526. [Google Scholar] [CrossRef] [Green Version]
- Prunet, P.; Sturm, A.; Milla, S. Multiple corticosteroid receptors in fish: From old ideas to new concepts. Gen. Comp. Endocrinol. 2006, 147, 17–23. [Google Scholar] [CrossRef]
- Milla, S.; Terrien, X.; Sturm, A.; Ibrahim, F.; Giton, F.; Fiet, J.; Prunet, P.; Le Gac, F. Plasma 11-deoxycorticosterone (DOC) and mineralocorticoid receptor testicular expression during rainbow trout Oncorhynchus mykiss spermiation: Implication with 17alpha, 20beta-dihydroxyprogesterone on the milt fluidity? Reprod. Biol. Endocrinol. 2008, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolte, E.H.; de Mazon, A.F.; Leon-Koosterziel, K.M.; Jesiak, M.; Bury, N.R.; Sturm, A.; Savelkoul, H.F.; van Kemenade, B.L.V.; Flik, G. Corticosteroid receptors involved in stress regulation in common carp, Cyprinus carpio. J. Endocrinol. 2008, 198, 403–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturm, A.; Bury, N.; Dengreville, L.; Fagart, J.; Flouriot, G.; Rafestin-Oblin, M.; Prunet, P. 11-deoxycorticosterone is a potent agonist of the rainbow trout (Oncorhynchus mykiss) mineralocorticoid receptor. Endocrinology 2005, 146, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.E.; Katsu, Y. 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: Evolution of the mineralocorticoid receptor: Sequence, structure and function. J. Endocrinol. 2017, 234, T1–T16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.E.; Funder, J.W.; Kattoula, S.R. Evolution of hormone selectivity in glucocorticoid and mineralocorticoid receptors. J. Steroid. Biochem. Mol. Biol. 2013, 137, 57–70. [Google Scholar] [CrossRef]
- Baker, M.E.; Katsu, Y. Evolution of the Mineralocorticoid Receptor. Vitam. Horm. 2019, 109, 17–36. [Google Scholar] [CrossRef]
- Baker, M.E.; Chandsawangbhuwana, C.; Ollikainen, N. Structural analysis of the evolution of steroid specificity in the mineralocorticoid and glucocorticoid receptors. BMC Evol. Biol. 2007, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Myers, B.; McKlveen, J.M.; Herman, J.P. Glucocorticoid actions on synapses, circuits, and behavior: Implications for the energetics of stress. Front. Neuroendocrinol. 2014, 35, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Sakamoto, T. The role of ‘mineralocorticoids’ in teleost fish: Relative importance of glucocorticoid signaling in the osmoregulation and ‘central’actions of mineralocorticoid receptor. Gen. Comp. Endocrinol. 2013, 181, 223–228. [Google Scholar] [CrossRef]
- Arterbery, A.S.; Deitcher, D.L.; Bass, A.H. Corticosteroid receptor expression in a teleost fish that displays alternative male reproductive tactics. Gen. Comp. Endocrinol. 2010, 165, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, A.K.; Butler, P.C.; White, R.B.; DeMarco, U.; Pearce, D.; Fernald, R.D. Multiple corticosteroid receptors in a teleost fish: Distinct sequences, expression patterns, and transcriptional activities. Endocrinology 2003, 144, 4226–4236. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Yoshiki, M.; Takahashi, H.; Yoshida, M.; Ogino, Y.; Ikeuchi, T.; Nakamachi, T.; Konno, N.; Matsuda, K.; Sakamoto, H. Principal function of mineralocorticoid signaling suggested by constitutive knockout of the mineralocorticoid receptor in medaka fish. Sci. Rep. 2016, 6, 37991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockrem, J.F. Individual variation in glucocorticoid stress responses in animals. Gen. Comp. Endocrinol. 2013, 181, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Hosono, K.; Yamashita, J.; Kawabata, Y.; Okubo, K. Glucocorticoid receptor exhibits sexually dimorphic expression in the medaka brain. Gen. Comp. Endocrinol. 2015, 223, 47–53. [Google Scholar] [CrossRef]
- Sakamoto, T.; Sakamoto, H. ‘Central’ Actions of Corticosteroid Signaling Suggested by Constitutive Knockout of Corticosteroid Receptors in Small Fish. Nutrients 2019, 11, E611. [Google Scholar] [CrossRef] [Green Version]
- Kolber, B.J.; Wieczorek, L.; Muglia, L.J. Hypothalamic-pituitary-adrenal axis dysregulation and behavioral analysis of mouse mutants with altered glucocorticoid or mineralocorticoid receptor function. Stress 2008, 11, 321–338. [Google Scholar] [CrossRef] [Green Version]
- Teitsma, C.A.; Anglade, I.; Toutirais, G.; Muñoz-cueto, J.a.; Saligaut, D.; Ducouret, B.; Kah, O. Immunohistochemical localization of glucocorticoid receptors in the forebrain of the rainbow trout (Oncorhynchus mykiss). J. Comp. Neurol. 1998, 401, 395–410. [Google Scholar] [CrossRef]
- Kolber, B.J.; Muglia, L.J. Defining brain region-specific glucocorticoid action during stress by conditional gene disruption in mice. Brain Res. 2009, 1293, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Wood, C.M.; McDonald, M.D.; Sundin, L.; Laurent, P.; Walsh, P.J. Pulsatile urea excretion in the gulf toadfish: Mechanisms and controls. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 667–684. [Google Scholar] [CrossRef]
- Geerling, J.C.; Loewy, A.D. Aldosterone in the brain. Am. J. Physiol. Renal. Physiol. 2009, 297, F559–F576. [Google Scholar] [CrossRef]
- Kawata, M.; Nishi, M.; Matsuda, K.; Sakamoto, H.; Kaku, N.; Masugi-Tokita, M.; Fujikawa, K.; Hirahara-Wada, Y.; Takanami, K.; Mori, H. Steroid receptor signalling in the brain-lessons learned from molecular imaging. J. Neuroendocrinol. 2008, 20, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Rozeboom, A.M.; Akil, H.; Seasholtz, A.F. Mineralocorticoid receptor overexpression in forebrain decreases anxiety-like behavior and alters the stress response in mice. Proc. Natl. Acad. Sci. USA 2007, 104, 4688–4693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziv, L.; Muto, A.; Schoonheim, P.J.; Meijsing, S.H.; Strasser, D.; Ingraham, H.A.; Schaaf, M.J.; Yamamoto, K.R.; Baier, H. An affective disorder in zebrafish with mutation of the glucocorticoid receptor. Mol. Psychiatry 2013, 18, 681. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Hyodo, S.; Takagi, W. A possible principal function of corticosteroid signaling that is conserved in vertebrate evolution: Lessons from receptor-knockout small fish. J. Steroid Biochem. Mol. Biol. 2018, 184, 57–61. [Google Scholar] [CrossRef]
- Faught, E.; Vijayan, M.M. Loss of the glucocorticoid receptor in zebrafish improves muscle glucose availability and increases growth. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E1093–E1104. [Google Scholar] [CrossRef]
- Faught, E.; Vijayan, M.M. The mineralocorticoid receptor is essential for stress axis regulation in zebrafish larvae. Sci. Rep. 2018, 8, 18081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatzopoulou, A.; Heijmans, J.P.; Burgerhout, E.; Oskam, N.; Spaink, H.P.; Meijer, A.H.; Schaaf, M.J. Glucocorticoid-Induced Attenuation of the Inflammatory Response in Zebrafish. Endocrinology 2016, 157, 2772–2784. [Google Scholar] [CrossRef]
- Filosa, A.; Barker, A.J.; Dal Maschio, M.; Baier, H. Feeding State Modulates Behavioral Choice and Processing of Prey Stimuli in the Zebrafish Tectum. Neuron 2016, 90, 596–608. [Google Scholar] [CrossRef]
- Mosser, E.A.; Chiu, C.N.; Tamai, T.K.; Hirota, T.; Li, S.N.; Hui, M.; Wang, A.; Singh, C.; Giovanni, A.; Kay, S.A.; et al. Identification of pathways that regulate circadian rhythms using a larval zebrafish small molecule screen. Sci. Rep. UK 2019, 9, 12405. [Google Scholar] [CrossRef]
- Sireeni, J.; Bakker, N.; Jaikumar, G.; Obdam, D.; Slabbekoorn, H.; Tudorache, C.; Schaaf, M. Profound effects of glucocorticoid resistance on anxiety-related behavior in zebrafish adults but not in larvae. Gen. Comp. Endocrinol. 2020, 292, 113461. [Google Scholar] [CrossRef] [PubMed]
- Morbiato, E.; Frigato, E.; Dinarello, A.; Maradonna, F.; Facchinello, N.; Argenton, F.; Carnevali, O.; Dalla Valle, L.; Bertolucci, C. Feeding Entrainment of the Zebrafish Circadian Clock Is Regulated by the Glucocorticoid Receptor. Cells (Basel) 2019, 8, 1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.B.; Schwab, T.L.; Sigafoos, A.N.; Gauerke, J.L.; Krug, R.G., 2nd; Serres, M.R.; Jacobs, D.C.; Cotter, R.P.; Das, B.; Petersen, M.O.; et al. Novel zebrafish behavioral assay to identify modifiers of the rapid, nongenomic stress response. Genes Brain Behav. 2019, 18, e12549. [Google Scholar] [CrossRef] [PubMed]
- Kalamarz-Kubiak, H.; Kleszczyńska, A.; Kulczykowska, E. Cortisol stimulates arginine vasotocin and isotocin release from the hypothalamo-pituitary complex of round goby (Neogobius melanostomus): Probable mechanisms of action. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2015, 323, 616–626. [Google Scholar] [CrossRef]
- Coirini, H.; Marusic, E.T.; De Nicola, A.F.; Rainbow, T.C.; McEwen, B.S. Identification of mineralocorticoid binding sites in rat brain by competition studies and density gradient centrifugation. Neuroendocrinology 1983, 37, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Kiguchi, N.; Sukhtankar, D.D.; Ding, H.; Tanaka, K.-i.; Kishioka, S.; Peters, C.M.; Ko, M.-C. Spinal functions of B-type natriuretic peptide, gastrin-releasing peptide, and their cognate receptors for regulating itch in mice. J. Pharmacol. Exp. Ther. 2016, 356, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Kusube, F.; Tominaga, M.; Kawasaki, H.; Yamakura, F.; Naito, H.; Ogawa, H.; Tomooka, Y.; Takamori, K. Electrophysiological properties of brain-natriuretic peptide-and gastrin-releasing peptide-responsive dorsal horn neurons in spinal itch transmission. Neurosci. Lett. 2016, 627, 51–60. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Wan, L.; Huo, F.-Q.; Barry, D.M.; Li, H.; Zhao, Z.-Q.; Chen, Z.-F. B-type natriuretic peptide is neither itch-specific nor functions upstream of the GRP-GRPR signaling pathway. Mol. Pain 2014, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.K.; Hoon, M.A. The cells and circuitry for itch responses in mice. Science 2013, 340, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.-G.; Chen, Z.-F. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007, 448, 700–703. [Google Scholar] [CrossRef]
- Lozano-Vilano, M.L.; De La Maza-Benignos, M. Diversity and status of Mexican killifishes. J. Fish Biol. 2017, 90, 3–38. [Google Scholar] [CrossRef] [Green Version]
- Turko, A.J.; Wright, P.A. Evolution, ecology and physiology of amphibious killifishes (Cyprinodontiformes). J. Fish Biol. 2015, 87, 815–835. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Lee, B.Y.; Lee, J.H.; Rhee, J.S.; Lee, J.S. Conservation of Hox gene clusters in the self-fertilizing fish Kryptolebias marmoratus (Cyprinodontiformes; Rivulidae). J. Fish Biol. 2016, 88, 1249–1256. [Google Scholar] [CrossRef]
- Lins, L.S.F.; Trojahn, S.; Sockell, A.; Yee, M.C.; Tatarenkov, A.; Bustamante, C.D.; Earley, R.L.; Kelley, J.L. Whole-genome sequencing reveals the extent of heterozygosity in a preferentially self-fertilizing hermaphroditic vertebrate. Genome 2018, 61, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.S.; Lee, J.S. Whole genome data for omics-based research on the self-fertilizing fish Kryptolebias marmoratus. Mar. Pollut. Bull. 2014, 85, 532–541. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Region | MR |
|---|---|
| Stress axis Forebrain pallial area Corticotrophin-releasing hormone cells in preoptic nucleus Adrenocorticotropic hormone-containing cells in pituitary pars distalis | ++ + + |
| Ventral parts of the lateral zone of dorsal telencephalon (presumed hippocampus) Commissural and subcommissural nuclei of telencephalon (presumed amygdala) Several hypothalamic nuclei Glomerulus complex of thalamus Mesencephalic tegmentum Granular layer of optic tectum Cerebellum | ++ ++ ~ ++ + ++ +++ +++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katayama, Y.; Saito, K.; Sakamoto, T. Introducing the Amphibious Mudskipper Goby as a Unique Model to Evaluate Neuro/Endocrine Regulation of Behaviors Mediated by Buccal Sensation and Corticosteroids. Int. J. Mol. Sci. 2020, 21, 6748. https://doi.org/10.3390/ijms21186748
Katayama Y, Saito K, Sakamoto T. Introducing the Amphibious Mudskipper Goby as a Unique Model to Evaluate Neuro/Endocrine Regulation of Behaviors Mediated by Buccal Sensation and Corticosteroids. International Journal of Molecular Sciences. 2020; 21(18):6748. https://doi.org/10.3390/ijms21186748
Chicago/Turabian StyleKatayama, Yukitoshi, Kazuhiro Saito, and Tatsuya Sakamoto. 2020. "Introducing the Amphibious Mudskipper Goby as a Unique Model to Evaluate Neuro/Endocrine Regulation of Behaviors Mediated by Buccal Sensation and Corticosteroids" International Journal of Molecular Sciences 21, no. 18: 6748. https://doi.org/10.3390/ijms21186748
APA StyleKatayama, Y., Saito, K., & Sakamoto, T. (2020). Introducing the Amphibious Mudskipper Goby as a Unique Model to Evaluate Neuro/Endocrine Regulation of Behaviors Mediated by Buccal Sensation and Corticosteroids. International Journal of Molecular Sciences, 21(18), 6748. https://doi.org/10.3390/ijms21186748