In Vitro Inhibition of Phosphodiesterase 3B (PDE 3B) by Anthocyanin-Rich Fruit Juice Extracts and Selected Anthocyanins

 and
and
Abstract
:1. Introduction
2. Results
2.1. HPLC-ESI-MSn Analysis
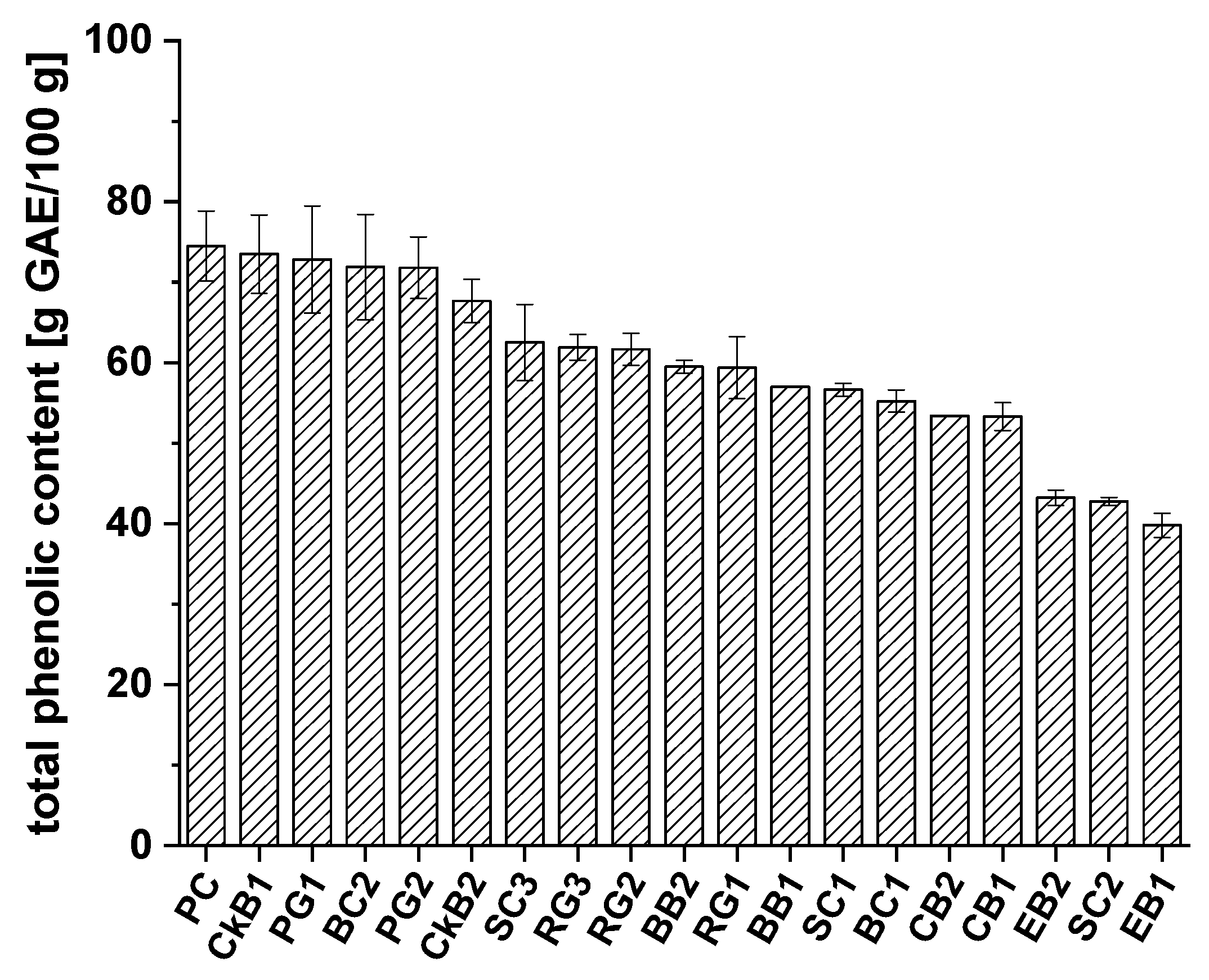
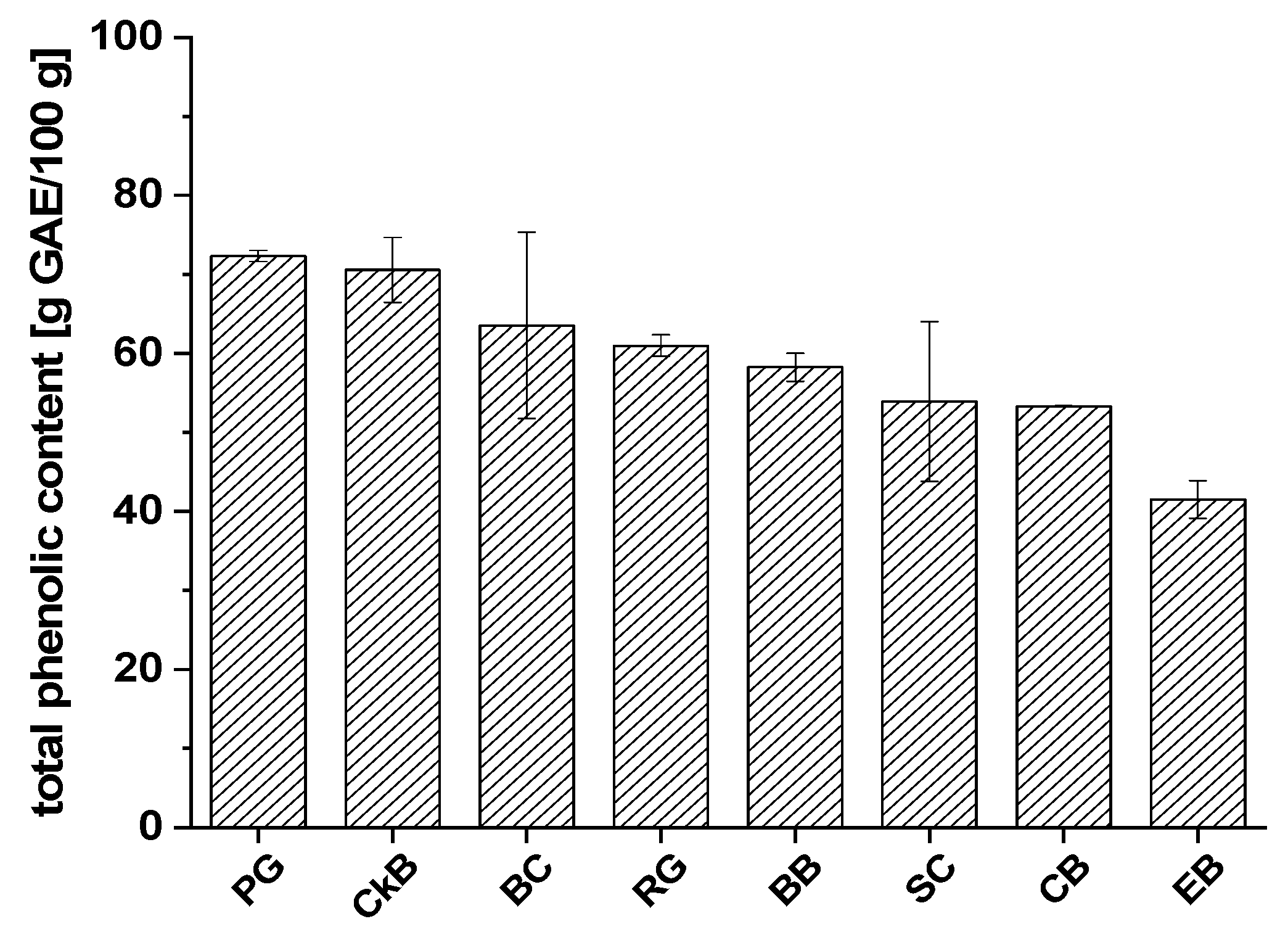
2.2. The Folin–Ciocalteu Assay
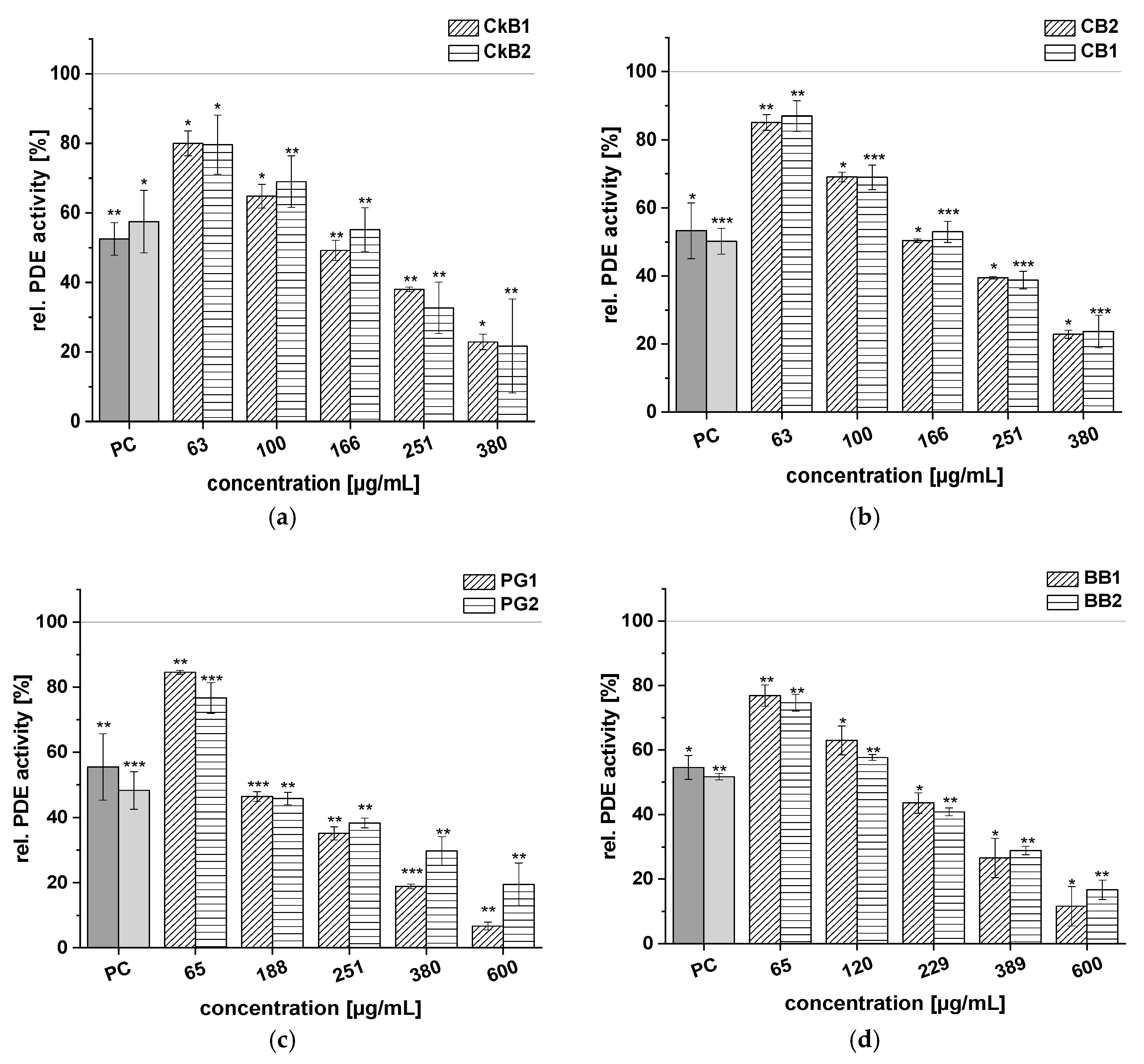
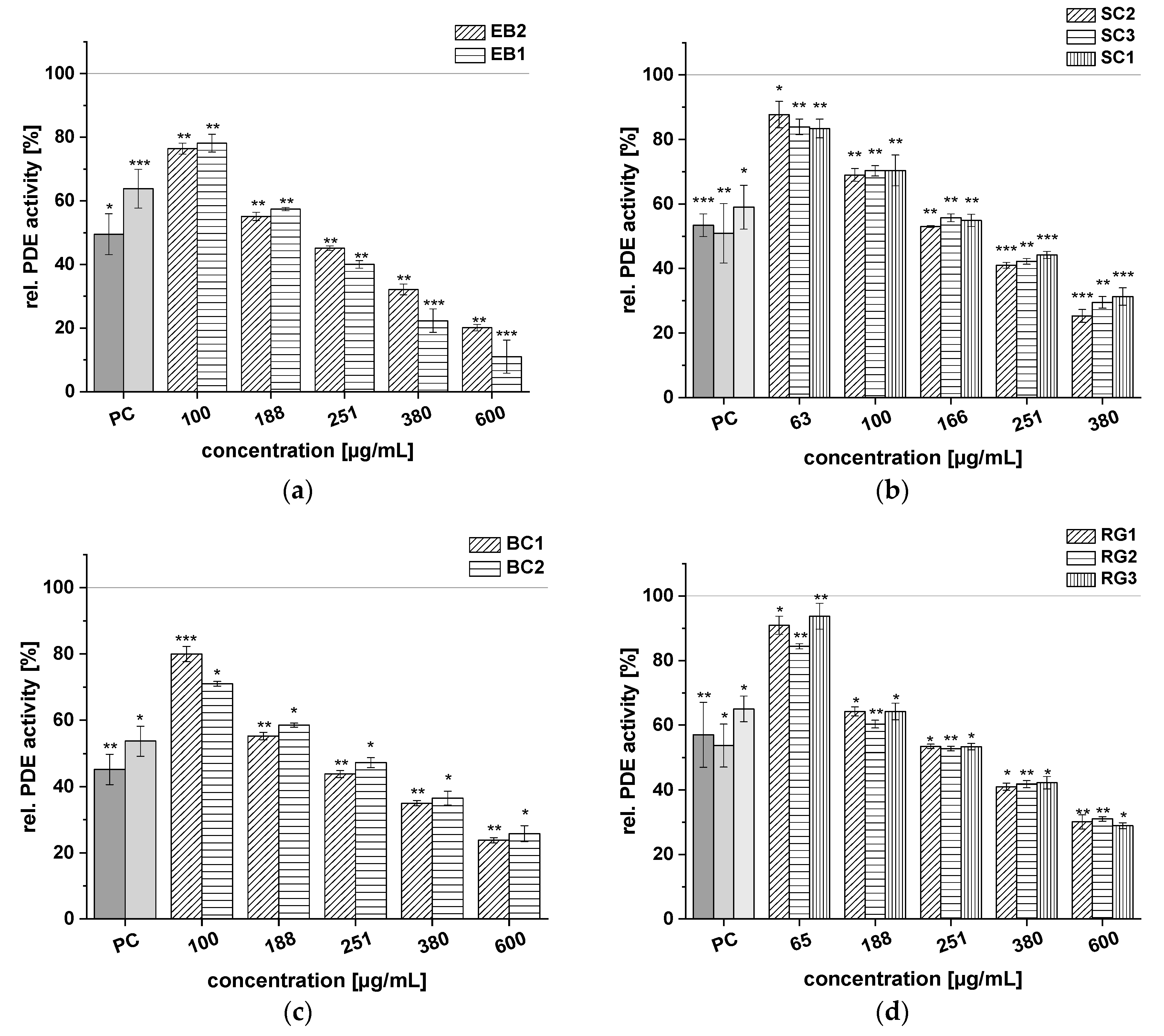
2.3. Inhibitory Effects of Anthocyanin-Rich Fruit Juice Extracts on PDE 3B Activity In Vitro
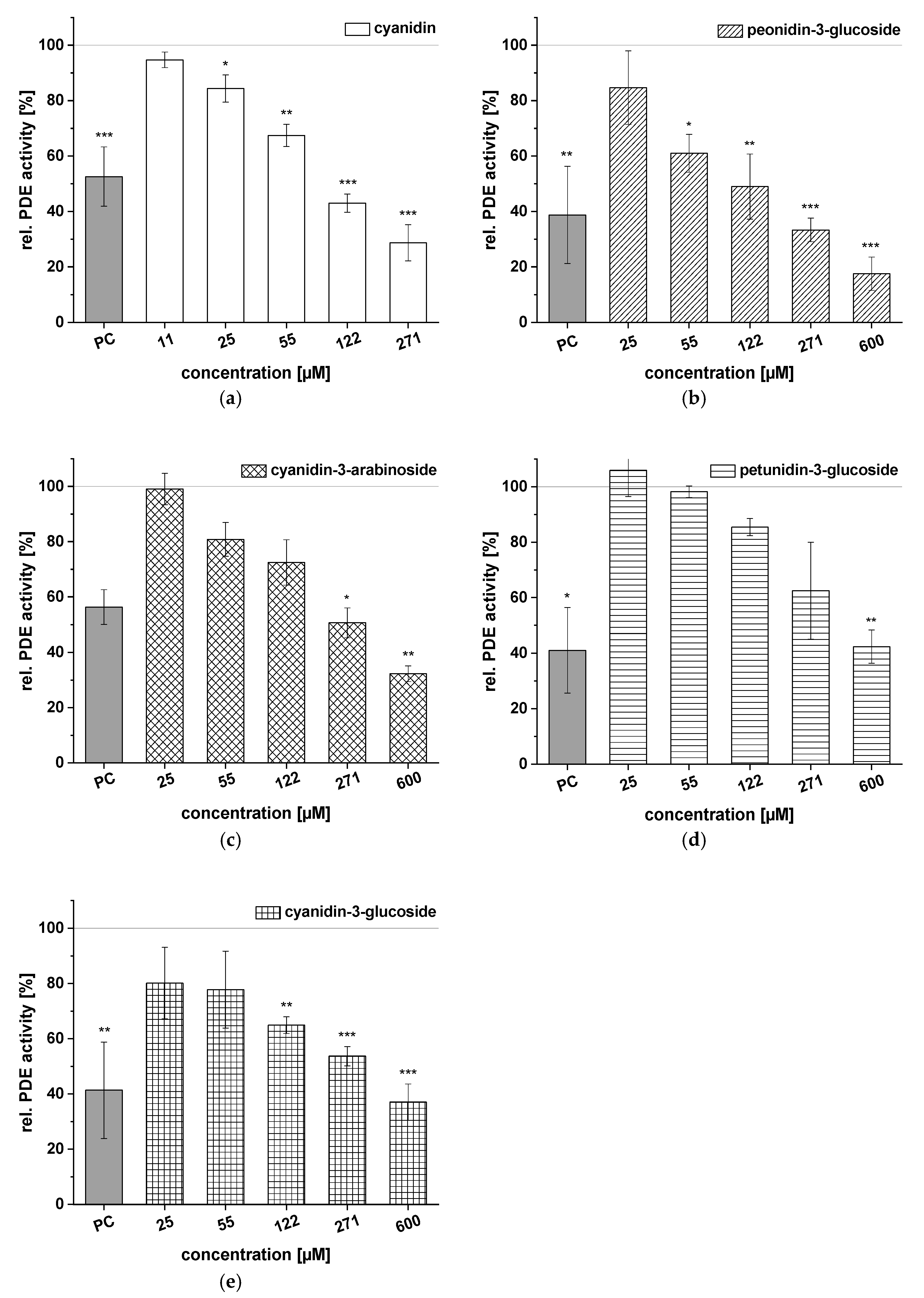
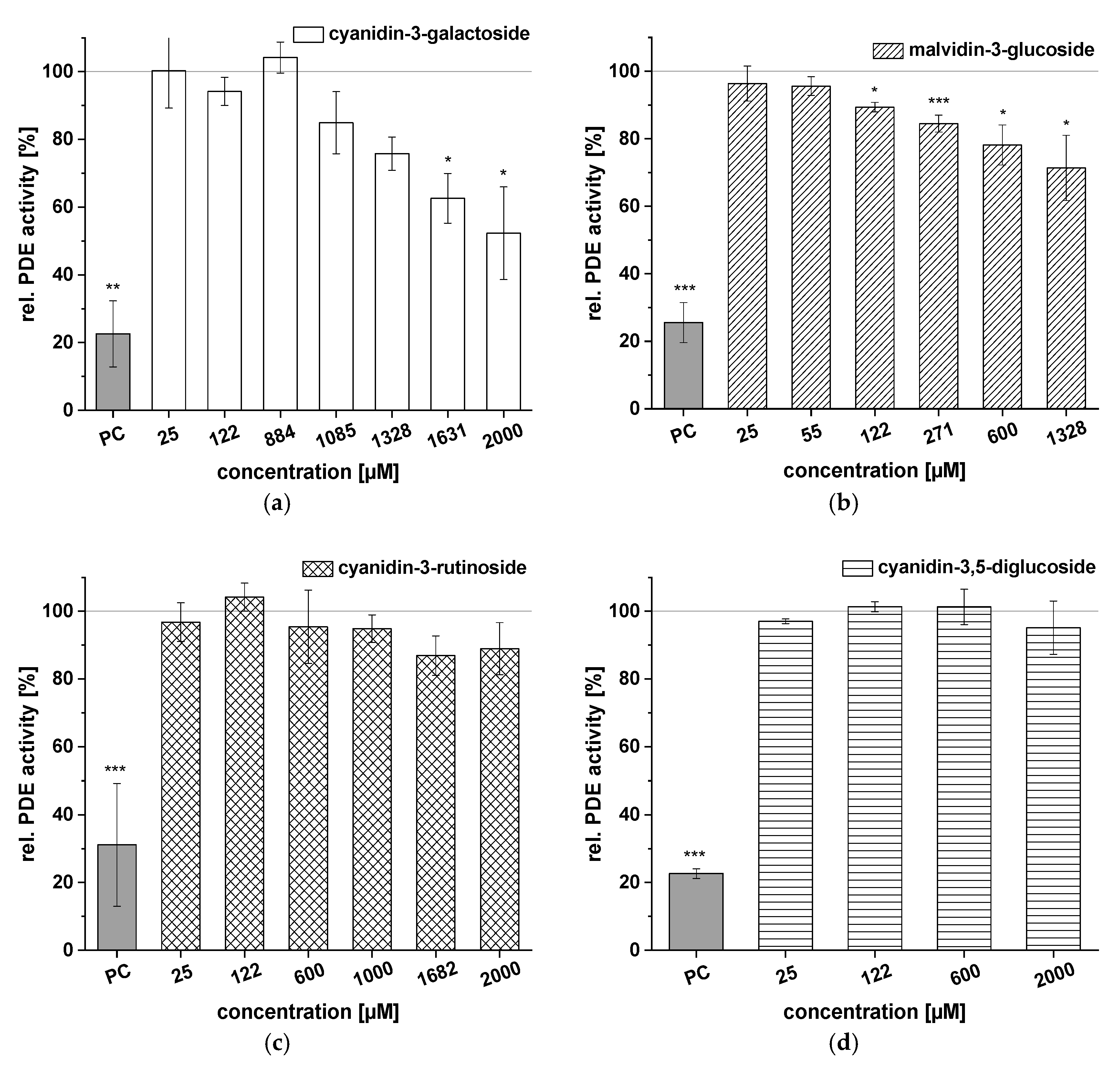
2.4. Inhibitory Effects of Selected Anthocyanins on PDE 3B Activity In Vitro
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Samples
4.3. HPLC-ESI-MS/MS Analysis
4.4. The Folin–Ciocalteu Assay
4.5. The cAMP-Specific PDE Activity Assay
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | anthocyanin |
| BB | blueberry |
| BC | black currant |
| cAMP | 3′,5′-cyclic adenosine monophosphate |
| CB | cranberry |
| cGMP | 3′,5′-cyclic guanosine monophosphate |
| CkB | chokeberry |
| DMSO | dimethyl sulfoxide |
| EB | elderberry |
| FFA | free fatty acid |
| GAE | gallic acid equivalents |
| HPLC-ESI-MSn | high-performance liquid chromatography with electrospray ionization mass spectrometry |
| IC50 value | half maximal inhibitory concentration |
| JC | juice concentrate |
| n.d. | not determined |
| NFC | not from concentrate, direct juices |
| PC | positive control |
| PDE | phosphodiesterase |
| PG | pomegranate |
| rel. | relative |
| RG | red grape |
| SC | sour cherry |
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 29 June 2020).
- Williamson, G. The role of polyphenols in modern nutrition. Nutr. Bull. 2017, 42, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentrations of anthocyanins in common foods in the United States and estimation of normal consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef] [PubMed]
- Meskin, M.S.; Bidlack, W.R.; Davies, A.J.; Lewis, D.S.; Randolph, R.K. Phytochemicals: Mechanisms of Action. Absorption and Metabolism of Anthocyanins: Potential Health Effects, 1st ed.; Prior, R.L., Ed.; CRC Press: Boca Raton, FL, USA, 2004; ISBN 9780849316722. [Google Scholar]
- De Pascual-Teresa, S.; Sanchez-Ballesta, M.T. Anthocyanins: From plant to health. Phytochem. Rev. 2008, 7, 281–299. [Google Scholar] [CrossRef]
- Weisel, T.; Baum, M.; Eisenbrand, G.; Dietrich, H.; Will, F.; Stockis, J.-P.; Kulling, S.; Rüfer, C.; Johannes, C.; Janzowski, C. An anthocyanin/polyphenolic-rich fruit juice reduces oxidative DNA damage and increases glutathione level in healthy probands. Biotechnol. J. 2006, 1, 388–397. [Google Scholar] [CrossRef]
- Kuntz, S.; Kunz, C.; Herrmann, J.; Borsch, C.H.; Abel, G.; Fröhling, B.; Dietrich, H.; Rudloff, S. Anthocyanins from fruit juices improve the antioxidant status of healthy young female volunteers without affecting anti-inflammatory parameters: Results from the randomised, double-blind, placebo-controlled, cross-over ANTHONIA (ANTHOcyanins in Nutrition Investigation Alliance) study. Br. J. Nutr. 2014, 112, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Triebel, S.; Trieu, H.-L.; Richling, E. Modulation of inflammatory gene expression by a bilberry (Vaccinium myrtillus L.) extract and single anthocyanins considering their limited stability under cell culture conditions. J. Agric. Food Chem. 2012, 60, 8902–8910. [Google Scholar] [CrossRef] [PubMed]
- Schantz, M.; Mohn, C.; Baum, M.; Richling, E. Antioxidative efficiency of an anthocyanin rich bilberry extract in the human colon tumor cell lines Caco-2 and HT-29. J. Berry Res. 2010, 1, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Bakuradze, T.; Tausend, A.; Galan, J.; Groh, I.A.M.; Berry, D.; Tur, J.A.; Marko, D.; Richling, E. Antioxidative activity and health benefits of anthocyanin-rich fruit juice in healthy volunteers. Free Radic. Res. 2019, 53, 1045–1055. [Google Scholar] [CrossRef]
- Geetha, T.; Malhotra, V.; Chopra, K.; Kaur, I.P. Antimutagenic and antioxidant/prooxidant activity of quercetin. Indian J. Exp. Biol. 2005, 43, 61–67. [Google Scholar]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, flavones, flavanones, and human health: Epidemiological evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef]
- Kuppusamy, U.R.; Das, N.P. Effects of flavonoids on cyclic AMP phosphodiesterase and lipid mobilization in rat adipocytes. Biochem. Pharmacol. 1992, 44, 1307–1315. [Google Scholar] [CrossRef]
- Dallas, C.; Gerbi, A.; Tenca, G.; Juchaux, F.; Bernard, F.-X. Lipolytic effect of a polyphenolic citrus dry extract of red orange, grapefruit, orange (SINETROL) in human body fat adipocytes. Mechanism of action by inhibition of cAMP-phosphodiesterase (PDE). Phytomedicine 2008, 15, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Girotti, C.; Ginet, M.; Demarne, F.C.; Lagarde, M.; Géloën, A. Lipolytic activity of cirsimarin extracted from Microtea debilis. Planta Med. 2005, 71, 1170–1172. [Google Scholar] [CrossRef]
- Nielsen, T.S.; Jessen, N.; Jørgensen, J.O.L.; Møller, N.; Lund, S. Dissecting adipose tissue lipolysis: Molecular regulation and implications for metabolic disease. J. Mol. Endocrinol. 2014, 52, R199–R222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spector, A.A. Fatty acid binding to plasma albumin. J. Lipid Res. 1975, 16, 165–179. [Google Scholar]
- Boswell-Smith, V.; Spina, D.; Page, C.P. Phosphodiesterase inhibitors. Br. J. Pharmacol. 2006, 147 (Suppl. 1), S252–S257. [Google Scholar] [CrossRef]
- Gresele, P.; Momi, S.; Falcinelli, E. Anti-platelet therapy: Phosphodiesterase inhibitors. Br. J. Clin. Pharmacol. 2011, 72, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Rall, T.; Sutherland, E.W. Formation of a cyclic adenine ribonucleotide by tissue particles. J. Biol. Chem. 1958, 232, 1065–1076. [Google Scholar]
- Butcher, R.; Sutherland, E. Adenosine 3′,5′-phosphate in biological materials. J. Biol. Chem. 1962, 237, 1244–1250. [Google Scholar]
- Marko, D.; Puppel, N.; Tjaden, Z.; Jakobs, S.; Pahlke, G. The substitution pattern of anthocyanidins affects different cellular signaling cascades regulating cell proliferation. Mol. Nutr. Food Res. 2004, 48, 318–325. [Google Scholar] [CrossRef]
- Ko, W.-C.; Shih, C.-M.; Lai, Y.-H.; Chen, J.-H.; Huang, H.-L. Inhibitory effects of flavonoids on phosphodiesterase isozymes from guinea pig and their structure-activity relationships. Biochem. Pharmacol. 2004, 68, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Beretz, A.; Anton, R.; Stoclet, J.C. Flavonoid compounds are potent inhibitors of cyclic AMP phosphodiesterase. Experientia 1978, 34, 1054–1055. [Google Scholar] [CrossRef] [PubMed]
- Röhrig, T.; Pacjuk, O.; Hernández-Huguet, S.; Körner, J.; Scherer, K.; Richling, E. Inhibition of Cyclic Adenosine Monophosphate-Specific Phosphodiesterase by Various Food Plant-Derived Phytotherapeutic Agents. Medicines 2017, 4, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dell’Agli, M.; Galli, G.V.; Vrhovsek, U.; Mattivi, F.; Bosisio, E. In vitro inhibition of human cGMP-specific phosphodiesterase-5 by polyphenols from red grapes. J. Agric. Food Chem. 2005, 53, 1960–1965. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Hillebrand, S.; Winterhalter, P. Analysis of phenolic components of black chokeberries (Aronia melanocarpa). Dtsch. Lebensm.-Rundsch. 2010, 106, 374–382. [Google Scholar]
- Pawlowska, A.M.; de Leo, M.; Braca, A. Phenolics of Arbutus unedo L. (Ericaceae) fruits: Identification of anthocyanins and gallic acid derivatives. J. Agric. Food Chem. 2006, 54, 10234–10238. [Google Scholar] [CrossRef]
- Rauf, A.; Orhan, I.E.; Ertas, A.; Temel, H.; Hadda, T.B.; Saleem, M.; Raza, M.; Khan, H. Elucidation of Phosphodiesterase-1 Inhibitory Effect of Some Selected Natural Polyphenolics Using In Vitro and In Silico Methods. Curr. Top. Med. Chem. 2017, 17, 412–417. [Google Scholar] [CrossRef]
- Montoya, G.A.; Bakuradze, T.; Eirich, M.; Erk, T.; Baum, M.; Habermeyer, M.; Eisenbrand, G.; Richling, E. Modulation of 3′,5′-cyclic AMP homeostasis in human platelets by coffee and individual coffee constituents. Br. J. Nutr. 2014, 112, 1427–1437. [Google Scholar] [CrossRef] [Green Version]
- Ostberg-Potthoff, J.J.; Berger, K.; Richling, E.; Winterhalter, P. Activity-Guided Fractionation of Red Fruit Extracts for the Identification of Compounds Influencing Glucose Metabolism. Nutrients 2019, 11, 1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pöch, G. Assay of phosphodiesterase with radioactively labeled cyclic 3′,5′-AMP as substrate. Naunyn-Sehmiedebergs Arch. Pharmak. 1971, 268, 272–299. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Red Fruit | Number of Anthocyanins Identified | Major Anthocyanins | [M + H]+ m/z | Fragments m/z |
|---|---|---|---|---|
| chokeberry | 6 | Cyanidin-3-galactoside (a) | 449 | 287 |
| Cyanidin-3-arabinoside (a) | 419 | 287 | ||
| cranberry | 7 | Peonidin-3-galactoside (b) | 463 | 301 |
| Cyanidin-3-galactoside (b) | 449 | 287 | ||
| pomegranate | 4 | Cyanidin-3,5-diglucoside (b) | 611 | 287 |
| Cyanidin-3-glucoside (a) | 449 | 287 | ||
| blueberry | 14 | Peonidin-3-galactoside (b) | 463 | 301 |
| Cyanidin-3-galactoside (a) | 449 | 287 | ||
| elderberry | 4 | Cyanidin-3-sambubioside (b) | 581 | 287 |
| red grape | 21 | Malvidin-3-glucoside (a) | 493 | 331 |
| Peonidin-3-glucoside (a) | 463 | 301 | ||
| sour cherry | 4 | Cyanidin-3-(2G-glucosylrutinoside) (b) | 757 | 287 |
| Cyanidin-3-rutinoside (b) | 595 | 287 | ||
| black currant | 5 | Cyanidin-3-rutinoside (b) | 595 | 287 |
| Delphinidin-3-rutinoside (b) | 611 | 303 |
| Red Fruit | Number of Copigments Identified | Major Copigments | [M − H]− m/z | Fragments m/z |
|---|---|---|---|---|
| chokeberry | 10 | Chlorogenic acid (a) | 353 | 191, 179, 161 |
| Neochlorogenic acid (a) | 353 | 191, 179, 135 | ||
| cranberry | 26 | Quercetin-hexoside (b) | 463 | 301 |
| Myricetin-hexoside (b) | 479 | 316 | ||
| pomegranate | 28 | Pedunculagine (b) | 783 | 301 |
| Punicalagin (a) | 1083 | 601 | ||
| blueberry | 17 | Chlorogenic acid (a) | 353 | 191 |
| Cumaroyliridoid (b) | 535 | 371 | ||
| Quercetin-hexoside (b) | 463 | 301 | ||
| elderberry | 10 | Quercetin-3-rutinoside (a) | 609 | 301 |
| Quercetin-3-glucoside (a) | 463 | 301 | ||
| red grape | 9 | Isorhamnetin-hexoside (b) | 479 | 316 |
| Quercetin-hexoside (b) | 463 | 301 | ||
| sour cherry | 10 | Quercetin-3-(2G-glucosylrutinoside) (b) | 771 | 301 |
| Quercetin-3-rutinoside (a) | 609 | 301 | ||
| black currant | 11 | Isorhamnetin-rutinoside (b) | 625 | 317 |
| Extract | Abbreviation | Product | IC50 (µg/mL) |
|---|---|---|---|
| chokeberry | CkB1 | JC | 163 ± 3 |
| CkB2 | JC | 167 ± 5 | |
| blueberry | BB1 | JC | 174 ± 3 |
| BB2 | NFC | 165 ± 2 | |
| pomegranate | PG1 | JC | 169 ± 2 |
| PG2 | JC | 174 ± 1 | |
| cranberry | CB1 | JC | 175 ± 1 |
| CB2 | JC | 180 ± 3 | |
| sour cherry | SC1 | NFC | 185 ± 1 |
| SC2 | JC | 195 ± 2 | |
| SC3 | JC | 200 ± 2 | |
| elderberry | EB1 | JC | 222 ± 3 |
| EB2 | JC | 206 ± 2 | |
| black currant | BC1 | NFC | 212 ± 1 |
| BC2 | JC | 229 ± 3 | |
| red grape | RG1 | JC | 290 ± 2 |
| RG2 | NFC | 276 ± 2 | |
| RG3 | JC | 292 ± 3 |
| Anthocyanin/Anthocyanidin | IC50 (µM) |
|---|---|
| cyanidin | 105 ± 11 |
| peonidin-3-glucoside | 120 ± 44 |
| cyanidin-3-arabinoside | 257 ± 13 |
| cyanidin-3-glucoside | 299 ± 56 |
| petunidin-3-glucoside | 448 ± 39 |
| cyanidin-3-galactoside | n.d. |
| cyanidin-3-rutinoside | n.d. |
| cyanidin-3,5-diglucoside | n.d. |
| malvidin-3-glucoside | n.d. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Göttel, C.; Niesen, S.; Daub, V.; Werle, T.; Bakuradze, T.; Winterhalter, P.; Richling, E. In Vitro Inhibition of Phosphodiesterase 3B (PDE 3B) by Anthocyanin-Rich Fruit Juice Extracts and Selected Anthocyanins. Int. J. Mol. Sci. 2020, 21, 6934. https://doi.org/10.3390/ijms21186934
Göttel C, Niesen S, Daub V, Werle T, Bakuradze T, Winterhalter P, Richling E. In Vitro Inhibition of Phosphodiesterase 3B (PDE 3B) by Anthocyanin-Rich Fruit Juice Extracts and Selected Anthocyanins. International Journal of Molecular Sciences. 2020; 21(18):6934. https://doi.org/10.3390/ijms21186934
Chicago/Turabian StyleGöttel, Celina, Sonja Niesen, Vanessa Daub, Theresa Werle, Tamara Bakuradze, Peter Winterhalter, and Elke Richling. 2020. "In Vitro Inhibition of Phosphodiesterase 3B (PDE 3B) by Anthocyanin-Rich Fruit Juice Extracts and Selected Anthocyanins" International Journal of Molecular Sciences 21, no. 18: 6934. https://doi.org/10.3390/ijms21186934
APA StyleGöttel, C., Niesen, S., Daub, V., Werle, T., Bakuradze, T., Winterhalter, P., & Richling, E. (2020). In Vitro Inhibition of Phosphodiesterase 3B (PDE 3B) by Anthocyanin-Rich Fruit Juice Extracts and Selected Anthocyanins. International Journal of Molecular Sciences, 21(18), 6934. https://doi.org/10.3390/ijms21186934