Pediatric Tuberculosis: The Impact of “Omics” on Diagnostics Development



{kind=link}
{kind=link}
Abstract
:1. Introduction
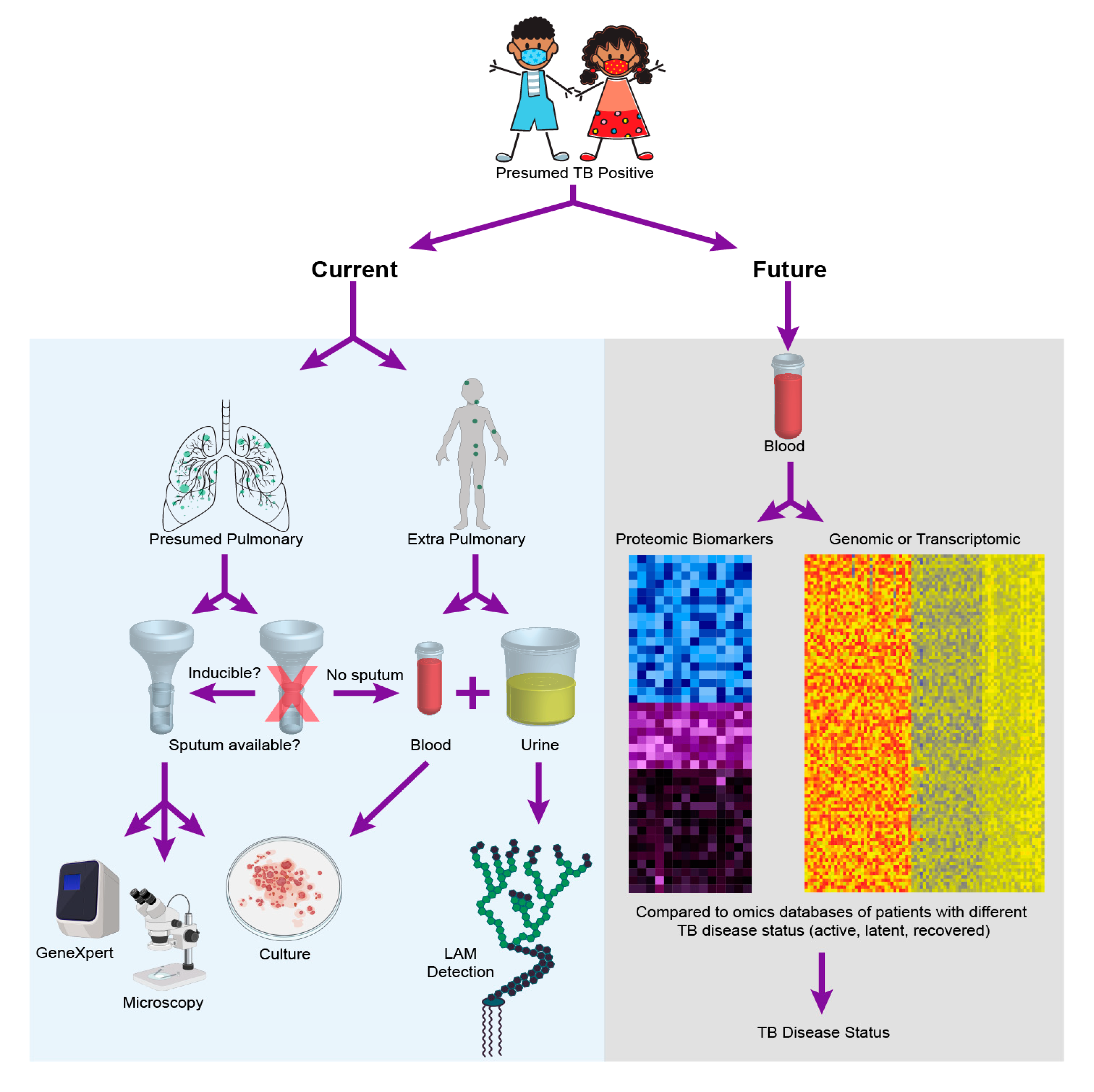
2. Current Diagnostics for Pediatric TB
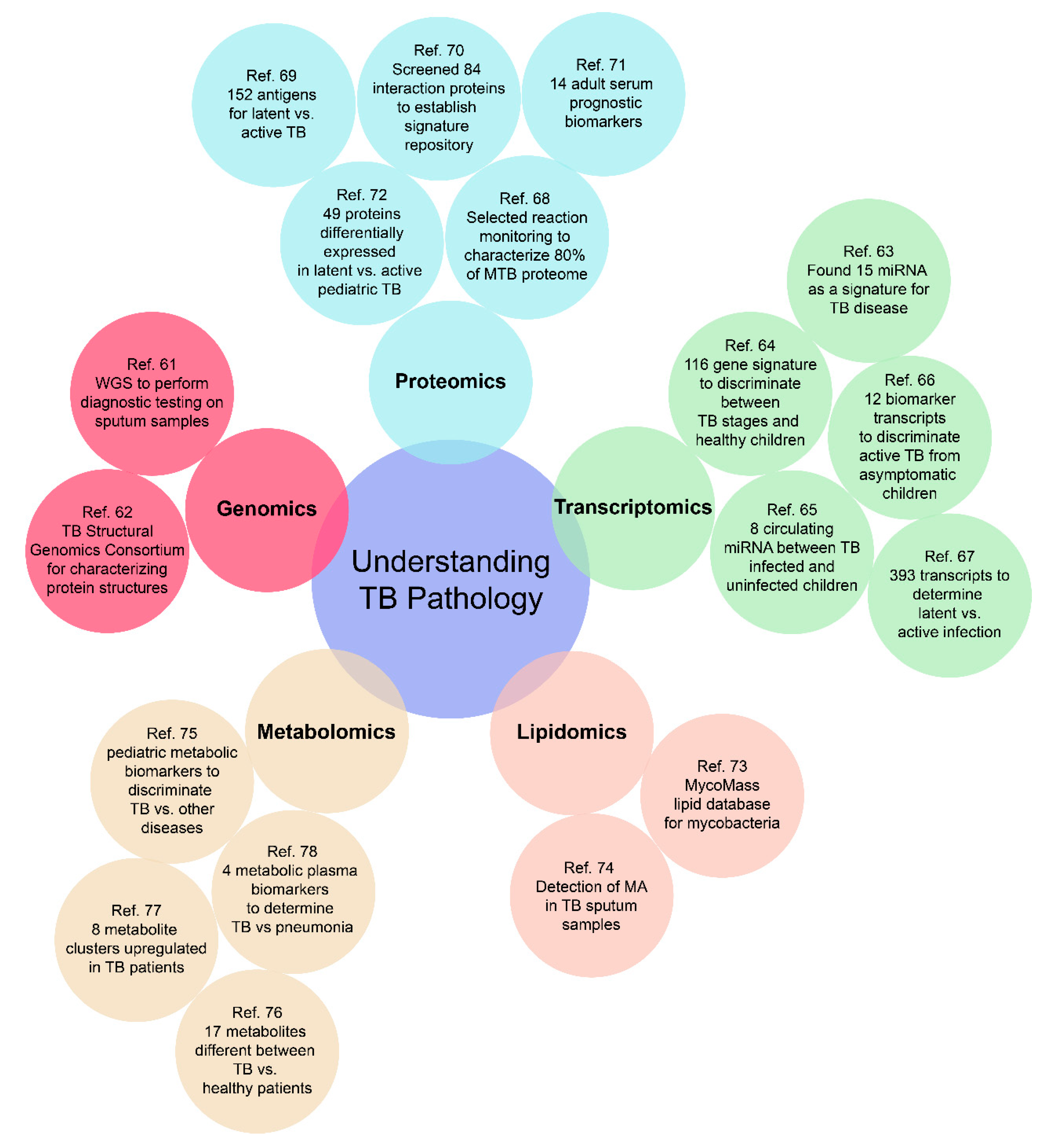
3. The Role of “Omics” in TB Diagnostic Development
3.1. Genomics
3.2. Transcriptomics
3.3. Proteomics
3.4. Lipidomics
3.5. Metabolomics
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- MacLean, E.; Sulis, G.; Denkinger, C.M.; Johnston, J.C.; Pai, M.; Khana, F.A. Diagnostic accuracy of Stool Xpert MTB/RIF for detection of pulmonary tuberculosis in children: A Systematic Review and Meta-analysis. J. Clin. Microbiol. 2019, 57, e02057-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Tuberculosis Report; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- WHO. Roadmap towards Ending TB in Children and Adolescents, 2nd ed.; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Starke, J.R. Transmission of Mycobacterium tuberculosis to and from children and adolescents. Semin. Pediatr. Infect. Dis. 2001, 12, 115–123. [Google Scholar] [CrossRef]
- Flynn, J.L.; Chan, J. Tuberculosis: Latency and reactivation. Infect. Immun. 2001, 69, 4195–4201. [Google Scholar] [CrossRef] [Green Version]
- Kapur, A.; Harries, A.D.; Lönnroth, K.; Wilson, P.; Sulistyowati, L.S. Diabetes and tuberculosis co-epidemic: The Bali Declaration. Lancet Diabetes Endocrinol. 2016, 4, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Sharan, R.; Bucşan, A.N.; Ganatra, S.; Paiardini, M.; Mohan, M.; Mehra, S.; Khader, S.A.; Kaushal, D. Chronic Immune Activation in TB/HIV Co-infection. Trends Microbiol. 2020, 28, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Wiegeshaus, E.H.; Taylor, B.T.; Smith, D.W. Pathogenesis of tuberculosis: Pathway to apical localization. Tuber. Lung Dis. 1994, 75, 68–78. [Google Scholar] [CrossRef]
- Riley, R.L. Airborne infection. Am. J. Med. 1974, 57, 466–475. [Google Scholar] [CrossRef]
- Whittaker, E.; Lopez-Varela, E.; Broderick, C.; Seddon, J.A. Examining the complex relationship between tuberculosis and other infectious diseases in children: A review. Front. Pediatr. 2019, 7, 233. [Google Scholar] [CrossRef] [Green Version]
- Nhu, N.T.Q.; Ha, D.T.M.; Anh, N.D.; Thu, D.D.A.; Duong, T.N.; Quang, N.D.; Lan, N.T.N.; Van Quyet, T.; Tuyen, N.T.B.; Ha, V.T.; et al. Evaluation of Xpert MTB/RIF and MODS assay for the diagnosis of pediatric tuberculosis. BMC Infect. Dis. 2013, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Marais, B.J.; Donald, P.R.; Gie, R.P.; Schaaf, H.S.; Beyers, N. Diversity of disease in childhood pulmonary tuberculosis. Ann. Trop. Paediatr. 2005, 25, 79–86. [Google Scholar] [CrossRef]
- Comstock, G.W.; Livesay, V.T.; Woolpert, S.F. The prognosis of a positive tuberculin reaction in childhood and adolescence. Am. J. Epidemiol. 1974, 99, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Marais, B.J.; Graham, S.M.; Maeurer, M.; Zumla, A. Progress and challenges in childhood tuberculosis. Lancet Infect. Dis. 2013, 13, 287–289. [Google Scholar] [CrossRef]
- Mandal, N.; Anand, P.K.; Gautam, S.; Das, S.; Hussain, T. Diagnosis and treatment of paediatric tuberculosis: An insight review. Crit. Rev. Microbiol. 2017, 43, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Lacourse, S.M.; Pavlinac, P.B.; Cranmer, L.M.; Njuguna, I.N.; Mugo, C.; Gatimu, J.; Stern, J.; Walson, J.L.; Maleche-Obimbo, E.; Oyugi, J.; et al. Stool Xpert MTB/RIF and urine lipoarabinomannan for the diagnosis of tuberculosis in hospitalized HIV-infected children. AIDS 2018, 32, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.J.; Starke, J.R.; Revell, P.A. Laboratory Diagnosis of Mycobacterium tuberculosis Infection and Disease in Children. J. Clin. Microbiol. 2016, 54, 1434–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, S.; Rekha, B. Pediatric Tuberculosis: Global Overview and Challenges. Clin. Infect. Dis. 2010, 50, S184–S194. [Google Scholar] [CrossRef]
- Zar, H.J.; Connell, T.G.; Nicol, M. Diagnosis of pulmonary tuberculosis in children: New advances. Expert Rev. Anti Infect. Ther. 2010, 8, 277–288. [Google Scholar] [CrossRef]
- Graham, S.M.; Ahmed, T.; Amanullah, F.; Browning, R.; Cardenas, V.; Casenghi, M.; Cuevas, L.E.; Gale, M.; Gie, R.P.; Grzemska, M.; et al. Evaluation of tuberculosis diagnostics in children: 1. Proposed clinical case definitions for classification of intrathoracic tuberculosis disease. Consensus from an expert panel. J. Infect. Dis. 2012. [Google Scholar] [CrossRef] [Green Version]
- Perez-Velez, C.M.; Marais, B.J. Tuberculosis in Children. N. Engl. J. Med. 2012, 367, 348–361. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.J.; Wells, C.D. Global epidemiology of childhood tuberculosis. Int. J. Tuberc. Lung Dis. 2004, 8, 636–647. [Google Scholar]
- Chiappini, E.; Lo Vecchio, A.; Garazzino, S.; Marseglia, G.L.; Bernardi, F.; Castagnola, E.; Tomà, P.; Cirillo, D.; Russo, C.; Gabiano, C.; et al. Recommendations for the diagnosis of pediatric tuberculosis. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sakamuri, R.M.; Moodley, P.; Yusim, K.; Feng, S.; Sturm, A.W.; Korber, B.T.M.; Mukundan, H. Current methods for diagnosis of human tuberculosis and considerations for global surveillance. In Tuberculosis, Leprosy and Other Mycobacterial Diseases of Man and Animals; CABI: Wallingford, UK, 2015; pp. 72–102. ISBN 978-178-064-396-0. [Google Scholar]
- Bianchi, L.; Galli, L.; Moriondo, M.; Veneruso, G.; Becciolini, L.; Azzari, C.; Chiappini, E.; De Martino, M. Interferon-gamma release assay improves the diagnosis of tuberculosis in children. Pediatr. Infect. Dis. J. 2009, 28, 510–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliot, C.; Marais, B.; Williams, P.; Joshua, P.; Towle, S.; Hart, G.; Zwi, K. Tuberculin skin test versus interferon-gamma release assay in refugee children: A retrospective cohort study. J. Paediatr. Child Health 2018, 54, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Detjen, A.K.; Keil, T.; Wahn, U.; Magdorf, K.; Mauch, H.; Roll, S.; Hauer, B. Interferon-Release Assays Improve the Diagnosis of Tuberculosis and Nontuberculous Mycobacterial Disease in Children in a Country with a Low Incidence of Tuberculosis. Clin. Infect. Dis. 2007, 45, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Aber, V.R.; Allen, B.W.; Mitchison, D.A.; Ayuma, P.; Edwards, E.A.; Keyes, A.B. Quality control in tuberculosis bacteriology. I. Laboratory studies on isolated positive cultures and the efficiency of direct smear examination. Tubercle 1980, 61, 123–133. [Google Scholar] [CrossRef]
- Burdash, N.M.; Manos, J.P.; Ross, D.; Bannister, E.R. Evaluation of the acid fast smear. J. Clin. Microbiol. 1976, 4, 190–191. [Google Scholar]
- Corper, H.J.; Stoner, R.E. An improved procedure for the diagnostic culture of mammalian tubercle bacilli. J. Lab. Clin. Med. 1946, 31, 1364–1371. [Google Scholar]
- Dye, C.; Watt, C.J.; Bleed, D.M.; Williams, B.G. What is the limit to case detection under the DOTS strategy for tuberculosis control? Tuberculosis 2003, 83, 35–43. [Google Scholar] [CrossRef]
- Hopewell, P.C.; Pai, M.; Maher, D.; Uplekar, M.; Raviglione, M.C. International Standards for Tuberculosis Care. Lancet Infect. Dis. 2006, 6, 710–725. [Google Scholar] [CrossRef]
- Kunkel, A.; Abel zur Wiesch, P.; Nathavitharana, R.R.; Marx, F.M.; Jenkins, H.E.; Cohen, T. Smear positivity in paediatric and adult tuberculosis: Systematic review and meta-analysis. BMC Infect. Dis. 2016, 16, 282. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Tuberculosis Report 2018; World Health Organization: Geneva, Switzerland, 2018; ISBN 978-924-156-551-6. [Google Scholar]
- Dorman, S.E.; Schumacher, S.G.; Alland, D.; Nabeta, P.; Armstrong, D.T.; King, B.; Hall, S.L.; Chakravorty, S.; Cirillo, D.M.; Tukvadze, N.; et al. Xpert MTB/RIF Ultra for detection of Mycobacterium tuberculosis and rifampicin resistance: A prospective multicentre diagnostic accuracy study. Lancet Infect. Dis. 2018, 18, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Steingart, K.R.; Schiller, I.; Horne, D.J.; Pai, M.; Boehme, C.C.; Dendukuri, N. Xpert(R) MTB/RIF assay for pulmonary tuberculosis and rifampicin resistance in adults. Cochrane Database Syst. Rev. 2014, 2014, CD009593. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. High-Priority Target Product Profiles for New Tuberculosis Diagnostics: Report of a Consensus Meeting; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Bjerrum, S.; Schiller, I.; Dendukuri, N.; Kohli, M.; Nathavitharana, R.R.; Zwerling, A.A.; Denkinger, C.M.; Steingart, K.R.; Shah, M. Lateral flow urine lipoarabinomannan assay for detecting active tuberculosis in people living with HIV. Cochrane Database Syst. Rev. 2019, 10, CD011420. [Google Scholar] [CrossRef] [Green Version]
- Nicol, M.P.; Allen, V.; Workman, L.; Isaacs, W.; Munro, J.; Pienaar, S.; Black, F.; Adonis, L.; Zemanay, W.; Ghebrekristos, Y.; et al. Urine lipoarabinomannan testing for diagnosis of pulmonary tuberculosis in children: A prospective study. Lancet Glob. Health 2014, 2, e278–e284. [Google Scholar] [CrossRef] [Green Version]
- Iskandar, A.; Nursiloningrum, E.; Arthamin, M.Z.; Olivianto, E.; Chandrakusuma, M.S. The diagnostic value of urine lipoarabinomannan (LAM) antigen in childhood tuberculosis. J. Clin. Diagn. Res. 2017, 11, EC32–EC35. [Google Scholar] [CrossRef] [PubMed]
- Bulterys, M.A.; Wagner, B.; Redard-Jacot, M.; Suresh, A.; Pollock, N.R.; Moreau, E.; Denkinger, C.M.; Drain, P.K.; Broger, T. Point-Of-Care Urine LAM Tests for Tuberculosis Diagnosis: A Status Update. J. Clin. Med. 2019, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Mukundan, H.; Price, D.N.; Goertz, M.; Parthasarathi, R.; Montaño, G.A.; Kumar, S.; Scholfield, M.R.; Anderson, A.S.; Gnanakaran, S.; Iyer, S.; et al. Understanding the interaction of Lipoarabinomannan with membrane mimetic architectures. Tuberculosis 2012, 92, 38–47. [Google Scholar] [CrossRef]
- Mukundan, H.; Kumar, S.; Price, D.N.; Ray, S.M.; Lee, Y.-J.; Min, S.; Eum, S.; Kubicek-Sutherland, J.; Resnick, J.M.; Grace, W.K.; et al. Rapid detection of Mycobacterium tuberculosis biomarkers in a sandwich immunoassay format using a waveguide-based optical biosensor. Tuberculosis 2012, 92, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Jakhar, S.; Lenz, K.; Mukundan, H. Current Status of Pediatric Tuberculosis Diagnostics, Needs, and Challenges. In Diagnosis and Management of Tuberculosis; Open Access ebooks: Las Vegas, NV, USA, 2019; pp. 1–13. ISBN 978-93-87500-67-9. [Google Scholar]
- Sakamuri, R.M.; Price, D.N.; Lee, M.; Cho, S.N.; Barry, C.E.; Via, L.E.; Swanson, B.I.; Mukundan, H. Association of lipoarabinomannan with high density lipoprotein in blood: Implications for diagnostics. Tuberculosis 2013, 93, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Amin, A.G.; De, P.; Spencer, J.S.; Brennan, P.J.; Daum, J.; Andre, B.G.; Joe, M.; Bai, Y.; Laurentius, L.; Porter, M.D.; et al. Detection of lipoarabinomannan in urine and serum of HIV-positive and HIV-negative TB suspects using an improved capture-enzyme linked immuno absorbent assay and gas chromatography/mass spectrometry. Tuberculosis 2018, 111, 178–187. [Google Scholar] [CrossRef]
- Broger, T.; Tsionksy, M.; Mathew, A.; Lowary, T.L.; Pinter, A.; Plisova, T.; Bartlett, D.; Barbero, S.; Denkinger, C.M.; Moreau, E.; et al. Sensitive electrochemiluminescence (ECL) immunoassays for detecting lipoarabinomannan (LAM) and ESAT-6 in urine and serum from tuberculosis patients. PLoS ONE 2019, 14, e0215443. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, S.G.; Van Smeden, M.; Dendukuri, N.; Joseph, L.; Nicol, M.P.; Pai, M.; Zar, H.J. Diagnostic Test Accuracy in Childhood Pulmonary Tuberculosis: A Bayesian Latent Class Analysis. Am. J. Epidemiol. 2016, 184, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Starke, J.R. Diagnosis of tuberculosis in children. Pediatr. Infect. Dis. J. 2000, 19, 1095–1096. [Google Scholar] [CrossRef] [PubMed]
- Geyer, P.E.; Holdt, L.M.; Teupser, D.; Mann, M. Revisiting biomarker discovery by plasma proteomics. Mol. Syst. Biol. 2017, 13, 942. [Google Scholar] [CrossRef] [PubMed]
- Wetterstrand, K. DNA Sequencing Costs: Data from the NHGRI Genome Sequencing Program (GSP). 2020. Available online: https://www.genome.gov/about-genomics/fact-sheets/DNA-Sequencing-Costs-Data (accessed on 30 August 2020).
- Colston, S. Field forward Sequencing in Naval Environments. 2019. Available online: https://nanoporetech.com/resource-centre/field-forward-sequencing-naval-environments (accessed on 30 August 2020).
- Nimmo, C.; Shaw, L.P.; Doyle, R.; Williams, R.; Brien, K.; Burgess, C.; Breuer, J.; Balloux, F.; Pym, A.S. Whole genome sequencing Mycobacterium tuberculosis directly from sputum identifies more genetic diversity than sequencing from culture. BMC Genom. 2019, 20, 389. [Google Scholar] [CrossRef] [Green Version]
- MD Anderson Center “Submission, Services and Pricing”. Available online: https://www.mdanderson.org/research/research-resources/core-facilities/functional-proteomics-rppa-core/submission-services-and-pricing.html (accessed on 30 August 2020).
- Huang, Y.; Zhu, H. Protein Array-based Approaches for Biomarker Discovery in Cancer. Genom. Proteom. Bioinf. 2017, 15, 73–81. [Google Scholar] [CrossRef]
- Stanislaus, R.; Carey, M.; Deus, H.F.; Coombes, K.; Hennessy, B.T.; Mills, G.B.; Almeida, J.S. RPPAML/RIMS: A metadata format and an information management system for reverse phase protein arrays. BMC Bioinform. 2008, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Li, P.E.; Lo, C.C.; Anderson, J.J.; Davenport, K.W.; Bishop-Lilly, K.A.; Xu, Y.; Ahmed, S.; Feng, S.; Mokashi, V.P.; Chain, P.S.G. Enabling the democratization of the genomics revolution with a fully integrated web-based bioinformatics platform. Nucleic Acids Res. 2017, 45, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Posadas, E.M.; Simpkins, F.; Liotta, L.A.; MacDonald, C.; Kohn, E.C. Proteomic analysis for the early detection and rational treatment of cancer—Realistic hope? Ann. Oncol. 2005, 16, 16–22. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Mouritsen, J.; Schubert, O.T.; Aebersold, R.; Kaufmann, S.H.E. Mycobacterium tuberculosis in the Proteomics Era. Microbiol. Spectr. 2014, 2, 239–260. [Google Scholar] [CrossRef] [Green Version]
- Köser, C.U.; Bryant, J.M.; Becq, J.; Török, M.E.; Ellington, M.J.; Marti-Renom, M.A.; Carmichael, A.J.; Parkhill, J.; Smith, G.P.; Peacock, S.J. Whole-genome sequencing for rapid susceptibility testing of M. tuberculosis. N. Engl. J. Med. 2013, 369, 290–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, C.T.; Roe, J.K.; Pollara, G.; Mehta, M.; Noursadeghi, M. Diagnostic ‘omics’ for active tuberculosis. BMC Med. 2016, 14, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terwilliger, T.; Park, M.; Waldo, G.; Berendzen, J.; Hung, L.-W.; Kim, C.-Y.; Smith, C.; Sacchettini, J.; Bellinzoni, M.; Bossi, R.; et al. The TB structural genomics consortium: A resource for Mycobacterium tuberculosis biology. Tuberculosis 2003, 83, 223–249. [Google Scholar] [CrossRef]
- Miotto, P.; Mwangoka, G.; Valente, I.C.; Norbis, L.; Sotgiu, G.; Bosu, R.; Ambrosi, A.; Codecasa, L.R.; Goletti, D.; Matteelli, A.; et al. miRNA Signatures in Sera of Patients with Active Pulmonary Tuberculosis. PLoS ONE 2013, 8, e80149. [Google Scholar] [CrossRef] [Green Version]
- Verhagen, L.M.; Zomer, A.; Maes, M.; Villalba, J.A.; del Nogal, B.; Eleveld, M.; van Hijum, S.A.F.T.; de Waard, J.H.; Hermans, P.W.M. A predictive signature gene set for discriminating active from latent tuberculosis in Warao Amerindian children. BMC Genom. 2013, 14, 74. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Yu, G.; Yang, X.; Zhu, C.; Zhang, Z.; Zhan, X. Circulating microRNAs as biomarkers for the early diagnosis of childhood tuberculosis infection. Mol. Med. Rep. 2016, 13, 4620–4626. [Google Scholar] [CrossRef] [Green Version]
- Jenum, S.; Dhanasekaran, S.; Lodha, R.; Mukherjee, A.; Kumar Saini, D.; Singh, S.; Singh, V.; Medigeshi, G.; Haks, M.C.; Ottenhoff, T.H.M.; et al. Approaching a diagnostic point-of-care test for pediatric tuberculosis through evaluation of immune biomarkers across the clinical disease spectrum. Sci. Rep. 2016, 6, 18520. [Google Scholar] [CrossRef]
- Yong, Y.K.; Tan, H.Y.; Saeidi, A.; Wong, W.F.; Vignesh, R.; Velu, V.; Eri, R.; Larsson, M.; Shankar, E.M. Immune Biomarkers for Diagnosis and Treatment Monitoring of Tuberculosis: Current Developments and Future Prospects. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Schubert, O.T.; Mouritsen, J.; Ludwig, C.; Röst, H.L.; Rosenberger, G.; Arthur, P.K.; Claassen, M.; Campbell, D.S.; Sun, Z.; Farrah, T.; et al. The Mtb Proteome Library: A Resource of Assays to Quantify the Complete Proteome of Mycobacterium tuberculosis. Cell Host Microbe 2013, 13, 602–612. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.H.; Chen, Y.Q.; Sun, Y.; Liu, Y.; Zheng, S.H.; Zhnag, Z.G.; Li, C.Y. Screening of Serum Biomarkers for Distinguishing between Latent and Active Tuberculosis Using Proteome Microarray. Biomed. Env. Sci. 2018, 31, 515–526. [Google Scholar] [CrossRef]
- Cao, T.; Lyu, L.; Jia, H.; Wang, J.; Du, F.; Pan, L.; Li, Z.; Xing, A.; Xiao, J.; Ma, Y.; et al. A Two-Way Proteome Microarray Strategy to Identify Novel Mycobacterium tuberculosis-Human Interactors. Front. Cell. Infect. Microbiol. 2019, 9, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, J.; Bi, L.; Zhou, L.; Guo, S.; Fleming, J.; Jiang, H.; Zhou, Y.; Gu, J.; Zhong, Q.; Wang, Z.; et al. Mycobacterium Tuberculosis Proteome Microarray for Global Studies of Protein Function and Immunogenicity. Cell Rep. 2014, 9, 2317–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Sun, L.; Xu, F.; Xiao, J.; Jiao, W.; Qi, H.; Shen, C.; Shen, A. Characterization of plasma proteins in children of different Mycobacterium tuberculosis infection status using label-free quantitative proteomics. Oncotarget 2017, 8, 103290–103301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layre, E.; Sweet, L.; Hong, S.; Madigan, C.A.; Desjardins, D.; Young, D.C.; Cheng, T.-Y.; Annand, J.W.; Kim, K.; Shamputa, I.C.; et al. A Comparative Lipidomics Platform for Chemotaxonomic Analysis of Mycobacterium tuberculosis. Chem. Biol. 2011, 18, 1537–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shui, G.; Bendt, A.K.; Jappar, I.A.; Lim, H.M.; Laneelle, M.; Hervé, M.; Via, L.E.; Chua, G.H.; Bratschi, M.W.; Zainul Rahim, S.Z.; et al. Mycolic acids as diagnostic markers for tuberculosis case detection in humans and drug efficacy in mice. EMBO Mol. Med. 2012, 4, 27–37. [Google Scholar] [CrossRef]
- Andreas, N.J.; Basu Roy, R.; Gomez-Romero, M.; Horneffer-van der Sluis, V.; Lewis, M.R.; Camuzeaux, S.S.M.; Jiménez, B.; Posma, J.M.; Tientcheu, L.; Egere, U.; et al. Performance of metabonomic serum analysis for diagnostics in paediatric tuberculosis. Sci. Rep. 2020, 10, 7302. [Google Scholar] [CrossRef]
- Zhou, A.; Ni, J.; Xu, Z.; Wang, Y.; Lu, S.; Sha, W.; Karakousis, P.C.; Yao, Y.-F. Application of 1 H NMR Spectroscopy-Based Metabolomics to Sera of Tuberculosis Patients. J. Proteome Res. 2013, 12, 4642–4649. [Google Scholar] [CrossRef] [Green Version]
- Frediani, J.K.; Jones, D.P.; Tukvadze, N.; Uppal, K.; Sanikidze, E.; Kipiani, M.; Tran, V.T.; Hebbar, G.; Walker, D.I.; Kempker, R.R.; et al. Plasma metabolomics in human pulmonary tuberculosis disease: A pilot study. PLoS ONE 2014, 9, e108854. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Lee, K.C.; Curreem, S.O.T.; Chow, W.N.; To, K.K.W.; Hung, I.F.N.; Ho, D.T.Y.; Sridhar, S.; Li, I.W.S.; Ding, V.S.Y.; et al. Metabolomic profiling of plasma from patients with tuberculosis by use of untargeted mass spectrometry reveals novel biomarkers for diagnosis. J. Clin. Microbiol. 2015, 53, 3750–3759. [Google Scholar] [CrossRef] [Green Version]
- Kato-Maeda, M.; Ho, C.; Passarelli, B.; Banaei, N.; Grinsdale, J.; Flores, L.; Anderson, J.; Murray, M.; Rose, G.; Kawamura, L.M.; et al. Use of Whole Genome Sequencing to Determine the Microevolution of Mycobacterium tuberculosis during an Outbreak. PLoS ONE 2013, 8, e58235. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Assunção, J.; Crampin, A.; Houben, R.; Mzembe, T.; Mallard, K.; Coll, F.; Khan, P.; Banda, L.; Chiwaya, A.; Pereira, R.; et al. Large-scale whole genome sequencing of M. tuberculosis provides insights into transmission in a high prevalence area. Elife 2015, 4, e05166. [Google Scholar] [CrossRef] [PubMed]
- Zetola, N.M.; Shin, S.S.; Tumedi, K.A.; Moeti, K.; Ncube, R.; Nicol, M.; Collman, R.G.; Klausner, J.D.; Modongo, C. Mixed Mycobacterium tuberculosis Complex Infections and False-Negative Results for Rifampin Resistance by GeneXpert MTB/RIF Are Associated with Poor Clinical Outcomes. J. Clin. Microbiol. 2014, 52, 2422–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, P.; Wang, X.; Shen, X.; Shi, M.; Zhu, X.; Yu, X.; Liu, J.; Ling, C.; Wu, M. Use of DNA microarray chips for the rapid detection of Mycobacterium tuberculosis resistance to rifampicin and isoniazid. Exp. Ther. Med. 2017, 13, 2332–2338. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Weng, J.-Y.; Huang, H.-H.; Yen, W.-C.; Tsai, Y.-H.; Cheng, T.C.; Jou, R. A new oligonucleotide array for the detection of multidrug and extensively drug-resistance tuberculosis. Sci. Rep. 2019, 9, 4425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iketleng, T.; Lessells, R.; Dlamini, M.T.; Mogashoa, T.; Mupfumi, L.; Moyo, S.; Gaseitsiwe, S.; de Oliveira, T. Mycobacterium tuberculosis Next-Generation Whole Genome Sequencing: Opportunities and Challenges. Tuberc. Res. Treat. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gardy, J.L.; Johnston, J.C.; Sui, S.J.H.; Cook, V.J.; Shah, L.; Brodkin, E.; Rempel, S.; Moore, R.; Zhao, Y.; Holt, R.; et al. Whole-Genome Sequencing and Social-Network Analysis of a Tuberculosis Outbreak. N. Engl. J. Med. 2011, 364, 730–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Tagle, C.; Naves, R.; Balcells, M.E. Unraveling the Role of MicroRNAs in Mycobacterium tuberculosis Infection and Disease: Advances and Pitfalls. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, Z.; Wei, W.; Liu, Z.; Fleming, J.; Zhang, S.; Lin, N.; Wang, M.; Chen, M.; Xu, Y.; et al. Identification of Serum microRNA Biomarkers for Tuberculosis Using RNA-seq. PLoS ONE 2014, 9, e88909. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, L.S.; Ribeiro-Alves, M.; Leal-Calvo, T.; Leung, J.; Durán, V.; Samir, M.; Talbot, S.; Tallam, A.; de Mello, F.C.Q.; Geffers, R.; et al. Reprogramming of Small Noncoding RNA Populations in Peripheral Blood Reveals Host Biomarkers for Latent and Active Mycobacterium tuberculosis Infection. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.T.; Kaforou, M.; Brent, A.J.; Wright, V.J.; Banwell, C.M.; Chagaluka, G.; Crampin, A.C.; Dockrell, H.M.; French, N.; Hamilton, M.S.; et al. Diagnosis of Childhood Tuberculosis and Host RNA Expression in Africa. N. Engl. J. Med. 2014, 370, 1712–1723. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, T.E.; Braviak, L.; Tato, C.M.; Khatri, P. Genome-wide expression for diagnosis of pulmonary tuberculosis: A multicohort analysis. Lancet Respir. Med. 2016, 4, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Tornheim, J.A.; Madugundu, A.K.; Paradkar, M.; Fukutani, K.F.; Queiroz, A.T.L.; Gupte, N.; Gupte, A.N.; Kinikar, A.; Kulkarni, V.; Balasubramanian, U.; et al. Transcriptomic Profiles of Confirmed Pediatric Tuberculosis Patients and Household Contacts Identifies Active Tuberculosis, Infection, and Treatment Response Among Indian Children. J. Infect. Dis. 2020, 221, 1647–1658. [Google Scholar] [CrossRef]
- Gjøen, J.E.; Jenum, S.; Sivakumaran, D.; Mukherjee, A.; Macaden, R.; Kabra, S.K.; Lodha, R.; Ottenhoff, T.H.M.; Haks, M.C.; Doherty, T.M.; et al. Novel transcriptional signatures for sputum-independent diagnostics of tuberculosis in children. Sci. Rep. 2017, 7, 5839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergne, I.; Chua, J.; Singh, S.B.; Deretic, V. Cell Biology of Mycobacterium tuberculosis Phagosome. Annu. Rev. Cell Dev. Biol. 2004, 20, 367–394. [Google Scholar] [CrossRef] [PubMed]
- Dyer, M.D.; Neff, C.; Dufford, M.; Rivera, C.G.; Shattuck, D.; Bassaganya-Riera, J.; Murali, T.M.; Sobral, B.W. The Human-Bacterial Pathogen Protein Interaction Networks of Bacillus anthracis, Francisella tularensis, and Yersinia pestis. PLoS ONE 2010, 5, e12089. [Google Scholar] [CrossRef] [Green Version]
- König, R.; Zhou, Y.; Elleder, D.; Diamond, T.L.; Bonamy, G.M.C.; Irelan, J.T.; Chiang, C.; Tu, B.P.; De Jesus, P.D.; Lilley, C.E.; et al. Global Analysis of Host-Pathogen Interactions that Regulate Early-Stage HIV-1 Replication. Cell 2008, 135, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunnath-Velayudhan, S.; Porcelli, S.A. Recent Advances in Defining the Immunoproteome of Mycobacterium tuberculosis. Front. Immunol. 2013, 4, 335. [Google Scholar] [CrossRef] [Green Version]
- Kunnath-Velayudhan, S.; Salamon, H.; Wang, H.-Y.; Davidow, A.L.; Molina, D.M.; Huynh, V.T.; Cirillo, D.M.; Michel, G.; Talbot, E.A.; Perkins, M.D.; et al. Dynamic antibody responses to the Mycobacterium tuberculosis proteome. Proc. Natl. Acad. Sci. USA 2010, 107, 14703–14708. [Google Scholar] [CrossRef] [Green Version]
- Britton, W.J.; Hellqvist, L.; Ivanyi, J.; Basten, A. Immunopurification of Radiolabelled Antigens of Mycobacterium leprae and Mycobacterium bovis (Bacillus Calmette-Guerin) with Monoclonal Antibodies. Scand. J. Immunol. 1987, 26, 149–159. [Google Scholar] [CrossRef]
- Daugelat, S.; Guile, H.; Schoel, B.; Kaufmann, S.H.E. Secreted Antigens of Mycobacterium tuberculosis: Characterization with T Lymphocytes from Patients and Contacts after Two-Dimensional Separation. J. Infect. Dis. 1992, 166, 186–190. [Google Scholar] [CrossRef]
- Wong, D.K.; Lee, B.-Y.; Horwitz, M.A.; Gibson, B.W. Identification of Fur, Aconitase, and Other Proteins Expressed by Mycobacterium tuberculosis under Conditions of Low and High Concentrations of Iron by Combined Two-Dimensional Gel Electrophoresis and Mass Spectrometry. Infect. Immun. 1999, 67, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollenkopf, H.-J.; Jungblut, P.R.; Raupach, B.; Mattow, J.; Lamer, S.; Zimny-Arndt, U.; Schaible, U.E.; Kaufmann, S.H.E. A dynamic two-dimensional polyacrylamide gel electrophoresis database: The mycobacterial proteomevia Internet. Electrophoresis 1999, 20, 2172–2180. [Google Scholar] [CrossRef]
- Kelkar, D.S.; Kumar, D.; Kumar, P.; Balakrishnan, L.; Muthusamy, B.; Yadav, A.K.; Shrivastava, P.; Marimuthu, A.; Anand, S.; Sundaram, H.; et al. Proteogenomic Analysis of Mycobacterium tuberculosis By High Resolution Mass Spectrometry. Mol. Cell. Proteomics 2011, 10, M111.011627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-S.; Korobkova, E.; Chen, H.; Zhu, J.; Jian, X.; Tao, S.-C.; He, C.; Zhu, H. A proteome chip approach reveals new DNA damage recognition activities in Escherichia coli. Nat. Methods. 2008, 5, 69–74. [Google Scholar] [CrossRef]
- Zhu, H. Global Analysis of Protein Activities Using Proteome Chips. Science 2001, 293, 2101–2105. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, J.; Shi, J.; Wang, M.; Rao, M.; Xue, C.; Du, Y.; He, Z.-G. A Proteome-Scale Identification of Novel Antigenic Proteins in Mycobacterium tuberculosis toward Diagnostic and Vaccine Development. J. Proteome Res. 2010, 9, 4812–4822. [Google Scholar] [CrossRef]
- Sette, A.; Moutaftsi, M.; Moyron-Quiroz, J.; McCausland, M.M.; Davies, D.H.; Johnston, R.J.; Peters, B.; Rafii-El-Idrissi Benhnia, M.; Hoffmann, J.; Su, H.-P.; et al. Selective CD4+ T Cell Help for Antibody Responses to a Large Viral Pathogen: Deterministic Linkage of Specificities. Immunity 2008, 28, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Penn-Nicholson, A.; Hraha, T.; Thompson, E.G.; Sterling, D.; Mbandi, S.K.; Wall, K.M.; Fisher, M.; Suliman, S.; Shankar, S.; Hanekom, W.A.; et al. Discovery and validation of a prognostic proteomic signature for tuberculosis progression: A prospective cohort study. PLOS Med. 2019, 16, e1002781. [Google Scholar] [CrossRef]
- Perkowski, E.F.; Zulauf, K.E.; Weerakoon, D.; Hayden, J.D.; Ioerger, T.R.; Oreper, D.; Gomez, S.M.; Sacchettini, J.C.; Braunstein, M. The EXIT Strategy: An Approach for Identifying Bacterial Proteins Exported during Host Infection. MBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Chen, Q.; Zhang, M.; Cai, Y.; Yang, F.; Zhang, J.; Deng, G.; Ye, T.; Deng, Q.; Li, G.; et al. Identification of eight-protein biosignature for diagnosis of tuberculosis. Thorax 2020, 75, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Kunnath-Velayudhan, S.; Gennaro, M.L. Immunodiagnosis of Tuberculosis: A Dynamic View of Biomarker Discovery. Clin. Microbiol. Rev. 2011, 24, 792–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, P. Structure, function, and biogenesis of the cell wall of Mycobacterium tuberculosis. Tuberculosis 2003, 83, 91–97. [Google Scholar] [CrossRef]
- Pal, R.; Hameed, S.; Kumar, P.; Singh, S.; Fatima, Z. Comparative lipidomics of drug sensitive and resistant Mycobacterium tuberculosis reveals altered lipid imprints. 3 Biotech. 2017, 7, 325. [Google Scholar] [CrossRef] [PubMed]
- Raghunandanan, S.; Jose, L.; Gopinath, V.; Kumar, R.A. Comparative label-free lipidomic analysis of Mycobacterium tuberculosis during dormancy and reactivation. Sci. Rep. 2019, 9, 3660. [Google Scholar] [CrossRef] [PubMed]
- Korf, J.; Stoltz, A.; Verschoor, J.; De Baetselier, P.; Grooten, J. The Mycobacterium tuberculosis cell wall component mycolic acid elicits pathogen-associated host innate immune responses. Eur. J. Immunol. 2005, 35, 890–900. [Google Scholar] [CrossRef]
- Marrakchi, H.; Lanéelle, M.-A.; Daffé, M. Mycolic Acids: Structures, Biosynthesis, and Beyond. Chem. Biol. 2014, 21, 67–85. [Google Scholar] [CrossRef] [Green Version]
- Nataraj, V.; Varela, C.; Javid, A.; Singh, A.; Besra, G.S.; Bhatt, A. Mycolic acids: Deciphering and targeting the Achilles’ heel of the tubercle bacillus. Mol. Microbiol. 2015, 98, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Aoyagi, Y.; Mitome, H.; Fujita, T.; Naoki, H.; Ridell, M.; Minnikin, D.E. Location of functional groups in mycobacterial meromycolate chains; the recognition of new structural principles in mycolic acids. Microbiology 2002, 148, 1881–1902. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, N.; Takayama, K.; Jordi, H.C.; Schnoes, H.K. Characterization of the purified components of a new homologous series of α-mycolic acids from Mycobacterium tuberculosis H37Ra. J. Biol. Chem. 1978, 253, 5411–5417. [Google Scholar]
- Perumal, J.; Dinish, U.; Bendt, A.; Kazakeviciute, A.; Fu, C.Y.; Ong, I.L.H.; Olivo, M. Identification of mycolic acid forms using surface-enhanced Raman scattering as a fast detection method for tuberculosis. Int. J. Nanomed. 2018, 13, 6029–6038. [Google Scholar] [CrossRef] [Green Version]
- Ndlandla, F.L.; Ejoh, V.; Stoltz, A.C.; Naicker, B.; Cromarty, A.D.; van Wyngaardt, S.; Khati, M.; Rotherham, L.S.; Lemmer, Y.; Niebuhr, J.; et al. Standardization of natural mycolic acid antigen composition and production for use in biomarker antibody detection to diagnose active tuberculosis. J. Immunol. Methods 2016, 435, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Thanyani, S.T.; Roberts, V.; Siko, D.G.R.; Vrey, P.; Verschoor, J.A. A novel application of affinity biosensor technology to detect antibodies to mycolic acid in tuberculosis patients. J. Immunol. Methods 2008, 332, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druszczynska, M.; Wawrocki, S.; Szewczyk, R.; Rudnicka, W. Mycobacteria-derived biomarkers for tuberculosis diagnosis. Indian J. Med. Res. 2017, 146, 700. [Google Scholar] [CrossRef]
- Layre, E.; Al-Mubarak, R.; Belisle, J.T.; Branch Moody, D. Mycobacterial Lipidomics. Microbiol. Spectr. 2014, 2, 341–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Khoo, K.H. Mycobacterial lipoarabinomannan: An extraordinary lipoheteroglycan with profound physiological effects. Glycobiology 1998, 8, 113–120. [Google Scholar] [CrossRef]
- Lawn, S.D. Point-of-care detection of lipoarabinomannan (LAM) in urine for diagnosis of HIV-associated tuberculosis: A state of the art review. BMC Infect. Dis. 2012, 12, 103. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.J.; Diels, O.; Fischer, F.G.; Pauling, L.; Siedel, W.; Anderson, R.J. The Chemistry of the Lipoids of the Tubercle Bacillus and certain other Microorganisms. In Fortschritte der Chemie Organischer Naturstoffe; Springer: Berlin/Heidelberg, Germany, 1939. [Google Scholar]
- Astarie-Dequeker, C.; Nigou, J.; Passemar, C.; Guilhot, C. The role of mycobacterial lipids in host pathogenesis. Drug Discov. Today 2010, 7, e33–e41. [Google Scholar] [CrossRef]
- Kurz, S.G.; Rivas-Santiago, B. Time to Expand the Picture of Mycobacterial Lipids: Spotlight on Nontuberculous Mycobacteria. Am. J. Respir. Cell Mol. Biol. 2020, 62, 275–276. [Google Scholar] [CrossRef]
- Hunter, S.W.; Brennan, P.J. Evidence for the presence of a phosphatidylinositol anchor on the lipoarabinomannan and lipomannan of Mycobacterium tuberculosis. J. Biol. Chem. 1990, 265, 9272–9279. [Google Scholar]
- Chatterjee, D.; Lowell, K.; Rivoire, B.; McNeil, M.R.; Brennan, P.J. Lipoarabinomannan of Mycobacterium tuberculosis. Capping with mannosyl residues in some strains. J. Biol. Chem. 1992, 267, 6234–6239. [Google Scholar]
- Peter, J.G.; Cashmore, T.J.; Meldau, R.; Theron, G.; van Zyl-Smit, R.; Dheda, K. Diagnostic accuracy of induced sputum LAM ELISA for tuberculosis diagnosis in sputum-scarce patients. Int. J. Tuberc. Lung Dis. 2012, 16, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Hamasur, B.; Bruchfeld, J.; Haile, M.; Pawlowski, A.; Bjorvatn, B.; Källenius, G.; Svenson, S.B. Rapid diagnosis of tuberculosis by detection of mycobacterial lipoarabinomannan in urine. J. Microbiol. Methods 2001, 45, 41–52. [Google Scholar] [CrossRef]
- Sada, E.; Aguilar, D.; Torres, M.; Herrera, T. Detection of lipoarabinomannan as a diagnostic test for tuberculosis. J. Clin. Microbiol. 1992, 30, 2415–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia-Neves, M.; Fröberg, G.; Korshun, L.; Viegas, S.; Vaz, P.; Ramanlal, N.; Bruchfeld, J.; Hamasur, B.; Brennan, P.; Källenius, G. Biomarkers for tuberculosis: The case for lipoarabinomannan. ERJ Open Res. 2019, 5, 00115–02018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connell, T.G.; Ritz, N.; Paxton, G.A.; Buttery, J.P.; Curtis, N.; Ranganathan, S.C. A Three-Way Comparison of Tuberculin Skin Testing, QuantiFERON-TB Gold and T-SPOT.TB in Children. PLoS ONE 2008, 3, e2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroidl, I.; Clowes, P.; Reither, K.; Mtafya, B.; Rojas-Ponce, G.; Ntinginya, E.N.; Kalomo, M.; Minja, L.T.; Kowuor, D.; Saathoff, E.; et al. Performance of urine lipoarabinomannan assays for paediatric tuberculosis in Tanzania. Eur. Respir. J. 2015, 46, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Stromberg, L.R.; Mendez, H.M.; Kubicek-Sutherland, J.Z.; Graves, S.W.; Hengartner, N.W.; Mukundan, H. Presentation matters: Impact of association of amphiphilic LPS with serum carrier proteins on innate immune signaling. PLoS ONE 2018, 13, e0198531. [Google Scholar] [CrossRef]
- Vu, D.M.; Sakamuri, R.M.; Waters, W.R.; Swanson, B.I.; Mukundan, H. Detection of Lipomannan in Cattle Infected with Bovine Tuberculosis. Anal. Sci. 2017, 33, 457–460. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Matsumura, T.; Ato, M.; Hamasaki, M.; Nishiuchi, Y.; Murakami, Y.; Maeda, Y.; Yoshimori, T.; Matsumoto, S.; Kobayashi, K.; et al. Critical Roles for Lipomannan and Lipoarabinomannan in Cell Wall Integrity of Mycobacteria and Pathogenesis of Tuberculosis. MBio 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Buddle, B.; Lisle, G.; Waters, W.; Vordermeier, H. Diagnosis of mycobacterium bovis infection in cattle. In Tuberculosis, Leprosy and Mycobacterial Diseases of Man and Animals: The Many Hosts of Mycobacteria; Mukundan, H., Chambers, M., Waters, W., Larsen, M., Eds.; CAB International: Boston, MA, USA, 2015; pp. 168–184. [Google Scholar]
- Kubicek-Sutherland, J.Z.; Vu, D.M.; Noormohamed, A.; Mendez, H.M.; Stromberg, L.R.; Pedersen, C.A.; Hengartner, A.C.; Klosterman, K.E.; Bridgewater, H.A.; Otieno, V.; et al. Direct detection of bacteremia by exploiting host-pathogen interactions of lipoteichoic acid and lipopolysaccharide. Sci. Rep. 2019, 9, 6203. [Google Scholar] [CrossRef] [Green Version]
- Mukundan, H.; Kubicek-Sutherland, J.Z.; Jakhar, S.; Noormohamed, A.; Sakamuri, R.; Swanson, B.I.; Stromberg, L.R. Immunoassays for the Differentiation of Bacterial Pathogens in Human Serum. U.S. Patent Application No. 15/640,865, 26 October 2017. [Google Scholar]
- Kubicek-Sutherland, J.Z.; Vu, D.M.; Mendez, H.M.; Jakhar, S.; Mukundan, H. Detection of lipid and amphiphilic biomarkers for disease diagnostics. Biosensors 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noormohamed, A.; Stromberg, L.R.; Anderson, A.S.; Karim, Z.; Dighe, P.; Kempaiah, P.; Ong’echa, J.M.; Perkins, D.J.; Doggett, N.; McMahon, B.; et al. Detection of lipopolysaccharides in serum using a waveguide-based optical biosensor. In Optical Diagnostics and Sensing XVII: Toward Point-of-Care Diagnostics; Coté, G.L., Ed.; SPIE: Bellingham, WA, USA, 2017; p. 100720A. [Google Scholar]
- Anes, E.; Kühnel, M.P.; Bos, E.; Moniz-Pereira, J.; Habermann, A.; Griffiths, G. Selected lipids activate phagosome actin assembly and maturation resulting in killing of pathogenic mycobacteria. Nat. Cell Biol. 2003, 5, 793–802. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakhar, S.; Bitzer, A.A.; Stromberg, L.R.; Mukundan, H. Pediatric Tuberculosis: The Impact of “Omics” on Diagnostics Development. Int. J. Mol. Sci. 2020, 21, 6979. https://doi.org/10.3390/ijms21196979
Jakhar S, Bitzer AA, Stromberg LR, Mukundan H. Pediatric Tuberculosis: The Impact of “Omics” on Diagnostics Development. International Journal of Molecular Sciences. 2020; 21(19):6979. https://doi.org/10.3390/ijms21196979
Chicago/Turabian StyleJakhar, Shailja, Alexis A. Bitzer, Loreen R. Stromberg, and Harshini Mukundan. 2020. "Pediatric Tuberculosis: The Impact of “Omics” on Diagnostics Development" International Journal of Molecular Sciences 21, no. 19: 6979. https://doi.org/10.3390/ijms21196979
APA StyleJakhar, S., Bitzer, A. A., Stromberg, L. R., & Mukundan, H. (2020). Pediatric Tuberculosis: The Impact of “Omics” on Diagnostics Development. International Journal of Molecular Sciences, 21(19), 6979. https://doi.org/10.3390/ijms21196979