Diclofenac Degradation—Enzymes, Genetic Background and Cellular Alterations Triggered in Diclofenac-Metabolizing Strain Pseudomonas moorei KB4






Abstract
:1. Introduction
2. Results
2.1. Degradation and Metabolites Formation of DCF under Monosubstrate and Co-Metabolic Conditions
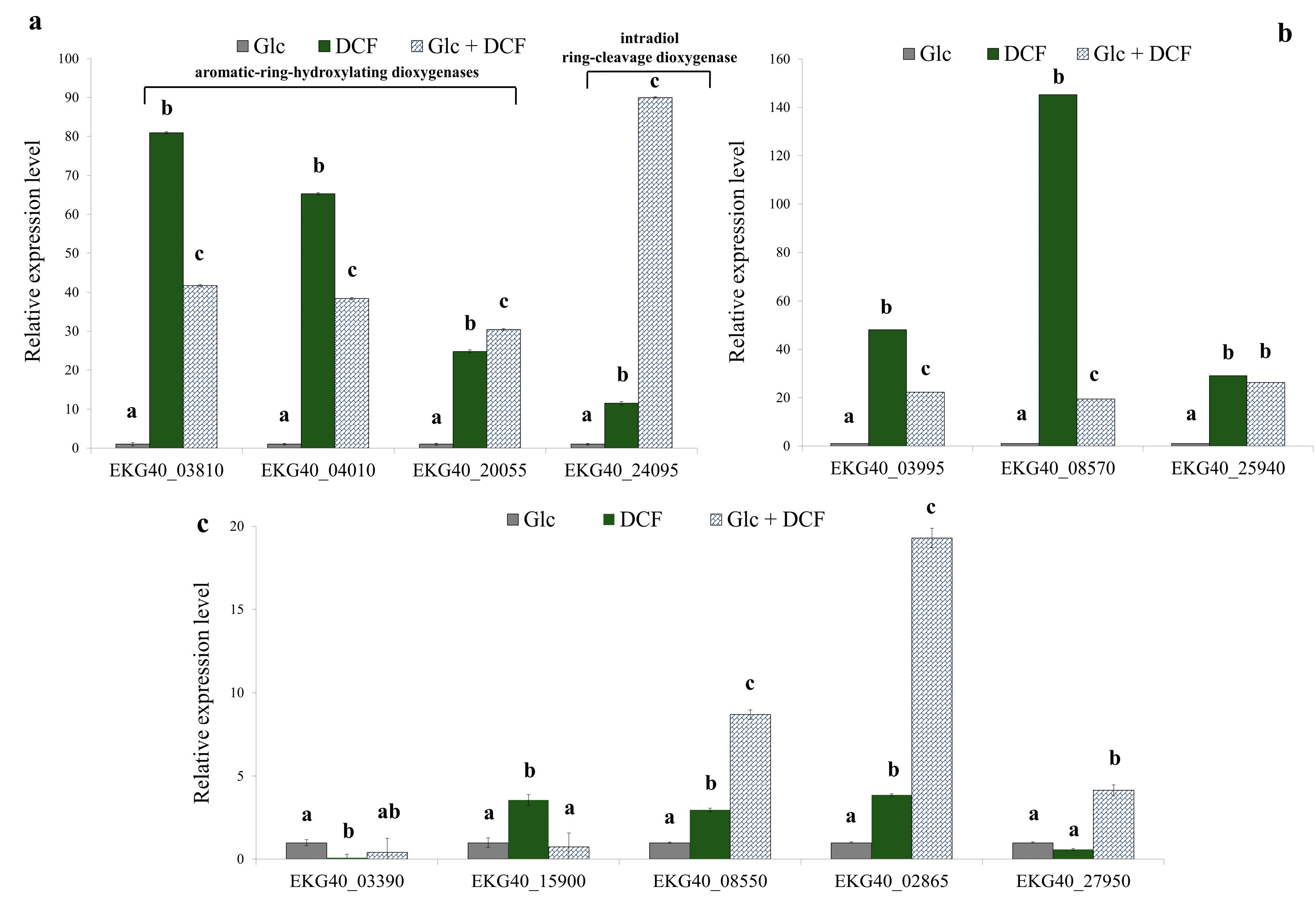
2.2. Analysis of the Specific Activity of Enzymes Involved in DCF Degradation and Its Genetic Background
2.3. Zeta Potential and Cell Wall Hydrophobicity Analysis
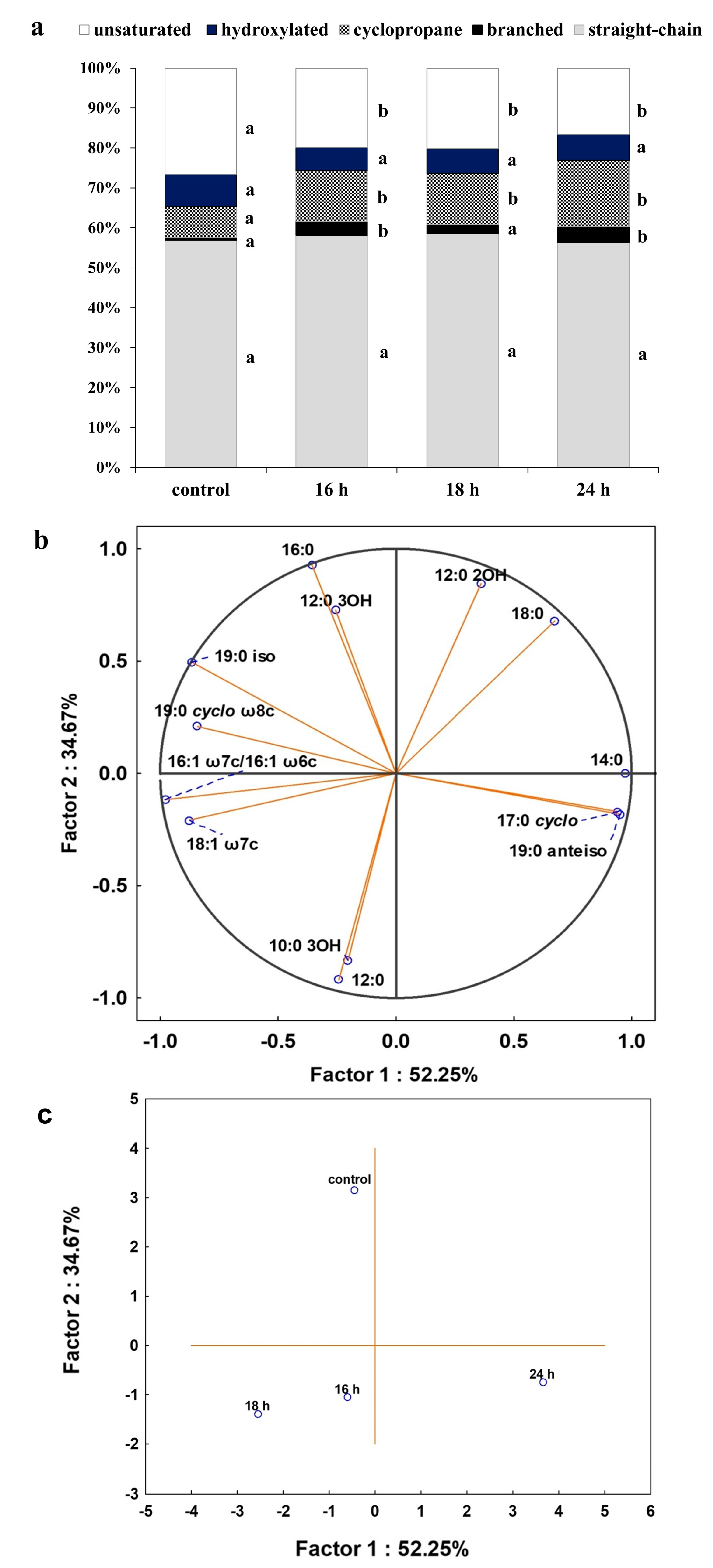
2.4. The Influence of DCF on Fatty Acids Composition and Membrane Permeability
2.5. Oxidative Stress and Cells Injury Induced by DCF
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain
4.2. Degradation Experiments
4.3. Secondary Metabolites Identification
4.4. Analysis of the Specific Activity of Selected Enzymes Involved in DCF Degradation
4.5. Analysis of Expression of Selected Genes Encoding Enzymes Involved in Microbial Degradation of DCF
4.6. Zeta Potential Measuring
4.7. Cell Wall Hydrophobicity Measurements
4.8. Fatty Acid Extraction and Analysis
4.9. Membrane Permeability Analysis
4.10. Cells Injury and Oxidative Stress Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DCF | Diclofenac |
| FAME | Fatty acid methyl ester |
| Glc | Glucose |
| SA | Sodium acetate |
| OD | Optical density |
| SOD | Superoxide dismutase |
| CV | Crystal violet |
| ANOVA | Analysis of variance |
| HPLC | High-Performance Liquid Chromatography |
| UHPLC/MS/MS | Ultra-High Performance Liquid Chromatography coupled with Mass Spectrometry |
| LB | Lysogeny broth |
| MSM | Mineral salts medium |
| mV | Millivolts |
| 4′-OH-DCF | 4′-hydroxylated diclofenac |
| 5-OH-DCF | 5-hydroxylated diclofenac |
| DCF-lactam | Lactam diclofenac |
| DAD | Diode array detector |
| LSD | Least significant differences test |
| CFU | Colony forming unit |
References
- Khan, U.; Nicell, J. Human health relevance of pharmaceutically active compounds in drinking water. AAPS J. 2015, 17, 558–585. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Tyumina, E.A.; Kuzmina, M.V.; Vikhareva, E.V. Features of diclofenac biodegradation by Rhodococcus ruber IEGM 346. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkoom, M.; Lu, G.; Liu, J.; Dong, H.; Yang, H. Bioconcentration, behavioral, and biochemical effects of the non-steroidal anti-inflammatory drug diclofenac in Daphnia magna. Environ. Sci. Pollut. Res. 2019, 26, 5704–5712. [Google Scholar] [CrossRef] [PubMed]
- Sui, Q.; Cao, X.; Lu, S.; Zhao, W.; Qiu, Z.; Yu, G. Occurrence, sources and fate of pharmaceuticals and personal care products in the groundwater: A review. Emerg. Contam. 2015, 1, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; He, J.T.; Su, S.H.; Cui, Y.F.; Huang, D.L.; Wang, G.C. Occurrence, distribution, and attenuation of pharmaceuticals and personal care products in the riverside groundwater of the Beiyun River of Beijing, China. Environ. Sci. Pollut. Res. 2017, 24, 15838–15851. [Google Scholar] [CrossRef]
- Huebner, M.; Weber, E.; Niessner, R.; Boujday, S.; Knopp, D. Rapid analysis of diclofenac in freshwater and wastewater by a monoclonal antibody-based highly sensitive ELISA. Anal. Bioanal. Chem. 2015, 407, 8873–8882. [Google Scholar] [CrossRef] [PubMed]
- Nebot, C.; Falcon, R.; Boyd, K.G.; Gibb, S.W. Introduction of human pharmaceuticals from wastewater treatment plants into the aquatic environment: A rural perspective. Environ. Sci. Pollut. Res. 2015, 22, 10559–10568. [Google Scholar] [CrossRef]
- Osorio, V.; Sanchís, J.; Abad, J.L.; Ginebreda, A.; Farré, M.; Pérez, S.; Barceló, D. Investigating the formation and toxicity of nitrogen transformation products of diclofenac and sulfamethoxazole in wastewater treatment plants. J. Hazard. Mater. 2016, 309, 157–164. [Google Scholar] [CrossRef]
- Vieno, N.; Sillanpää, M. Fate of diclofenac in municipal wastewater treatment plant—A review. Environ. Int. 2014, 69, 28–39. [Google Scholar] [CrossRef]
- Moreira, I.S.; Bessa, V.S.; Murgolo, S.; Piccirillo, C.; Mascolo, G.; Castro, P.M.L. Biodegradation of diclofenac by the bacterial strain Labrys portucalensis F11. Ecotoxicol. Environ. Saf. 2018, 152, 104–113. [Google Scholar] [CrossRef]
- Langenhoff, A.; Inderfurth, N.; Veuskens, T.; Schraa, G.; Blokland, M.; Kujawa-Roeleveld, K.; Rijnaarts, H. Microbial removal of the pharmaceutical compounds ibuprofen and diclofenac from wastewater. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Żur, J.; Piński, A.; Marchlewicz, A.; Hupert-Kocurek, K.; Wojcieszyńska, D.; Guzik, U. Organic micropollutants paracetamol and ibuprofen—Toxicity, biodegradation, and genetic background of their utilization by bacteria. Environ. Sci. Pollut. Res. 2018, 25, 21498–21524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadkaew, N.; Hai, F.I.; McDonald, J.A.; Khan, S.J.; Nghiem, L.D. Removal of trace organics by MBR treatment: The role of molecular properties. Water Res. 2011, 45, 2439–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandclément, C.; Piram, A.; Petit, M.E.; Seyssiecq, I.; Laffont-Schwob, I.; Vanot, G.; Tiliacos, N.; Roche, N.; Doumenq, P. Biological Removal and Fate Assessment of Diclofenac Using Bacillus subtilis and Brevibacillus laterosporus Strains and Ecotoxicological Effects of Diclofenac and 4′-Hydroxy-diclofenac. J. Chem. 2020, 2020. [Google Scholar] [CrossRef]
- Kondakova, T.; Cronan, J.E. Transcriptional regulation of fatty acid cis–trans isomerization in the solvent-tolerant soil bacterium Pseudomonas putida F1. Environ. Microbiol. 2019, 21, 1659–1676. [Google Scholar] [CrossRef]
- Aissaoui, S.; Ouled-Haddar, H.; Sifour, M.; Harrouche, K.; Sghaier, H. Metabolic and co-metabolic transformation of diclofenac by Enterobacter hormachei D15 isolated from activated sludge. Curr. Microbiol. 2016, 74, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Domaradzka, D.; Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. Toxicity od diclofenac and its biotransformation by Raoultella sp. DD4. Pol. J. Environ. Stud. 2016, 25, 2211–2216. [Google Scholar] [CrossRef]
- Bessa, V.S.; Moreira, I.S.; Tiritan, M.E.; Castro, P.M.L. Enrichment of bacterial strains for the biodegradation of diclofenac and carbamazepine from activated sludge. Int. Biodeterior. Biodegrad. 2017, 120, 135–142. [Google Scholar] [CrossRef]
- Stylianou, K.; Hapeshi, E.; Vasquez, M.; Fatta-Kassinos, D.; Vyrides, I. Diclofenac biodegradation by newly isolated Klebsiella sp. KSC: Microbial intermediates and ecotoxicological assessment. J. Environ. Chem. Eng. 2018, 6, 3242–3248. [Google Scholar] [CrossRef]
- Palyzová, A.; Zahradník, J.; Marešová, H.; Řezanka, T. Characterization of the catabolic pathway of diclofenac in Raoultella sp. KDF8. Int. Biodeterior. Biodegrad. 2019, 137, 88–94. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Nghiem, L.D.; Pramanik, B.K.; Oh, S. Cometabolic biotransformation and impacts of the anti-inflammatory drug diclofenac on activated sludge microbial communities. Sci. Total. Environ. 2019, 657, 739–745. [Google Scholar] [CrossRef]
- Navrozidou, E.; Remmas, N.; Melidis, P.; Karpouzas, D.G.; Tsiamis, G.; Ntougias, S. Biodegradation Potential and Diversity of Diclofenac-degrading Microbiota in an Immobilized Cell Biofilter. Processes 2019, 7, 554. [Google Scholar] [CrossRef] [Green Version]
- Żur, J.; Michalska, J.; Piński, A.; Mrozik, A.; Nowak, A. Effects of Low Concentration of Selected Analgesics and Successive Bioaugmentation of the Activated Sludge on Its Activity and Metabolic Diversity. Water 2020, 12, 1133. [Google Scholar] [CrossRef] [Green Version]
- Kraigher, B.; Kosjek, T.; Heath, E.; Kompare, B.; Mandic-Mulec, I. Influence of pharmaceutical residues on the structure of activated sludge bacterial communities in wastewater treatment bioreactors. Water Res. 2008, 42, 4578–4588. [Google Scholar] [CrossRef] [PubMed]
- Bouju, H.; Nastold, P.; Beck, B.; Hollender, J.; Corvini, P.F.X.; Wintgens, T. Elucidation of biotransformation of diclofenac and 4′hydroxydiclofenac during biological wastewater treatment. J. Hazard. Mater. 2016, 301, 443–452. [Google Scholar] [CrossRef]
- Jewell, K.S.; Falås, P.; Wick, A.; Joss, A.; Ternes, T.A. Transformation of diclofenac in hybrid biofilm activated sludge processes. Water Res. 2016, 105, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Islas-Flores, H.; Gómez-Oliván, L.M.; Galar-Martínez, M.; Colín-Cruz, A.; Neri-Cruz, N.; García-Medina, S. Diclofenac-induced oxidative stress in brain, liver, gill and blood of common carp (Cyprinus carpio). Ecotoxicol. Environ. Saf. 2013, 92, 32–38. [Google Scholar] [CrossRef]
- Munari, M.; Matozzo, V.; Gagné, F.; Chemello, G.; Riedl, V.; Finos, L.; Pastore, P.; Badocco, D.; Marin, M.G. Does exposure to reduced pH and diclofenac induce oxidative stress in marine bivalves? A comparative study with the mussel Mytilus galloprovincialis and the clam Ruditapes philippinarum. Environ. Pollut. 2018, 240, 925–937. [Google Scholar] [CrossRef]
- Hoeger, B.; Dietrich, D.R.; Schmid, D.; Hartmann, A.; Hitzfeld, B. Distribution of intraperitoneally injected diclofenac in brown trout (Salmo trutta f. fario). Ecotoxicol. Environ. Saf. 2008, 71, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Kummerová, M.; Zezulka, Š.; Babula, P.; Tříska, J. Possible ecological risk of two pharmaceuticals diclofenac and paracetamol demonstrated on a model plant Lemna minor. J. Hazard. Mater. 2016, 302, 351–361. [Google Scholar] [CrossRef]
- Fu, Q.; Fedrizzi, D.; Kosfeld, V.; Schlechtriem, C.; Ganz, V.; Derrer, S.; Rentsch, D.; Hollender, J. Biotransformation changes bioaccumulation and toxicity of diclofenac in aquatic organisms. Environ. Sci. Technol. 2020, 54, 4400–4408. [Google Scholar] [CrossRef] [PubMed]
- Oaks, J.L.; Gilbert, M.; Virani, M.Z.; Watson, R.T.; Meteyer, C.U.; Rideout, B.A.; Shivaprasad, H.L.; Ahmed, S.; Chaudhry, M.J.I.; Arshad, M.; et al. Diclofenac residues as the cause of vulture population decline in Pakistan. Nature 2004, 427, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Żur, J.; Piński, A.; Michalska, J.; Hupert-Kocurek, K.; Nowak, A.; Wojcieszyńska, D.; Guzik, U. A whole-cell immobilization system on bacterial cellulose for the paracetamol-degrading Pseudomonas moorei KB4 strain. Int. Biodeterior. Biodegrad. 2020, 149, 104919. [Google Scholar] [CrossRef]
- Marchlewicz, A.; Domaradzka, D.; Guzik, U.; Wojcieszyńska, D. Bacillus thuringiensis B1(2015b) is a Gram-Positive Bacteria Able to Degrade Naproxen and Ibuprofen. Water. Air Soil Pollut. 2016, 227, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larcher, S.; Yargeau, V. The effect of ozone on the biodegradation of 17α-ethinylestradiol and sulfamethoxazole by mixed bacterial cultures. Appl. Microbiol. Biotechnol. 2013, 97, 2201–2210. [Google Scholar] [CrossRef]
- Palyzová, A.; Zahradník, J.; Maresová, H.; Sokolová, L.; Kyslíková, E.; Grulich, M.; Štěpánek, V.; Řezanka, T.; Kyslík, P. Potential of the strain Raoultella sp. KDF8 for removal of analgesics. Folia Microbiol. 2017, 63, 273–282. [Google Scholar] [CrossRef]
- Fang, H.H.P.; Liu, H. Effect of pH on hydrogen production from glucose by a mixed culture. Bioresour. Technol. 2002, 82, 87–93. [Google Scholar] [CrossRef]
- Pobudkowska, A.; Domańska, U. Study of pH-dependent drugs solubility in water. Chem. Ind. Chem. Eng. Q. 2014, 20, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Prior, J.E.; Shokati, T.; Christians, U.; Gill, R.T. Identification and characterization of a bacterial cytochrome P450 for the metabolism of diclofenac. Appl. Microbiol. Biotechnol. 2010, 85, 625–633. [Google Scholar] [CrossRef]
- Kosjek, T.; Heath, E.; Pérez, S.; Petrovic, M.; Barceló, D. Metabolism studies of diclofenac and clofibric acid in activated sludge bioreactors using liquid chromatography with quadrupole–time-of-flight mass spectrometry. J. Hydrol. 2009, 372, 109–117. [Google Scholar] [CrossRef]
- Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. Intradiol dioxygenases—The key enzymes in xenobiotics degradation. In Biodegradation of Hazardous and Special Products; Tech Open: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Lei, P.; Zhai, R.; Wen, Z.; Jin, M. Recent advances in lignin valorization with bacterial cultures: Microorganisms, metabolic pathways, and bio-products. Biotechnol. Biofuels 2019, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.; Lin, Z.; Liu, H.; Hu, H. Bacteria-mediated phthalic acid esters degradation and related molecular mechanisms. Appl. Microbiol. Biotechnol. 2018, 102, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Schaab, M.R.; Barney, B.M.; Francisco, W.A. Kinetic and Spectroscopic Studies on the Quercetin 2,3-Dioxygenase from Bacillus subtilis. Biochemistry 2006, 45, 1009–1016. [Google Scholar] [CrossRef]
- Kaczorek, E.; Smułek, W.; Zdarta, A.; Sawczuk, A.; Zgoła-Grześkowiak, A. Influence of saponins on the biodegradation of halogenated phenols. Ecotoxicol. Environ. Saf. 2016, 131, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Yalçin, E.; Çavusoglu, K.; Özen, E. Hydrocarbon degradation by a new Pseudomonas sp., strain RW-II, with polycationic surfactant to modify the cell hydrophobicity. Environ. Technol. 2011, 32, 1743–1747. [Google Scholar] [CrossRef]
- Mohanty, S.; Mukherji, S. Alteration in cell surface properties of Burkholderia spp. during surfactant-aided biodegradation of petroleum hydrocarbons. Appl. Microbiol. Biotechnol. 2012, 94, 193–204. [Google Scholar] [CrossRef]
- Smułek, W.; Zdarta, A.; Guzik, U.; Dudzińska-Bajorek, B.; Kaczorek, E. Rahnella sp. strain EK12: Cell surface properties and diesel oil biodegradation after long-term contact with natural surfactants and diesel oil. Microbiol. Res. 2015, 176, 38–47. [Google Scholar] [CrossRef]
- Murínova, S.; Dercová, K. Bacterial cell membrane adaptation responses on stress caused with the environmental pollutants. Acta Chim. Slovaca 2013, 6, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Marchlewicz, A.; Guzik, U.; Hupert-Kocurek, K.; Nowak, A.; Wilczyńska, S.; Wojcieszyńska, D. Toxicity and biodegradation of ibuprofen by Bacillus thuringiensis B1(2015b). Environ. Sci. Pollut. Res. 2017, 24, 7572–7584. [Google Scholar] [CrossRef] [Green Version]
- Nowak, A.; Mrozik, A. Degradation of 4-chlorophenol and microbial diversity in soil inoculated with single Pseudomonas sp. CF600 and Stenotrophomonas maltophilia KB2. J. Environ. Manag. 2018, 215, 216–229. [Google Scholar] [CrossRef]
- Lindström, F.; Thurnhofer, S.; Vetter, W.; Gröbner, G. Impact on lipid membrane organization by free branched-chain fatty acids. Phys. Chem. Chem. Phys. 2006, 8, 4792–4797. [Google Scholar] [CrossRef] [PubMed]
- Unell, M.; Kabelitz, N.; Jansson, J.K.; Heipieper, H.J. Adaptation of the psychrotroph Arthrobacter chlorophenolicus A6 to growth temperature and the presence of phenols by changes in the anteiso/iso ratio of branched fatty acids. FEMS Microbiol. Lett. 2007, 266, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Górny, D.; Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. Naproxen ecotoxicity and biodegradation by Bacillus thuringiensis B1(2015b) strain. Ecotoxicol. Environ. Saf. 2019, 167, 505–512. [Google Scholar] [CrossRef]
- Palyzová, A.; Marešová, H.; Novák, J.; Zahradník, J.; Řezanka, T. Effect of the anti-inflammatory drug diclofenac on lipid composition of bacterial strain Raoultella sp. KDF8. Folia Microbiol. 2020, 1–11. [Google Scholar] [CrossRef]
- Matejczyk, M.; Ofman, P.; Dąbrowska, K.; Świsłocka, R.; Lewandowski, W. The study of biological activity of transformation products of diclofenac and its interaction with chlorogenic acid. J. Environ. Sci. 2020, 91, 128–141. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, D.; Wang, Z.; Guo, H.; Wang, Y. Oxidative stress response in two representative bacteria exposed to atrazine. FEMS Microbiol. Lett. 2012, 334, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Pompei, R.; Ingianni, A.; Foddis, G.; Di Pietro, G.; Satta, G. Patterns of Phosphatase Activity among Enterobacterial Species. Int. J. Syst. Bacteriol. 1993, 43, 174–178. [Google Scholar] [CrossRef]
- Chouhan, S.; Sharma, S. Sub-chronic diclofenac sodium induced alterations of alkaline phosphatase activity in serum and skeletal muscle of mice. Indian J. Exp. Biol. 2011, 49, 446–454. [Google Scholar]
- Bagnyukova, T.V.; Chahrak, O.I.; Lushchak, V.I. Coordinated response of goldfish antioxidant defenses to environmental stress. Aquat. Toxicol. 2006, 78, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Saucedo-Vence, K.; Dublán-García, O.; López-Martínez, L.X.; Morachis-Valdes, G.; Galar-Martínez, M.; Islas-Flores, H.; Gómez-Oliván, L.M. Short and long-term exposure to diclofenac alter oxidative stress status in common carp Cyprinus carpio. Ecotoxicology 2015, 24, 527–539. [Google Scholar] [CrossRef]
- Gómez-Oliván, L.M.; Galar-Martínez, M.; García-Medina, S.; Valdés-Alanís, A.; Islas-Flores, H.; Neri-Cruz, N. Genotoxic response and oxidative stress induced by diclofenac, ibuprofen and naproxen in Daphnia magna. Drug Chem. Toxicol. 2014, 37, 391–399. [Google Scholar] [CrossRef]
- Doi, H.; Iwasaki, H.; Masubuchi, Y.; Nishigaki, R.; Horie, T. Chemiluminescence associated with the oxidative metabolism of salicylic acid in rat liver microsomes. Chem. Biol. Interact. 2002, 140, 109–119. [Google Scholar] [CrossRef]
- Feito, R.; Valcarcel, Y.; Catala, M. Biomarker assessment of toxicity with miniaturised bioassays: Diclofenac as a case study. Ecotoxicology 2012, 21, 289–296. [Google Scholar] [CrossRef]
- Żur, J.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Marchlewicz, A.; Guzik, U. Paracetamol—Toxicity and microbial utilization. Pseudomonas moorei KB4 as a case study for exploring degradation pathway. Chemosphere 2018, 206, 192–202. [Google Scholar] [CrossRef]
- Marchlewicz, A.; Guzik, U.; Smułek, W.; Wojcieszyńska, D. Exploring the degradation of ibuprofen by Bacillus thuringiensis B1(2015b): The new pathway and factors affecting degradation. Molecules 2017, 22, 1676. [Google Scholar] [CrossRef] [PubMed]
- Wojcieszyńska, D.; Domaradzka, D.; Hupert-Kocurek, K.; Guzik, U. Bacterial degradation of naproxen—undisclosed pollutant in the environment. J. Environ. Manag. 2014, 145, 157–161. [Google Scholar] [CrossRef]
- Wolfe, M.D.; Altier, D.J.; Stubna, A.; Popescu, C.V.; Münck, E.; Lipscomb, J.D. Benzoate 1,2-Dioxygenase from Pseudomonas putida: Single Turnover Kinetics and Regulation of a Two-Component Rieske Dioxygenase. Biochemistry 2002, 41, 9611–9626. [Google Scholar] [CrossRef]
- Sariaslani, F.S.; Harper, D.B.; Higgins, I.J. Microbial degradation of hydrocarbons. Catabolism of 1-phenylalkanes by Nocardia salmonicolor. Biochem. J. 1974, 140, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Livak, J.K.; Schmittgen, T.D. Analysis of Relative Genes Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sze, A.; Erickson, D.; Ren, L.; Li, D. Zeta-potential measurement using the Smoluchowski equation and the slope of the current–time relationship in electroosmotic flow. J. Colloid Interface Sci. 2003, 261, 402–410. [Google Scholar] [CrossRef]
- Pacholak, A.; Smułek, W.; Zdarta, A.; Zgoła-Grześkowiak, A.; Kaczorek, E. Bacterial biodegradation of 4-monohalogenated diphenyl ethers in one-substrate and co-metabolic systems. Catalysts 2018, 8, 472. [Google Scholar] [CrossRef] [Green Version]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachtigall, C.; Weber, C.; Rothenburger, S.; Jaros, D.; Rohm, H. Test parameters and cell chain length of Streptococcus thermophilus affect the microbial adhesion to hydrocarbons assay: A methodical approach. FEMS Microbiol. Lett. 2019, 366, fnz150. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, Y.; Anglade, F.; Mitjavilla, S. Potentiation of iron-induced lipid peroxidation by a series of bipyridyls in relation to their ability to reduce iron. Toxicol. Lett. 1997, 93, 65–71. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Glc | DCF | Glc + DCF |
|---|---|---|---|
| Aromatic monooxygenase | 3.065 ± 0.001 a | 11.660 ± 0.006 b | 110.115 ± 0.07 c |
| Dihydroxylating dioxygenase - naphthalene | 0.000 ± 0.000 a | 12.012 ± 0.001 b | 6.940 ± 0.043 c |
| Salicylate 1,2-dioxygenase | 0.000 ± 0.000 a | 0.207 ± 0.071 b | 0.302 ± 0.021 c |
| Gentisate 1,2-dioxygenase | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Homogentisate 1,2-dioxygenase | 0.000 ± 0.000 a | 1.034 ± 0.002 b | 3.006 ± 0.004 c |
| Catechol 1,2-dioxygenase | 1.000 ± 0.010 a | 2.062 ± 0.051 b | 1.731 ± 0.035 c |
| Catechol 2,3-dioxygenase | 1.850 ± 0.094 a | 0.000 ± 0.000 b | 0.000 ± 0.000 b |
| Protocatechuate 4,5-dioxygenase | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Protocatechuate 3,4-dioxygenase | 55.123 ± 0.010 a | 12.249 ± 0.010 b | 66.695 ± 0.189 c |
| Hydroquinone 1,2 dioxygenase | 0.000 ± 0.000 a | 0.004 ± 0.000 b | 0.000 ± 0.000 a |
| Hydroxyquinol 1,2-dioxygenase | 0.932 ± 0.046 a | 0.108 ± 0.025 b | 0.000 ± 0.000 c |
| Benzoate 1,2-dioxygenase | 0.000 ± 0.000 a | 1.395 ± 0.000 b | 3.033 ± 0.000 c |
| Quercetin 2,3-dioxygenase | 0.000 ± 0.000 a | 0.023 ± 0.001 a | 0.000 ± 0.000 a |
| Peroxidase | 0.017 ± 0.002 a | 0.004 ± 0.001 b | 0.004 ± 0.002 b |
| Laccase | 0.204 ± 0.152 a | 0.221 ± 0.058 a | 0.028 ± 0.011 b |
| Deaminase | 6.172 ± 0.023 a | 16.138 ± 0.045 b | 4.756 ± 0.018 c |
| Enzyme/Parameter | Glc | DCF | Glc + DCF |
|---|---|---|---|
| CAT | 0.30 ± 0.11 a | 1.04 ± 0.06 b | 2.34 ± 0.57 c |
| SOD | 10.51 ± 0.01 a | 80.31 ± 0.00 b | 12.59 ± 0.03 c |
| alkaline phosphatase | 0.02 ± 0.01 a | 2.10 ± 0.03 b | 2.07 ± 0.03 b |
| acid phosphatase | 0.17 ± 0.01 a | 0.04 ± 0.00 b | 0.34 ± 0.02 c |
| LPX | 0.00 ± 0.00 a | 8.67 × 10−6 b | 9.87 × 10−8 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żur, J.; Piński, A.; Wojcieszyńska, D.; Smułek, W.; Guzik, U. Diclofenac Degradation—Enzymes, Genetic Background and Cellular Alterations Triggered in Diclofenac-Metabolizing Strain Pseudomonas moorei KB4. Int. J. Mol. Sci. 2020, 21, 6786. https://doi.org/10.3390/ijms21186786
Żur J, Piński A, Wojcieszyńska D, Smułek W, Guzik U. Diclofenac Degradation—Enzymes, Genetic Background and Cellular Alterations Triggered in Diclofenac-Metabolizing Strain Pseudomonas moorei KB4. International Journal of Molecular Sciences. 2020; 21(18):6786. https://doi.org/10.3390/ijms21186786
Chicago/Turabian StyleŻur, Joanna, Artur Piński, Danuta Wojcieszyńska, Wojciech Smułek, and Urszula Guzik. 2020. "Diclofenac Degradation—Enzymes, Genetic Background and Cellular Alterations Triggered in Diclofenac-Metabolizing Strain Pseudomonas moorei KB4" International Journal of Molecular Sciences 21, no. 18: 6786. https://doi.org/10.3390/ijms21186786
APA StyleŻur, J., Piński, A., Wojcieszyńska, D., Smułek, W., & Guzik, U. (2020). Diclofenac Degradation—Enzymes, Genetic Background and Cellular Alterations Triggered in Diclofenac-Metabolizing Strain Pseudomonas moorei KB4. International Journal of Molecular Sciences, 21(18), 6786. https://doi.org/10.3390/ijms21186786