Exercise Training-Enhanced Lipolytic Potency to Catecholamine Depends on the Time of the Day

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
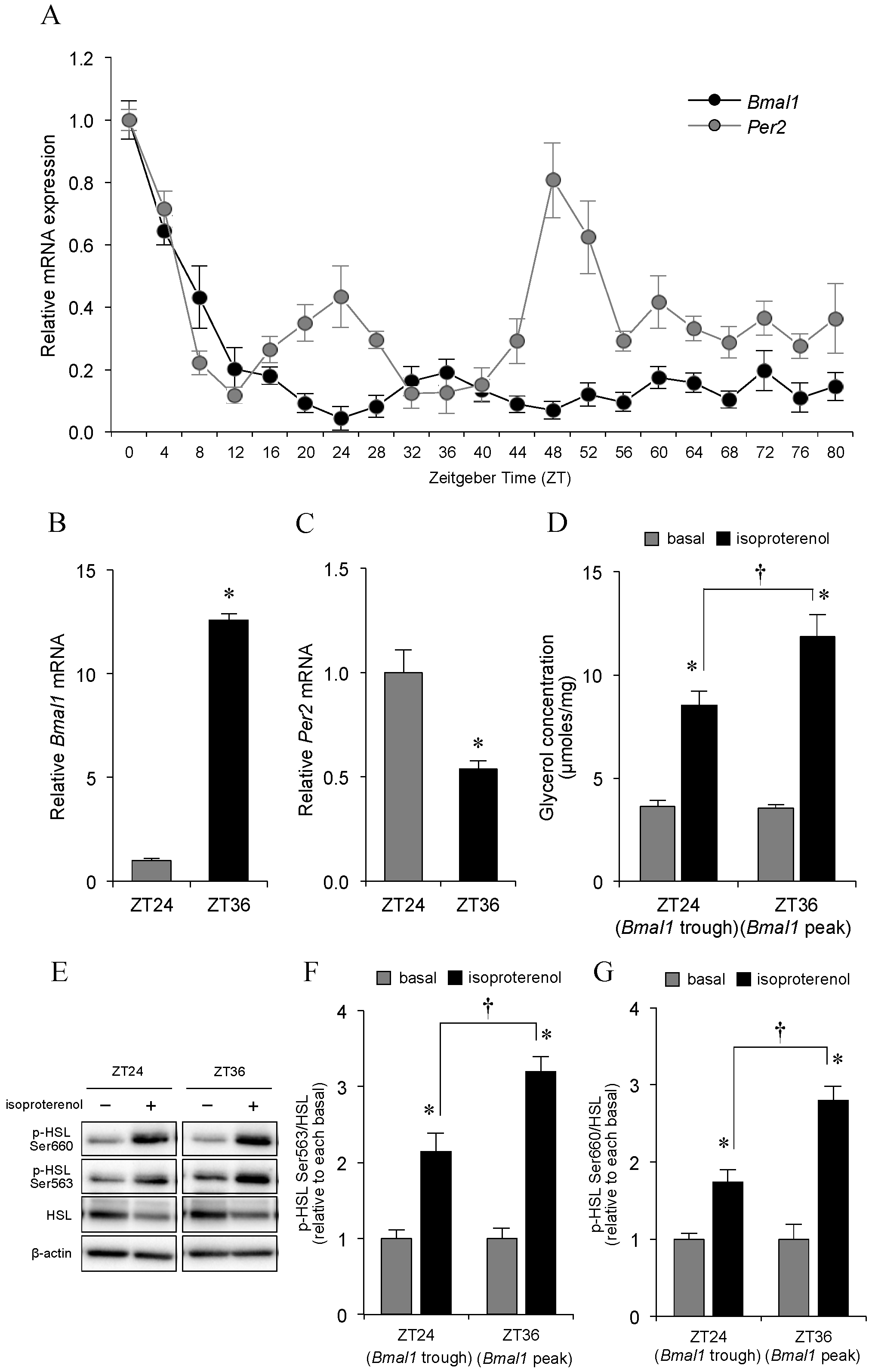
2.1. Effect of Clock Gene mRNA Expression on Adipocyte Lipolysis
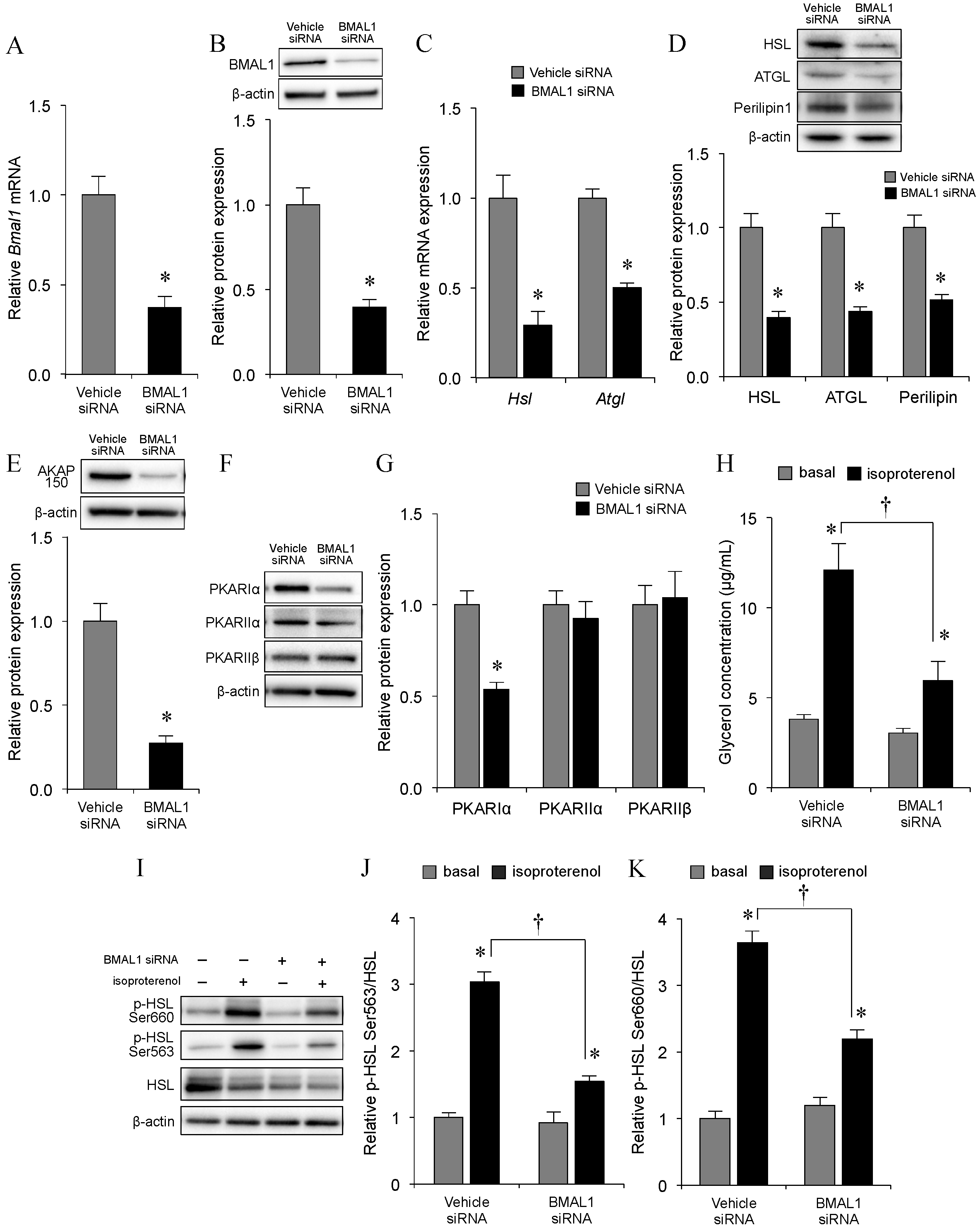
2.2. Effect of Bmal1 Knockdown on Adipocyte Lipolysis
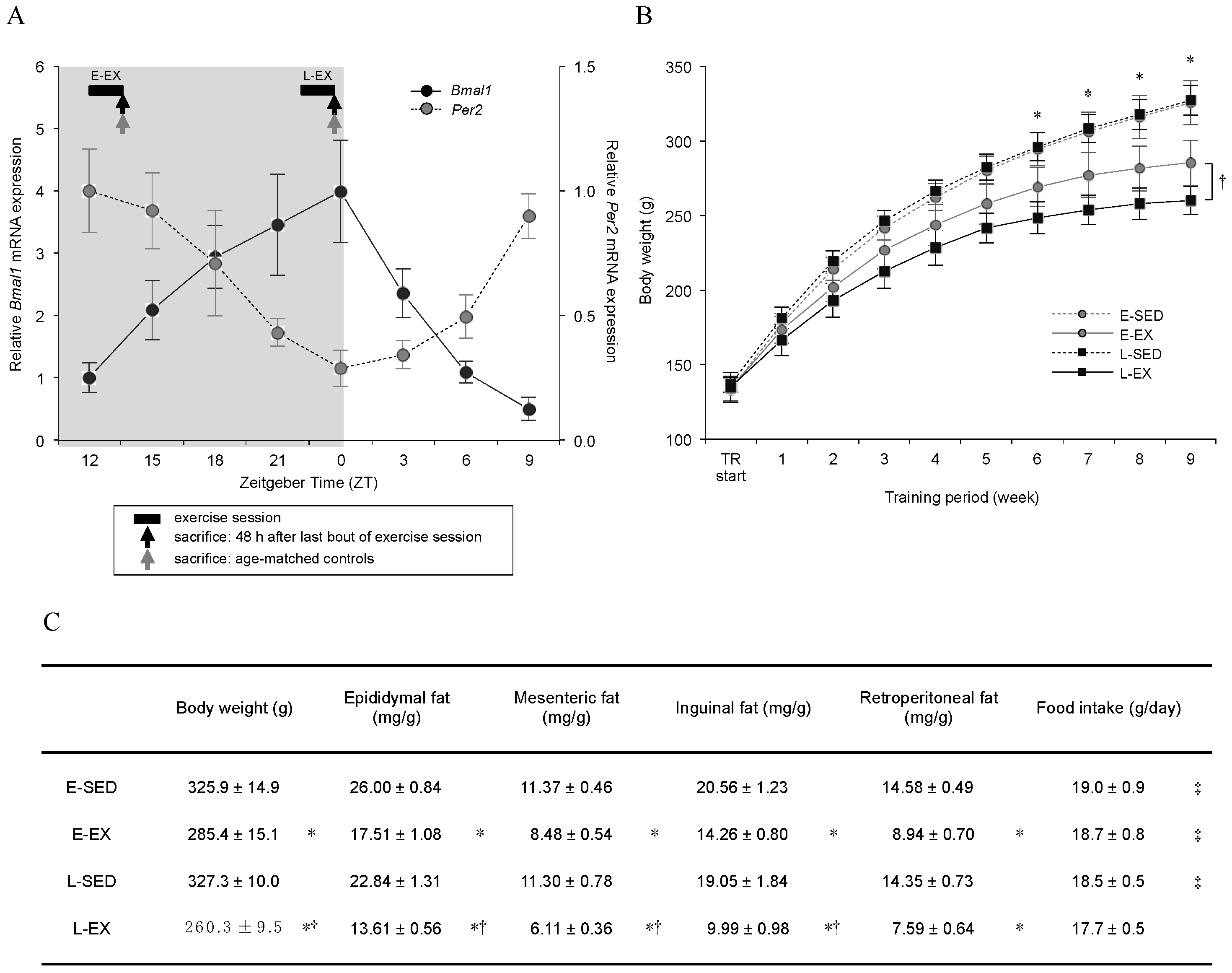
2.3. Circadian Oscillations in Bmal1 and Per2 Expression Patterns in Epididymal Adipose Tissue, and Physical Characteristics of the Rats
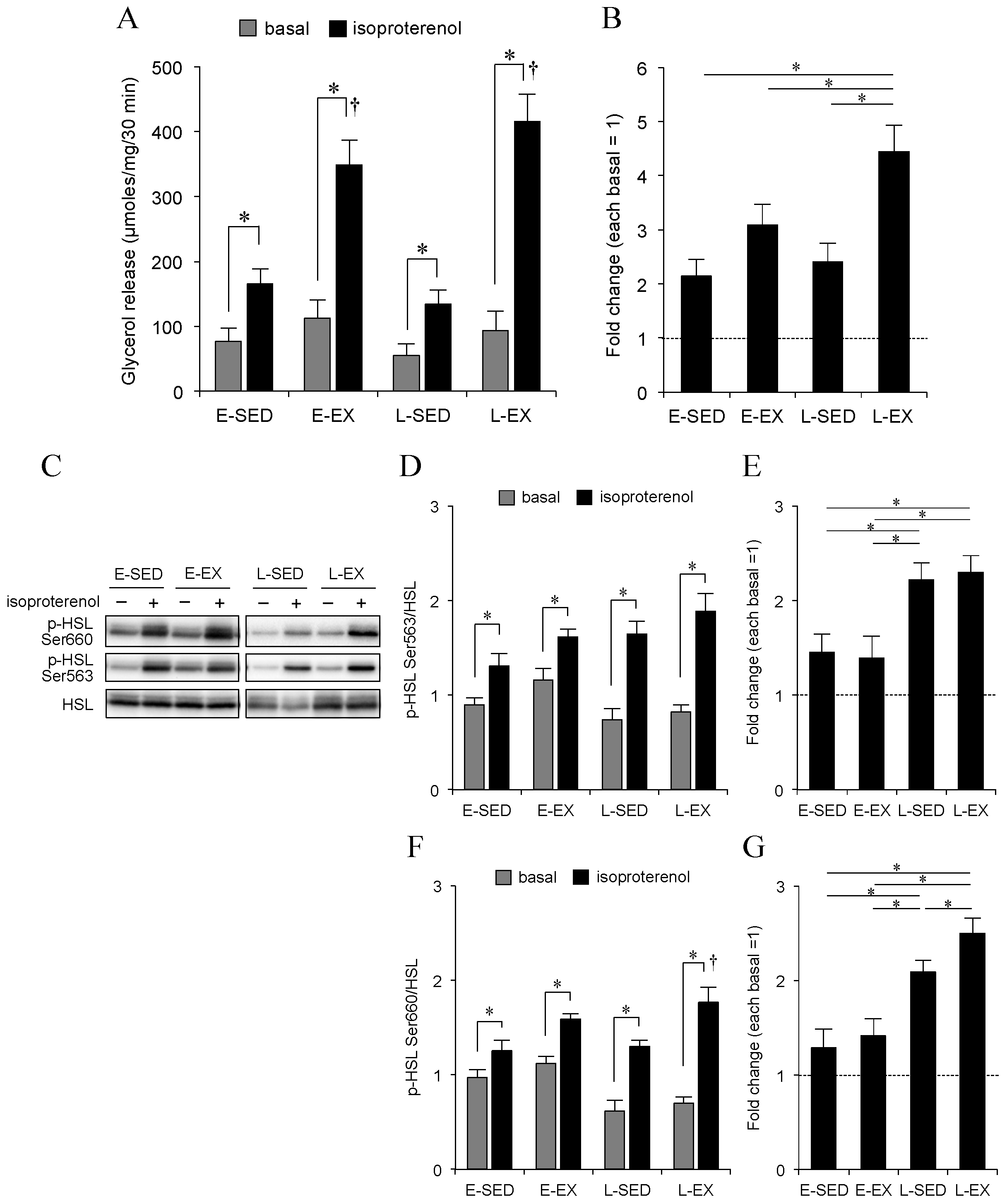
2.4. Adipocyte Lipolysis
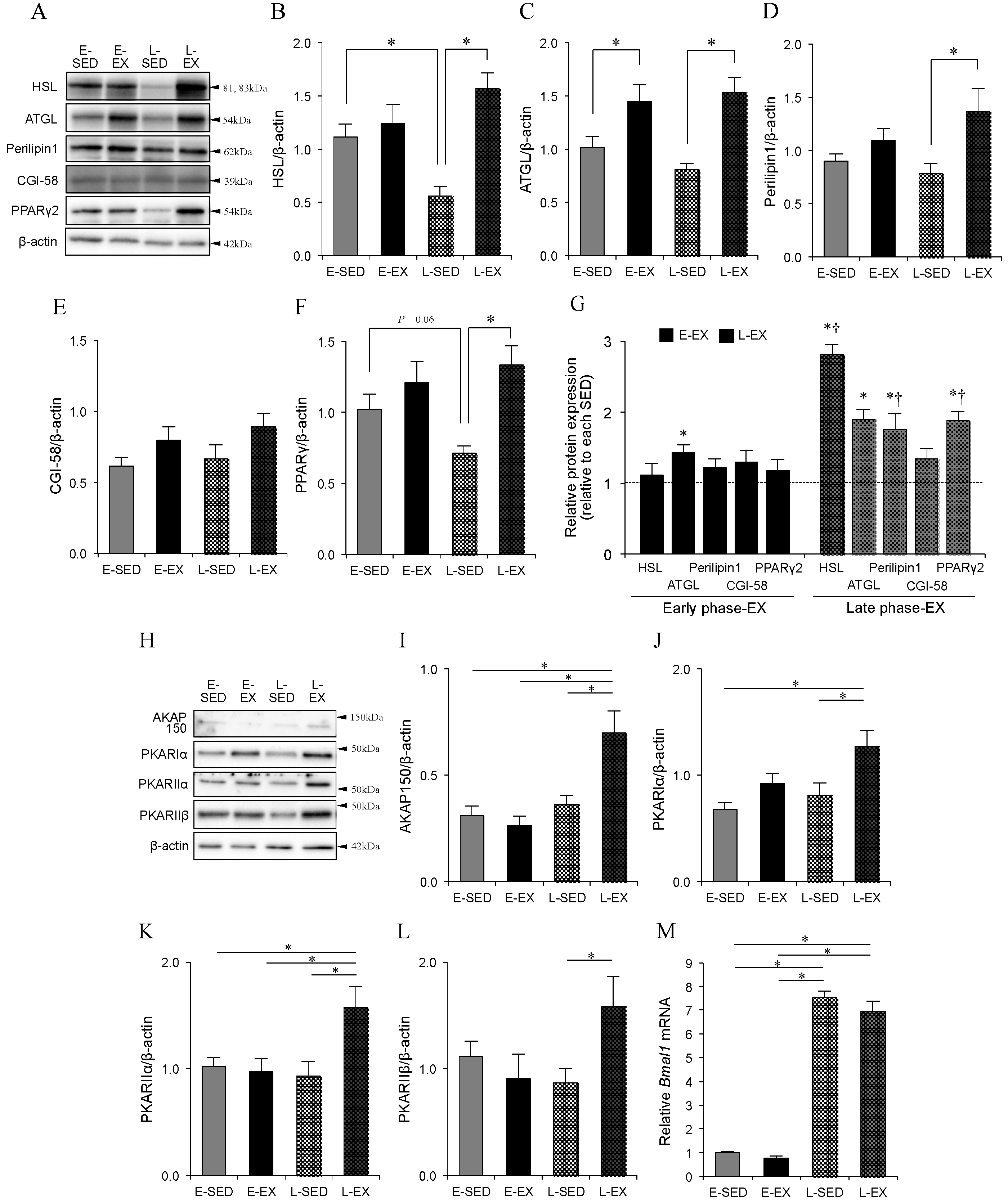
2.5. Changes in Lipolytic Machineries
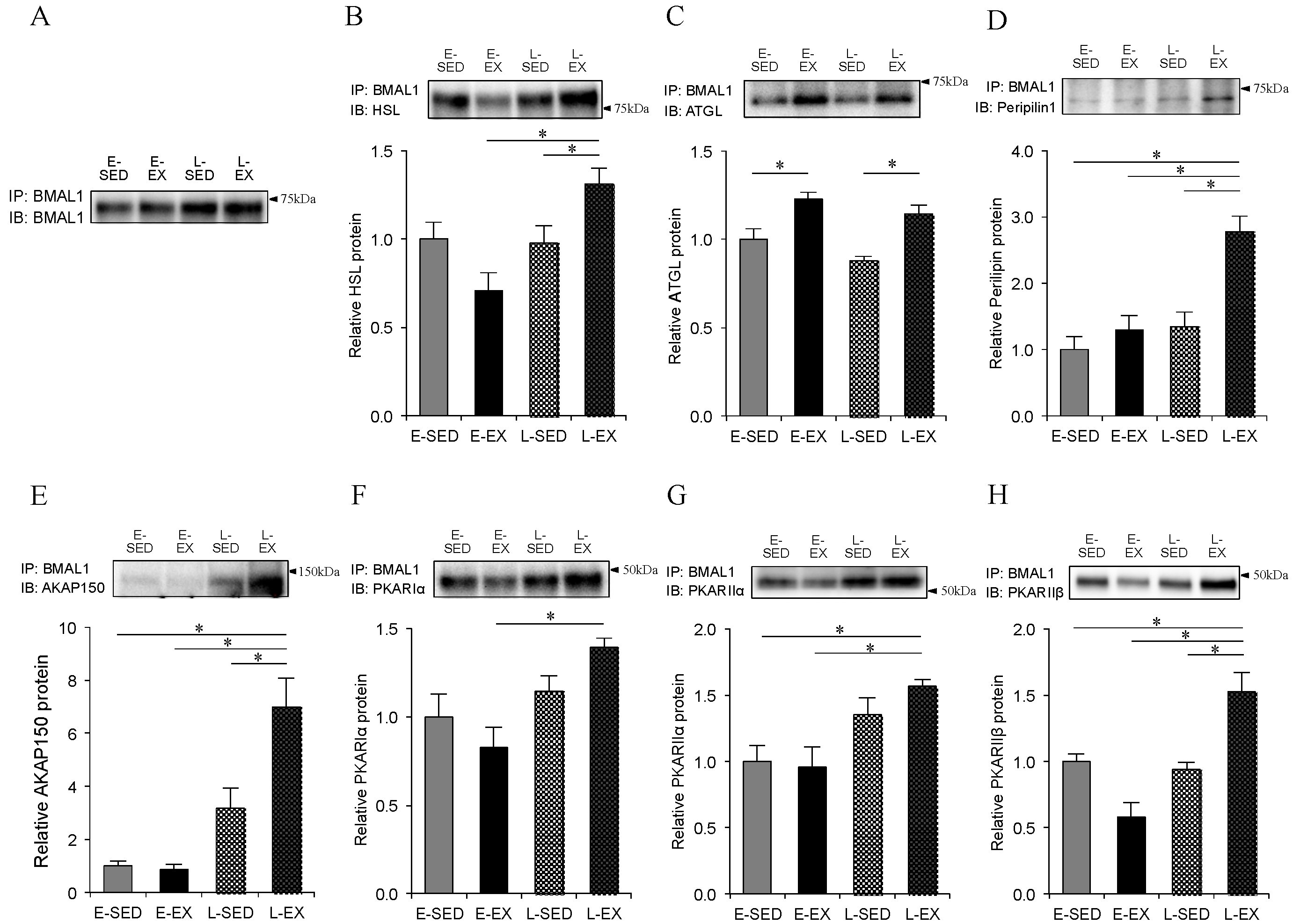
2.6. Association of BMAL1 Protein with Lipolytic Machineries
3. Discussion
4. Materials and Methods
4.1. Bmal1 and Per2 mRNA Circadian Rhythm in Rat Epididymal Adipose Tissue
4.2. Exercise Training Program and Sample Collection Times
4.3. Preparation of Primary Adipocytes
4.4. Lipolysis Assay
4.5. Gene Expression Analysis by Quantitative Real-Time PCR
4.6. Immunoblotting Analysis and Immunoprecipitation
4.7. Cell Culture of 3T3-L1 Mouse Embryo Fibroblasts
4.8. Transfection of 3T3-L1 Cells with Small Interfering RNA (siRNA)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKAP | protein kinase-anchoring protein |
| AMPK | AMP kinase |
| ATGL | adipose triacylglycerol lipase |
| CGI-58 | comparative gene identification-58 |
| DMEM | Dulbecco’s modified Eagle’s medium |
| E-EX | exercise trained at the time point of early part of active phase |
| E-SED | sedentary control for Early-EX |
| HSL | hormone-sensitive lipase |
| L-EX | exercise trained at the time point of late part of active phase |
| L-SED | sedentary control for Late-EX |
| PCR | quantitative real-time polymerase chain reaction |
| PKA | protein kinase |
| SDS | sodium dodecyl sulphate |
| PPARγ | peroxisome proliferator-activated receptor-γ |
| siRNA | small interfering RNA |
References
- Bouassida, A.; Chamari, K.; Zaouali, M.; Feki, Y.; Zbidi, A.; Tabka, Z. Review on leptin and adiponectin responses and adaptations to acute and chronic exercise. Br. J. Sports Med. 2010, 44, 620–630. [Google Scholar] [CrossRef]
- Ogasawara, J.; Izawa, T.; Sakurai, T.; Sakurai, T.; Shirato, K.; Ishibashi, Y.; Ishida, H.; Ohno, H.; Kizaki, T. The molecular mechanism underlying continuous exercise training-induced adaptive changes of lipolysis in white adipose cells. J. Obes. 2015, 2015, 473430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, T.; Ogasawara, J.; Kizaki, T.; Sato, S.; Ishibashi, Y.; Takahashi, M.; Kobayashi, O.; Oh-ishi, S.; Nagasawa, J.; Takahashi, K.; et al. The effects of exercise training on obesity-induced dysregulated expression of adipokines in white adipose tissue. Int. J. Endocrinol. 2013, 2013, 801743. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Karpe, F.; Lafontan, M.; Frayn, K. Physical activity and exercise in the regulation of human adipose tissue physiology. Physiol. Rev. 2012, 92, 157–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, T.; Nicklas, B.J. Effects of exercise on adipokines and the metabolic syndrome. Curr. Diab. Rep. 2008, 8, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, J.; Sakurai, T.; Kizaki, T.; Ishibashi, Y.; Izawa, T.; Sumitani, Y.; Ishida, H.; Radak, Z.; Haga, S.; Ohno, H. Higher levels of ATGL are associated with exercise-induced enhancement of lipolysis in rat epididymal adipocytes. PLoS ONE 2012, 7, e40876. [Google Scholar] [CrossRef]
- Ogasawara, J.; Izawa, T.; Sakurai, T.; Shirato, K.; Ishibashi, Y.; Ohira, Y.; Ishida, H.; Ohno, H.; Kizaki, T. Habitual exercise training acts as a physiological stimulator for constant activation of lipolytic enzymes in rat primary white adipocytes. Biochem. Biophys. Res. Commun. 2015, 464, 348–353. [Google Scholar] [CrossRef]
- Hashimoto, T.; Sato, K.; Iemitsu, M. Exercise-inducible factors to activate lipolysis in adipocytes. J. Appl. Physiol. 2013, 115, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Izawa, T.; Komabayashi, T. Ca2+ and lipolysis in adipocytes from exercise-trained rats. J. Appl. Physiol. 1994, 77, 2618–2624. [Google Scholar] [CrossRef]
- Nomura, S.; Kawanami, H.; Ueda, H.; Kizaki, T.; Ohno, H.; Izawa, T. Possible mechanisms by which adipocyte lipolysis is enhanced in exercise-trained rats. Biochem. Biophys. Res. Commun 2002, 295, 236–242. [Google Scholar] [CrossRef]
- Askew, E.W.; Hecker, A.L. Adipose tissue cell size and lipolysis in the rat: Response to exercise intensity and food restriction. J. Nutr. 1976, 106, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Askew, E.W.; Huston, R.L.; Plopper, C.G.; Hecker, A.L. Adipose tissue cellularity and lipolysis. Response to exercise and cortisol treatment. J. Clin. Investig. 1975, 56, 521–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, J.L.; Fuller, E.O.; Nutter, D.O.; DiGirolamo, M. Influence of moderate exercise on adipocyte metabolism and hormonal responsiveness. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1977, 43, 425–430. [Google Scholar] [CrossRef]
- Bukowiecki, L.; Lupien, J.; Follea, N.; Paradis, A.; Richard, D.; LeBlanc, J. Mechanism of enhanced lipolysis in adipose tissue of exercise-trained rats. Am. J. Physiol. 1980, 239, E422–E429. [Google Scholar] [CrossRef] [PubMed]
- Haemmerle, G.; Lass, A.; Zimmermann, R.; Gorkiewicz, G.; Meyer, C.; Rozman, J.; Heldmaier, G.; Maier, R.; Theussl, C.; Eder, S.; et al. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 2006, 312, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Goldberg, E.B.; Makarova, K.S.; Lin, L.; Brown, W.J.; Jackson, C.L. ATGL has a key role in lipid droplet/adiposome degradation in mammalian cells. EMBO Rep. 2006, 7, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, H.; Perfield, J.W., 2nd; Obin, M.S.; Greenberg, A.S. Adipose triglyceride lipase regulates basal lipolysis and lipid droplet size in adipocytes. J. Cell Biochem. 2008, 105, 1430–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansey, J.T.; Sztalryd, C.; Gruia-Gray, J.; Roush, D.L.; Zee, J.V.; Gavrilova, O.; Reitman, M.L.; Deng, C.-X.; Li, C.; Kimmel, A.R.; et al. Perilipin ablation results in a lean mouse with aberrant adipocyte lipolysis, enhanced leptin production, and resistance to diet-induced obesity. Proc. Natl. Acad. Sci. USA 2001, 98, 6494–6499. [Google Scholar] [CrossRef] [Green Version]
- Zhai, W.; Xu, C.; Ling, Y.; Liu, S.; Deng, J.; Qi, Y.; Londos, C.; Xu, G. Increased lipolysis in adipose tissues is associated with elevation of systemic free fatty acids and insulin resistance in perilipin null mice. Horm. Metab. Res. 2010, 42, 247–253. [Google Scholar] [CrossRef]
- Hashimoto, T.; Segawa, H.; Okuno, M.; Kano, H.; Hamaguchi, H.; Haraguchi, T.; Hiraoka, Y.; Hasui, S.; Yamaguchi, T.; Hirose, F.; et al. Active involvement of micro-lipid droplets and lipid-droplet-associated proteins in hormone-stimulated lipolysis in adipocytes. J. Cell Sci. 2012, 125, 6127–6136. [Google Scholar] [CrossRef] [Green Version]
- Haemmerle, G.; Zimmermann, R.; Hayn, M.; Theussl, C.; Waeg, G.; Wagner, E.; Sattler, W.; Magin, T.M.; Wagner, E.F.; Zechner, R. Hormone-sensitive lipase deficiency in mice causes diglyceride accumulation in adipose tissue, muscle, and testis. J. Biol. Chem. 2002, 277, 4806–4815. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, J.; Klein, S. A novel link between circadian clocks and adipose tissue energy metabolism. Diabetes 2013, 62, 2175–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shostak, A.; Meyer-Kovac, J.; Oster, H. Circadian regulation of lipid mobilization in white adipose tissues. Diabetes 2013, 62, 2195–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shostak, A.; Husse, j.; Oster, H. Circadian regulation of adipose function. Adipocyte 2013, 2, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsalobre, A.; Damiola, F.; Schibler, U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell 1998, 93, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Aoyagi, T.; Shimba, S.; Tezuka, M. Characteristics of circadian gene expressions in mice white adipose tissue and 3T3-L1 adipocytes. J. Health Sci. 2005, 51, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Otway, D.T.; Frost, G.; Johnston, J.D. Circadian rhythmicity in murine pre-adipocyte and adipocyte cells. Chronobiol. Int. 2009, 26, 1340–1354. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 controls lipid metabolism by direct regulation of PPARγ. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Deng, T.; Shan, S.; Li, P.-P.; Shen, Z.-F.; Lu, X.-P.; Cheng, J.; Ning, Z.-Q. Peroxisome proliferator-activated receptor-gamma transcriptionally up-regulates hormone-sensitive lipase via the involvement of specificity protein-1. Endocrinology 2006, 147, 875–884. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Downes, M.; Yu, R.T.; Bookout, A.L.; He, W.; Straume, M.; Mangelsdorf, D.J.; Evans, R. Nuclear receptor expression links the circadian clock to metabolism. Cell 2006, 126, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Petridou, A.; Tsalouhidou, S.; Tsalis, G.; Schulz, T.; Michna, H.; Mougios, V. Long-term exercise increases the DNA binding activity of peroxisome proliferator-activated receptor gamma in rat adipose tissue. Metabolism 2007, 56, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Fraser, I.D.; Cong, M.; Kim, J.; Rollins, E.N.; Daaka, Y.; Lefkowitz, R.J.; Scott, J.D. Assembly of an A kinase-anchoring protein-beta(2)-adrenergic receptor complex facilitates receptor phosphorylation and signaling. Curr. Biol. 2000, 10, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Wang, H.Y.; Malbon, C.C. Protein kinase A regulates AKAP250 (gravin) scaffold binding to the beta2-adrenergic receptor. EMBO J. 2003, 22, 6129–6419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coghlan, V.; Perrino, B.; Howard, M.; Langeberg, L.; Hicks, J.B.; Gallatin, W.M.; Scott, J.D. Association of protein kinase A and protein phosphatase 2B with a common anchoring protein. Science 1995, 267, 108–111. [Google Scholar] [CrossRef]
- Rubin, C.S. A kinase anchor proteins and the intracellular targeting of signals carried by cyclic AMP. Biochim. Biophys. Acta 1994, 1224, 467–479. [Google Scholar]
- McKnight, G.S.; Cummings, D.E.; Amieux, P.S.; Sikorski, M.A.; Brandon, E.P.; Planas, J.V.; Motamed, K.; Idzerda, R.L. Cyclic AMP, PKA, and the physiological regulation of adiposity. Recent Prog. Horm. Res. 1998, 53, 139–161. [Google Scholar]
- Schreyer, S.A.; Cummings, D.E.; McKnight, G.S.; LeBoeuf, R.C. Mutation of the RIIbeta subunit of protein kinase A prevents diet-induced insulin resistance and dyslipidemia in mice. Diatbetes 2001, 50, 2555–2562. [Google Scholar] [CrossRef] [Green Version]
- Planas, J.V.; Cummings, D.E.; Idzerda, R.L.; McKnight, G.S. Mutation of the RIIβ subunit of protein kinase A differentially affects lipolysis but not gene induction in white adipose tissue. J. Biol. Chem. 1999, 274, 36281–36287. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, G.; Bondioni, S.; Alberti, L.; Gilardini, L.; Invitti, C.; Corbetta, S.; Zappa, M.A.; Ferrero, S.; Lania, A.G.; Bosari, S.; et al. Protein Kinase A Regulatory Subunits in Human Adipose Tissue: Decreased R2B Expression and Activity in Adipocytes From Obese Subjects. Diabetes 2009, 58, 620–626. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, H.; Souza, S.C.; Zhang, H.H.; Strissel, K.J.; Christoffolete, M.A.; Kovsan, J.; Rudich, A.; Kraemer, F.B.; Bianco, A.C.; Obin, M.S.; et al. Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms. J. Biol. Chem. 2006, 281, 15837–15844. [Google Scholar] [CrossRef] [Green Version]
- Pidoux, G.; Witczak, O.; Jarnaess, E.; Myrvold, L.; Urlaub, H.; Stokka, A.J.; Kuntziger, T.; Tasken, K. Optic atrophy 1 is an A-kinase anchoring protein on lipid droplets that mediates adrenergic control of lipolysis. EMBO J. 2011, 30, 4371–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, P.K.; Challet, E.; Kalsbeek, A. Circadian rhythms in glucose and lipid metabolism in nocturnal and diurnal mammals. Mol. Cell Endocrinol. 2015, 418, 74–88. [Google Scholar]
- Peckett, A.J.; Wright, D.C.; Riddell, M.C. The effects of glucocorticoids on adipose tissue lipid metabolism. Metabolism 2011, 60, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Benso, M.P.; Rivero-Gutierrez, B.; Lopez-Minguez, J.; Anzola, A.; Diez-Noguera, A.; Madrid, J.A.; Madrid, J.A.; Lujan, J.A.; Martínez-Augustin, O.; Scheer, F.A.J.L.; et al. Human adipose tissue expresses intrinsic circadian rhythm in insulin sensitivity. FASEB J. 2016, 30, 3117–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froy, O.; Garaulet, M. The circadian clock in white and brown adipose tissue: Mechanistic, endocrine, and clinical aspects. Endocr. Rev. 2018, 39, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Dollet, L.; Zierath, J.R. Interplay between diet, exercise and the molecular circadian clock in orchestrating metabolic adaptations of adipose tissue. J. Physiol. 2019, 597, 1439–1450. [Google Scholar] [CrossRef] [Green Version]
- Dalbram, E.; Basse, A.L.; Zierath, J.R.; Treebak, J.T. Voluntary wheel running in the late dark phase ameliorates diet-induced obesity in mice without altering insulin action. J. Appl. Physiol. 2019, 126, 993–1005. [Google Scholar] [CrossRef]
- Sato, S.; Basse, A.L.; Schönke, M.; Chen, S.; Samad, M.; Altıntaş, A.; Laker, R.C.; Dalbram, E.; Barrès, R.; Baldi, P.; et al. Time of exercise specifies the impact on muscle metabolic pathways and systemic energy homeostasis. Cell Metab. 2019, 30, 92–110. [Google Scholar] [CrossRef]
- Ezagouri, S.; Zwighaft, Z.; Sobel, J.; Baillieul, S.; Doutreleau, S.; Ladeuix, B.; Golik, M.; Verges, S.; Asher, G. Physiological and molecular dissection of daily variance in exercise capacity. Cell Metab. 2019, 30, 78–91. [Google Scholar] [CrossRef]
- Gleeson, T.T.; Baldwin, K.M. Cardiovascular response to treadmill exercise in untrained rats. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 50, 1206–1211. [Google Scholar] [CrossRef]
- Rodrigues, B.; Figueroa, D.M.; Mostarda, C.T.; Heeren, M.V.; Irigoyen, M.C.; De Angelis, K. Maximal exercise test is a useful method for physical capacity and oxygen consumption determination in streptozotocin-diabetic rats. Cardiovasc Diabetology 2007, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisløff, U.; Helgerud, J.; Kemi, O.J.; Ellingsen, O. Intensity-controlled treadmill running in rats: Vo(2 max) and cardiac hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1301–H310. [Google Scholar] [CrossRef] [Green Version]
- Oh-ishi, S.; Kizaki, T.; Nagasawa, J.; Izawa, T.; Komabayashi, T.; Nagata, N.; Suzuki, K.; Taniguchi, N.; Ohno, H. Effects of endurance training on superoxide dismutase activity, content and mRNA expression in rat muscle. Clin. Exp. Pharmacol. Physiol. 1997, 24, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Tanaka, G.; Masuda, S.; Ogasawara, J.; Sakurai, T.; Kizaki, T.; Ohno, H.; Izawa, T. Melatonin promotes adipogenesis and mitochondrial biogenesis in 3T3-L1 preadipocytes. J. Pineal Res. 2015, 59, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Carpene, C.; Galitzky, J.; Fontana, E.; Atgie, C.; Lafontan, M.; Berlan, M. Selective activation of beta3-adrenoceptors by octopamine: Comparative studies in mammalian fat cells. Naunyn Schmiedeberg’s Arch. Pharmacol. 1999, 359, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Germack, R.; Starzec, A.B.; Vassy, R.; Perret, G.Y. β-Adrenoceptor subtype expression and function in rat white adipocytes. Br. J. Pharmacol. 1997, 120, 201–210. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, H.; Ogasawara, J.; Takakura, H.; Shirato, K.; Sakurai, T.; Kizaki, T.; Izawa, T. Exercise Training-Enhanced Lipolytic Potency to Catecholamine Depends on the Time of the Day. Int. J. Mol. Sci. 2020, 21, 6920. https://doi.org/10.3390/ijms21186920
Kato H, Ogasawara J, Takakura H, Shirato K, Sakurai T, Kizaki T, Izawa T. Exercise Training-Enhanced Lipolytic Potency to Catecholamine Depends on the Time of the Day. International Journal of Molecular Sciences. 2020; 21(18):6920. https://doi.org/10.3390/ijms21186920
Chicago/Turabian StyleKato, Hisashi, Junetsu Ogasawara, Hisashi Takakura, Ken Shirato, Takuya Sakurai, Takako Kizaki, and Tetsuya Izawa. 2020. "Exercise Training-Enhanced Lipolytic Potency to Catecholamine Depends on the Time of the Day" International Journal of Molecular Sciences 21, no. 18: 6920. https://doi.org/10.3390/ijms21186920
APA StyleKato, H., Ogasawara, J., Takakura, H., Shirato, K., Sakurai, T., Kizaki, T., & Izawa, T. (2020). Exercise Training-Enhanced Lipolytic Potency to Catecholamine Depends on the Time of the Day. International Journal of Molecular Sciences, 21(18), 6920. https://doi.org/10.3390/ijms21186920