Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
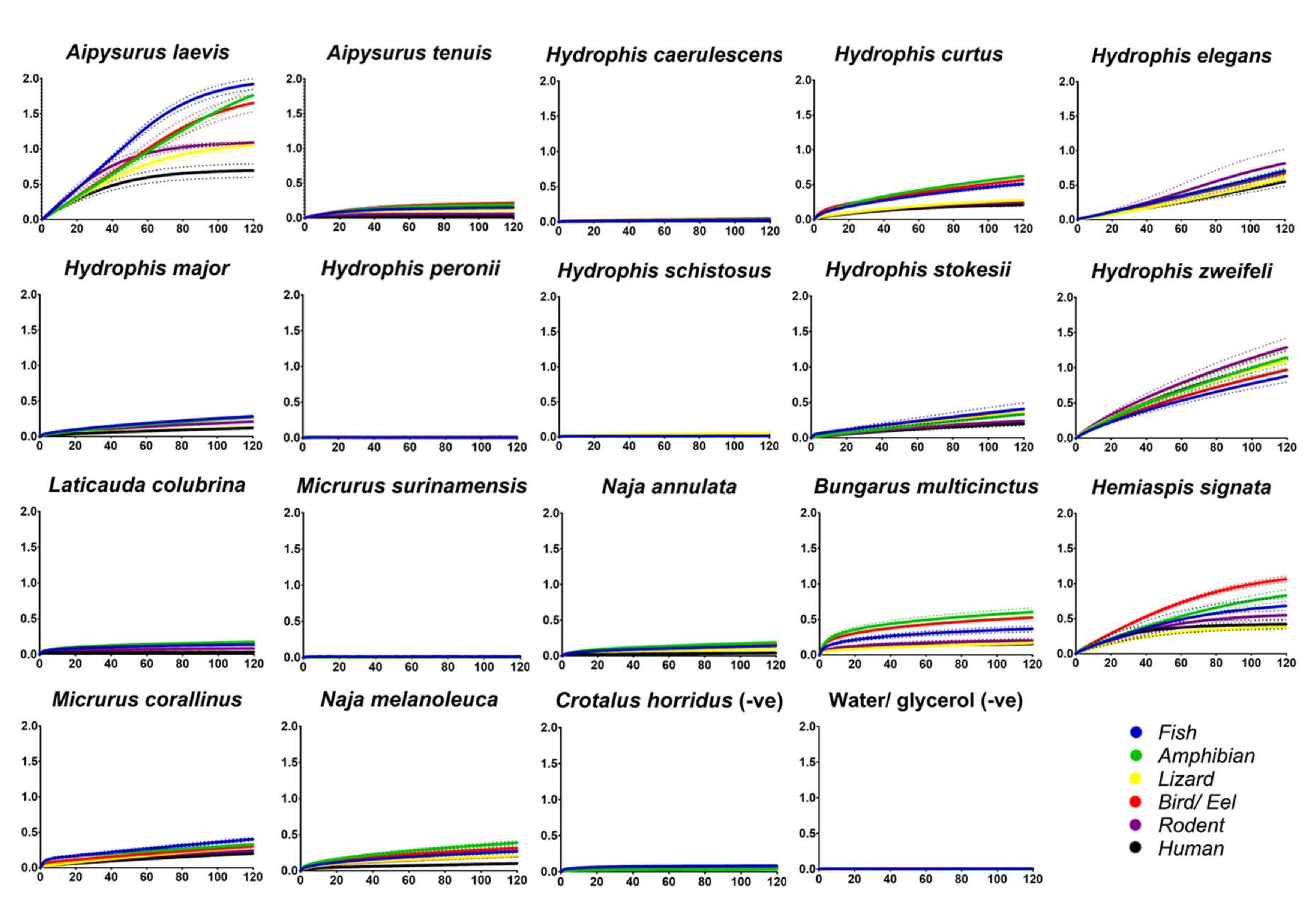
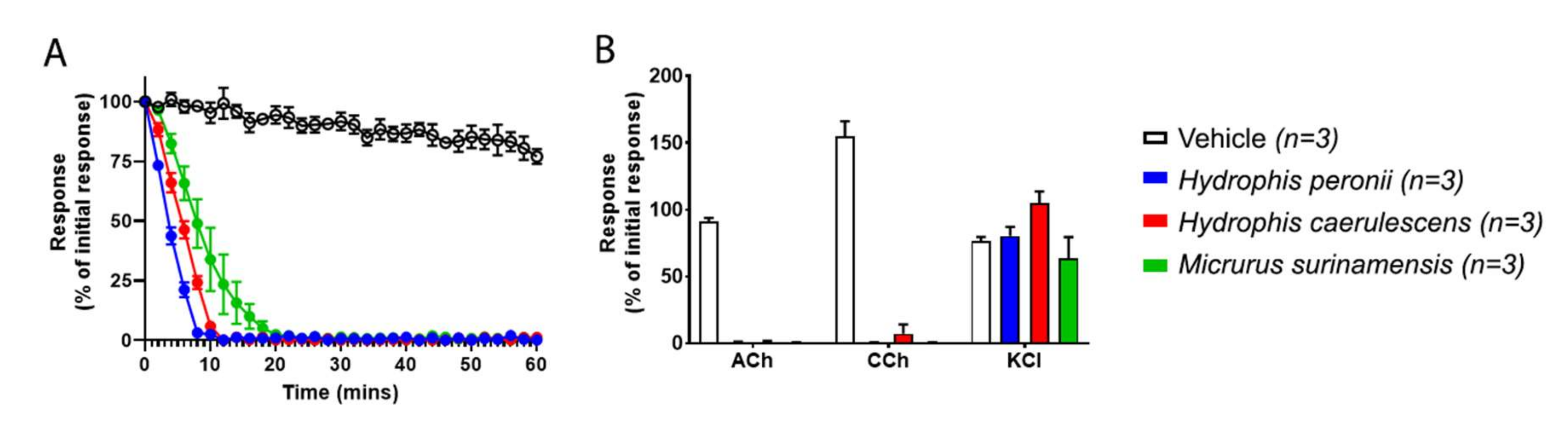
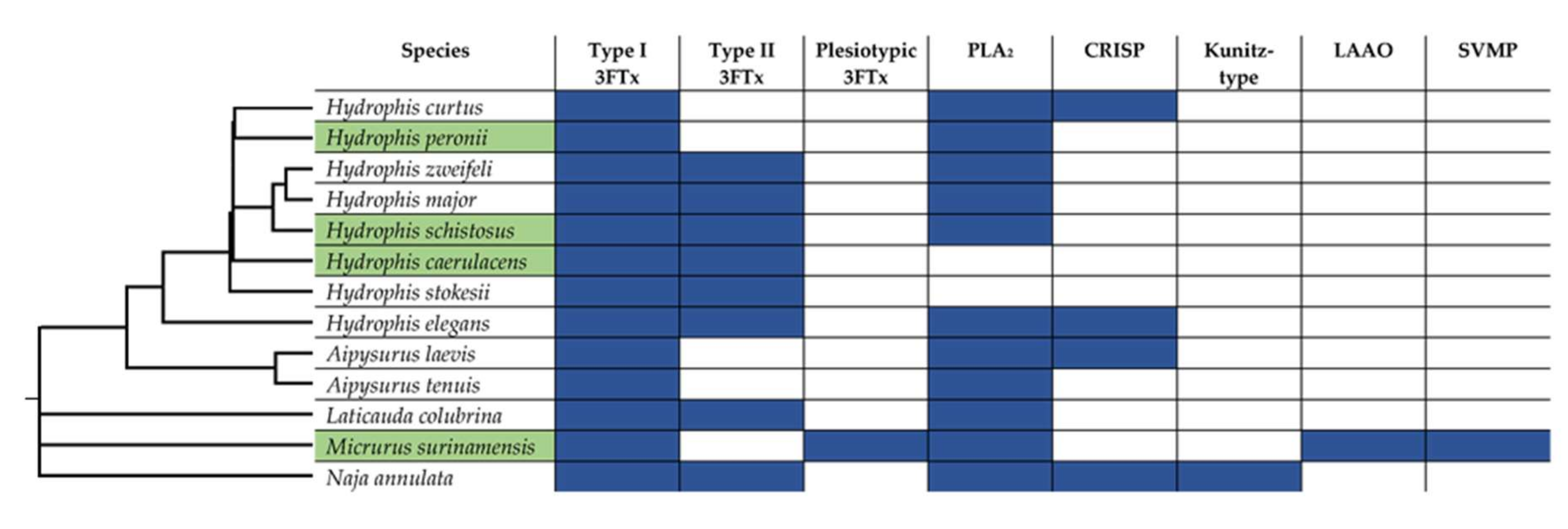
2. Results and Discussion
3. Materials and Methods
3.1. Venom Collection and Preparation
3.2. Mimotope Production and Preparation
3.3. Biolayer Interferometry (BLI)
3.4. Sample Preparation for Mass Spectrometry (LC-MS/MS)
3.5. Mass Spectrometry (LC-MS/MS)
3.6. Isolated Chick-Biventer Cervicis Nerve-Muscle Preparation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| nAChR | Nicotinic acetylcholine receptor |
| mAChR | Muscarinic acetylcholine receptor |
| 3FTx | Three-finger toxin |
| PLA2 | Phospholipase A2 |
| CRISP | Cystine -rich secretory protein |
| Kunitz-type | Kunitz-type protease inhibitor |
| LAAO | L-Amino acid oxidase |
| SVMP | Snake venom metalloprotease |
| SVSP | Snake venom serine protease |
| BLI | Biolayer interferometry |
| CBCNM | Chick-biventer cervicis nerve muscle |
| ACh | Acetylcholine |
| CCh | Carbochol |
| KCl | Potassium chloride |
References
- Young, B. The influences of the aquatic medium on the prey capture system of snakes. J. Nat. Hist. 1991, 25, 519–531. [Google Scholar] [CrossRef]
- Shine, R. Constraints on reproductive investment: A comparison between aquatic and terrestrial snakes. Evolution 1988, 42, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnakone, P.; Kochva, E. Venom glands and some associated muscles in sea snakes. J. Morphol. 1990, 205, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Sanders, K.L.; King, B.; Palci, A. Diversification rates and phenotypic evolution in venomous snakes (Elapidae). R. Soc. Open Sci. 2016, 3, 150277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Fry, B.; Kini, R.M. Eggs-only diet: Its implications for the toxin profile changes and ecology of the marbled sea snake (Aipysurus eydouxii). J. Mol. Evol. 2005, 60, 81–89. [Google Scholar] [CrossRef]
- Glodek, G.S.; Voris, H.K. Marine snake diets: Prey composition, diversity and overlap. Copeia 1982, 661–666. [Google Scholar] [CrossRef]
- Voris, H.K.; Voris, H.H. Feeding strategies in marine snakes: An analysis of evolutionary, morphological, behavioral and ecological relationships. Am. Zool. 1983, 23, 411–425. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Fry, B.G.; Kini, R.M. Putting the brakes on snake venom evolution: The unique molecular evolutionary patterns of Aipysurus eydouxii (Marbled sea snake) phospholipase A2 toxins. Mol. Biol. Evol. 2005, 22, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Chetty, N.; Du, A.; Hodgson, W.C.; Winkel, K.; Fry, B.G. The in vitro neuromuscular activity of Indo-Pacific sea-snake venoms: Efficacy of two commercially available antivenoms. Toxicon 2004, 44, 193–200. [Google Scholar] [CrossRef]
- Kishida, T.; Hikida, T. Degeneration patterns of the olfactory receptor genes in sea snakes. J. Evol. Biol. 2010, 23, 302–310. [Google Scholar] [CrossRef]
- Doolittle, R.F. Step-by-step evolution of vertebrate blood coagulation. Proc. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerenius, L.; Söderhäll, K. Coagulation in invertebrates. J. Innate Immun. 2011, 3, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Nirthanan, S.; Gwee, M.C. Three-finger α-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chang, C. The action of snake venoms on nerve and muscle. In Snake Venom; Springer: Berlin/Heidelberg, Germany, 1979; pp. 309–376. [Google Scholar]
- Servent, D.; Ménez, A. Snake neurotoxins that interact with nicotinic acetylcholine receptors. In Handbook of Neurotoxicology; Springer: Berlin/Heidelberg, Germany, 2002; pp. 38–425. [Google Scholar]
- Dellisanti, C.; Yao, Y.; Stroud, J.C.; Wang, Z.-Z.; Chen, L. Structural determinants for α-neurotoxin sensitivity in muscle nAChR and their implications for the gating mechanism. Channels 2007, 1, 234–237. [Google Scholar] [CrossRef] [Green Version]
- Dellisanti, C.D.; Yao, Y.; Stroud, J.C.; Wang, Z.-Z.; Chen, L. Crystal structure of the extracellular domain of nAChR α1 bound to α-bungarotoxin at 1.94 Å resolution. Nat. Neurosci. 2007, 10, 953–962. [Google Scholar] [CrossRef]
- Kachalsky, S.G.; Jensen, B.S.; Barchan, D.; Fuchs, S. Two subsites in the binding domain of the acetylcholine receptor: An aromatic subsite and a proline subsite. Proc. Natl. Acad. Sci. USA 1995, 92, 10801–10805. [Google Scholar] [CrossRef] [Green Version]
- Tzartos, S.; Remoundos, M.S. Fine localization of the major alpha-bungarotoxin binding site to residues alpha 189-195 of the Torpedo acetylcholine receptor. Residues 189, 190, and 195 are indispensable for binding. J. Biol. Chem. 1990, 265, 21462–21467. [Google Scholar]
- Bracci, L.; Lozzi, L.; Lelli, B.; Pini, A.; Neri, P. Mimotopes of the nicotinic receptor binding site selected by a combinatorial peptide library. Biochemistry 2001, 40, 6611–6619. [Google Scholar] [CrossRef]
- Gotti, C.; Clementi, F. Neuronal nicotinic receptors: From structure to pathology. Prog. Neurobiol. 2004, 74, 363–396. [Google Scholar] [CrossRef]
- Fry, B.G.; Lumsden, N.G.; Wüster, W.; Wickramaratna, J.C.; Hodgson, W.C.; Kini, R.M. Isolation of a neurotoxin (α-colubritoxin) from a nonvenomous colubrid: Evidence for early origin of venom in snakes. J. Mol. Evol. 2003, 57, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the Green Vinesnake (Oxybelis fulgidus; family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Mackessy, S.P.; Fry, B.G.; Bhatia, M.; Mourier, G.; Fruchart-Gaillard, C.; Servent, D.; Ménez, R.; Stura, E.; Ménez, A. Denmotoxin, a three-finger toxin from the colubrid snake Boiga dendrophila (Mangrove Catsnake) with bird-specific activity. J. Biol. Chem. 2006, 281, 29030–29041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlak, J.; Mackessy, S.P.; Sixberry, N.M.; Stura, E.A.; Le Du, M.H.; Ménez, R.; Foo, C.S.; Ménez, A.; Nirthanan, S.; Kini, R.M. Irditoxin, a novel covalently linked heterodimeric three-finger toxin with high taxon-specific neurotoxicity. FASEB J. 2009, 23, 534–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fry, B.G.; Scheib, H.; van der Weerd, L.; Young, B.; McNaughtan, J.; Ramjan, S.R.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an arsenal: Structural and functional diversification of the venom system in the advanced snakes (Caenophidia). Mol. Cell. Proteom. 2008, 7, 215–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashevsky, D.; Fry, B.G. Ancient diversification of three-finger toxins in Micrurus coral snakes. J. Mol. Evol. 2018, 86, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Quinton, L.; Girard, E.; Maiga, A.; Rekik, M.; Lluel, P.; Masuyer, G.; Larregola, M.; Marquer, C.; Ciolek, J.; Magnin, T. Isolation and pharmacological characterization of AdTx1, a natural peptide displaying specific insurmountable antagonism of the α1A--adrenoceptor. Br. J. Pharmacol. 2010, 159, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Rouget, C.; Quinton, L.; Maïga, A.; Gales, C.; Masuyer, G.; Malosse, C.; Chamot--Rooke, J.; Thai, R.; Mourier, G.; De Pauw, E. Identification of a novel snake peptide toxin displaying high affinity and antagonist behaviour for the α2--adrenoceptors. Br. J. Pharmacol. 2010, 161, 1361–1374. [Google Scholar] [CrossRef] [Green Version]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.-S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- De Weille, J.R.; Schweitz, H.; Maes, P.; Tartar, A.; Lazdunski, M. Calciseptine, a peptide isolated from black mamba venom, is a specific blocker of the L-type calcium channel. Proc. Natl. Acad. Sci. USA 1991, 88, 2437–2440. [Google Scholar] [CrossRef] [Green Version]
- Harvey, A.L.; Kornisiuk, E.; Bradley, K.N.; Cerveñansky, C.; Durán, R.; Adrover, M.; Sánchez, G.; Jerusalinsky, D. Effects of muscarinic toxins MT1 and MT2 from green mamba on different muscarinic cholinoceptors. Neurochem. Res. 2002, 27, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Kornisiuk, E.; Jerusalinsky, D.; Cerveñansky, C.; Harvey, A.L. Binding of muscarinic toxins MT×1 and MT×2 from the venom of the green mamba Dendroaspis angusticeps to cloned human muscarinic cholinoceptors. Toxicon 1995, 33, 11–18. [Google Scholar] [CrossRef]
- Mordvintsev, D.Y.; Polyak, Y.L.; Rodionov, D.I.; Jakubik, J.; Dolezal, V.; Karlsson, E.; Tsetlin, V.I.; Utkin, Y.N. Weak toxin WTX from Naja kaouthia cobra venom interacts with both nicotinic and muscarinic acetylcholine receptors. FEBS J. 2009, 276, 5065–5075. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Deuis, J.; Dashevsky, D.; Dobson, J.; Jackson, T.; Brust, A.; Xie, B.; Koludarov, I.; Debono, J.; Hendrikx, I. The snake with the scorpion’s sting: Novel three-finger toxin sodium channel activators from the venom of the long-glanded blue coral snake (Calliophis bivirgatus). Toxins 2016, 8, 303. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.; Mbugua, P.; Rodriguez-Ithurralde, D. Fasciculins, anticholinesterase toxins from the venom of the green mamba Dendroaspis angusticeps. J. Physiol. 1984, 79, 232–240. [Google Scholar]
- Banerjee, Y.; Lakshminarayanan, R.; Vivekanandan, S.; Anand, G.S.; Valiyaveettil, S.; Kini, R.M. Biophysical characterization of anticoagulant hemextin AB complex from the venom of snake Hemachatus haemachatus. Biophys. J. 2007, 93, 3963–3976. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, Y.; Mizuguchi, J.; Iwanaga, S.; Kini, R.M. Hemextin AB complex-a unique anticogulant protein complex from Hemachatus haemachatus (African Ringhals cobra) venom that inhibits clot initiation and factor VIIa activity. J. Biol. Chem. 2005, 280, 42601–42611. [Google Scholar] [CrossRef] [Green Version]
- McDowell, R.S.; Dennis, M.S.; Louie, A.; Shuster, M.; Mulkerrin, M.G.; Lazarus, R.A. Mambin, a potent glycoprotein IIb-IIIa antagonist and platelet aggregation inhibitor structurally related to the short neurotoxins. Biochemistry 1992, 31, 4766–4772. [Google Scholar] [CrossRef]
- Kini, R.M.; Evans, H.J. A common cytolytic region in myotoxins, hemolysins, cardiotoxins and antibacterial peptides. Int. J. Pept. Protein Res. 1989, 34, 277–286. [Google Scholar] [CrossRef]
- Kini, R.M.; Evans, H.J. Role of cationic residues in cytolytic activity: Modification of lysine residues in the cardiotoxin from Naja nigricollis venom and correlation between cytolytic and antiplatelet activity. Biochemistry 1989, 28, 9209–9215. [Google Scholar] [CrossRef]
- Da Silva, N.J., Jr.; Aird, S.D. Prey specificity, comparative lethality and compositional differences of coral snake venoms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 128, 425–456. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Sixberry, N.M.; Heyborne, W.H.; Fritts, T. Venom of the Brown Treesnake, Boiga irregularis: Ontogenetic shifts and taxa-specific toxicity. Toxicon 2006, 47, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.J.; Isbister, G.K.; O’Donnell, P.; Williamson, N.A.; Hodgson, W.C. Species differences in the neuromuscular activity of post-synaptic neurotoxins from two Australian black snakes (Pseudechis porphyriacus and Pseudechis colletti). Toxicol. Lett. 2013, 219, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.J.; Isbister, G.K.; Hodgson, W.C. In vitro neurotoxic effects of Pseudechis spp. venoms: A comparison of avian and murine skeletal muscle preparations. Toxicon 2013, 63, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Ghezellou, P.; Paiva, O.; Matainaho, T.; Ghassempour, A.; Goudarzi, H.; Kraus, F.; Sanz, L.; Williams, D.J. Snake venomics of two poorly known Hydrophiinae: Comparative proteomics of the venoms of terrestrial Toxicocalamus longissimus and marine Hydrophis cyanocinctus. J. Proteom. 2012, 75, 4091–4101. [Google Scholar] [CrossRef]
- Laustsen, A.H.; Gutiérrez, J.M.; Rasmussen, A.R.; Engmark, M.; Gravlund, P.; Sanders, K.L.; Lohse, B.; Lomonte, B. Danger in the reef: Proteome, toxicity, and neutralization of the venom of the olive sea snake, Aipysurus laevis. Toxicon 2015, 107, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Lomonte, B.; Pla, D.; Sasa, M.; Tsai, W.-C.; Solórzano, A.; Ureña-Díaz, J.M.; Fernández-Montes, M.L.; Mora-Obando, D.; Sanz, L.; Gutiérrez, J.M. Two color morphs of the pelagic yellow-bellied sea snake, Pelamis platura, from different locations of Costa Rica: Snake venomics, toxicity, and neutralization by antivenom. J. Proteom. 2014, 103, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Pahari, S.; Bickford, D.; Fry, B.G.; Kini, R.M. Expression pattern of three-finger toxin and phospholipase A2 genes in the venom glands of two sea snakes, Lapemis curtus and Acalyptophis peronii: Comparison of evolution of these toxins in land snakes, sea kraits and sea snakes. BMC Evol. Biol. 2007, 7, 175. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Wüster, W.; Ryan Ramjan, S.F.; Jackson, T.; Martelli, P.; Kini, R.M. Analysis of Colubroidea snake venoms by liquid chromatography with mass spectrometry: Evolutionary and toxinological implications. Rapid Commun. Mass Spectrom. 2003, 17, 2047–2062. [Google Scholar] [CrossRef]
- Sanz, L.; de Freitas-Lima, L.N.; Quesada-Bernat, S.; Graça-de-Souza, V.K.; Soares, A.M.; Calderón, L.d.A.; Calvete, J.J.; Caldeira, C.A. Comparative venomics of Brazilian coral snakes: Micrurus frontalis, Micrurus spixii spixii, and Micrurus surinamensis. Toxicon 2019, 166, 39–45. [Google Scholar] [CrossRef]
- Aird, S.D.; Da Silva, N.J.; Qiu, L.; Villar-Briones, A.; Saddi, V.A.; Pires de Campos Telles, M.; Grau, M.L.; Mikheyev, A.S. Coralsnake venomics: Analyses of venom gland transcriptomes and proteomes of six Brazilian taxa. Toxins 2017, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Debono, J.; Harrich, D.; Fry, B.G. Evolutionary Interpretations of Nicotinic Acetylcholine Receptor Targeting Venom Effects by a Clade of Asian Viperidae Snakes. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 236, 108769. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Harrich, D.; Frank, N.; Fry, B.G. An appetite for destruction: Detecting prey-selective binding of α-neurotoxins in the venom of Afro-Asian elapids. Toxins 2020, 12, 205. [Google Scholar] [CrossRef] [Green Version]
- Zdenek, C.N.; Harris, R.J.; Kuruppu, S.; Youngman, N.J.; Dobson, J.S.; Debono, J.; Khan, M.; Smith, I.; Yarski, M.; Harrich, D. A Taxon-Specific and High-Throughput Method for Measuring Ligand Binding to Nicotinic Acetylcholine Receptors. Toxins 2019, 11, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heatwole, H.; Poran, N.S. Resistances of sympatric and allopatric eels to sea snake venoms. Copeia 1995, 15, 136–147. [Google Scholar] [CrossRef]
- Heatwole, H.; Powell, J. Resistance of eels (Gymnothorax) to the venom of sea kraits (Laticauda colubrina): A test of coevolution. Toxicon 1998, 36, 619–625. [Google Scholar] [CrossRef]
- Drabeck, D.H.; Dean, A.M.; Jansa, S.A. Why the honey badger don’t care: Convergent evolution of venom-targeted nicotinic acetylcholine receptors in mammals that survive venomous snake bites. Toxicon 2015, 99, 68–72. [Google Scholar] [CrossRef]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Cobra (Naja spp.) nicotinic acetylcholine receptor exhibits resistance to erabu sea snake (Laticauda semifasciata) short-chain α-neurotoxin. J. Mol. Evol. 2004, 58, 516–526. [Google Scholar] [CrossRef]
- Barchan, D.; Kachalsky, S.; Neumann, D.; Vogel, Z.; Ovadia, M.; Kochva, E.; Fuchs, S. How the mongoose can fight the snake: The binding site of the mongoose acetylcholine receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 7717–7721. [Google Scholar] [CrossRef] [Green Version]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [Green Version]
- Voris, H. The role of sea snakes (Hydrophiidae) in the trophic structure of coastal ocean communities. J. Mar. Biol. Assoc. India 1972, 14, 429–442. [Google Scholar]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet breadth mediates the prey specificity of venom potency in snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokyta, D.R.; Wray, K.P.; Margres, M.J. The genesis of an exceptionally lethal venom in the timber rattlesnake (Crotalus horridus) revealed through comparative venom-gland transcriptomics. BMC Genom. 2013, 14, 394. [Google Scholar] [CrossRef] [Green Version]
- Rokyta, D.R.; Wray, K.P.; McGivern, J.J.; Margres, M.J. The transcriptomic and proteomic basis for the evolution of a novel venom phenotype within the Timber Rattlesnake (Crotalus horridus). Toxicon 2015, 98, 34–48. [Google Scholar] [CrossRef]
- Pardal, P.P.d.O.; Pardal, J.S.d.O.; Gadelha, M.A.d.C.; Rodrigues, L.d.S.; Feitosa, D.T.; Prudente, A.L.d.C.; Fan, H.W. Envenomation by Micrurus coral snakes in the Brazilian Amazon region: Report of two cases. Rev. Inst. Med. Trop. São Paulo 2010, 52, 333–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, C.M. Sea snake envenomation. Dermatol. Ther. 2002, 15, 58–61. [Google Scholar] [CrossRef]
- Panagides, N.; Jackson, T.; Ikonomopoulou, M.; Arbuckle, K.; Pretzler, R.; Yang, D.; Ali, S.; Koludarov, I.; Dobson, J.; Sanker, B. How the cobra got its flesh-eating venom: Cytotoxicity as a defensive innovation and its co-evolution with hooding, aposematic marking, and spitting. Toxins 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Chiappinelli, V.A.; Weaver, W.R.; McLane, K.E.; Conti-Fine, B.M.; Fiordalisi, J.J.; Grant, G.A. Binding of native κ-neurotoxins and site-directed mutants to nicotinic acetylcholine receptors. Toxicon 1996, 34, 1243–1256. [Google Scholar] [CrossRef]
- Harris, R.J.; Jenner, R.A. Evolutionary ecology of fish venom: Adaptations and consequences of evolving a venom system. Toxins 2019, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.J. Diversity, phylogenetic distribution, and origins of venomous catfishes. BMC Evol. Biol. 2009, 9, 282. [Google Scholar] [CrossRef] [Green Version]
- McLane, K.E.; Wu, X.; Conti-Tronconi, B.M. An α-bungarotoxin-binding sequence on the Torpedo nicotinic acetylcholine receptor α-subunit: Conservative amino acid substitutions reveal side-chain specific interactions. Biochemistry 1994, 33, 2576–2585. [Google Scholar] [CrossRef] [PubMed]
- McLane, K.E.; Wu, X.; Diethelm, B.; Conti-Tronconi, B.M. Structural determinants of α-bungarotoxin binding to the sequence segment 181-200 of the muscle nicotinic acetylcholine receptor. α-subunit: Effects of cysteine/cystine modification and species-specific amino acid substitutions. Biochemistry 1991, 30, 4925–4934. [Google Scholar] [CrossRef] [PubMed]
- Testai, F.D.; Venera, G.D.; Peña, C.; de Jiménez Bonino, M.J.B. Histidine 186 of the nicotinic acetylcholine receptor α subunit requires the presence of the 192–193 disulfide bridge to interact with α-bungarotoxin. Neurochem. Int. 2000, 36, 27–33. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, R.J.; Youngman, N.J.; Zdenek, C.N.; Huynh, T.M.; Nouwens, A.; Hodgson, W.C.; Harrich, D.; Dunstan, N.; Portes-Junior, J.A.; Fry, B.G. Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models. Int. J. Mol. Sci. 2020, 21, 7377. https://doi.org/10.3390/ijms21197377
Harris RJ, Youngman NJ, Zdenek CN, Huynh TM, Nouwens A, Hodgson WC, Harrich D, Dunstan N, Portes-Junior JA, Fry BG. Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models. International Journal of Molecular Sciences. 2020; 21(19):7377. https://doi.org/10.3390/ijms21197377
Chicago/Turabian StyleHarris, Richard J., Nicholas J. Youngman, Christina N. Zdenek, Tam M. Huynh, Amanda Nouwens, Wayne C. Hodgson, David Harrich, Nathan Dunstan, José A. Portes-Junior, and Bryan G. Fry. 2020. "Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models" International Journal of Molecular Sciences 21, no. 19: 7377. https://doi.org/10.3390/ijms21197377
APA StyleHarris, R. J., Youngman, N. J., Zdenek, C. N., Huynh, T. M., Nouwens, A., Hodgson, W. C., Harrich, D., Dunstan, N., Portes-Junior, J. A., & Fry, B. G. (2020). Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models. International Journal of Molecular Sciences, 21(19), 7377. https://doi.org/10.3390/ijms21197377