Allopregnanolone Enhances GABAergic Inhibition in Spinal Motor Networks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
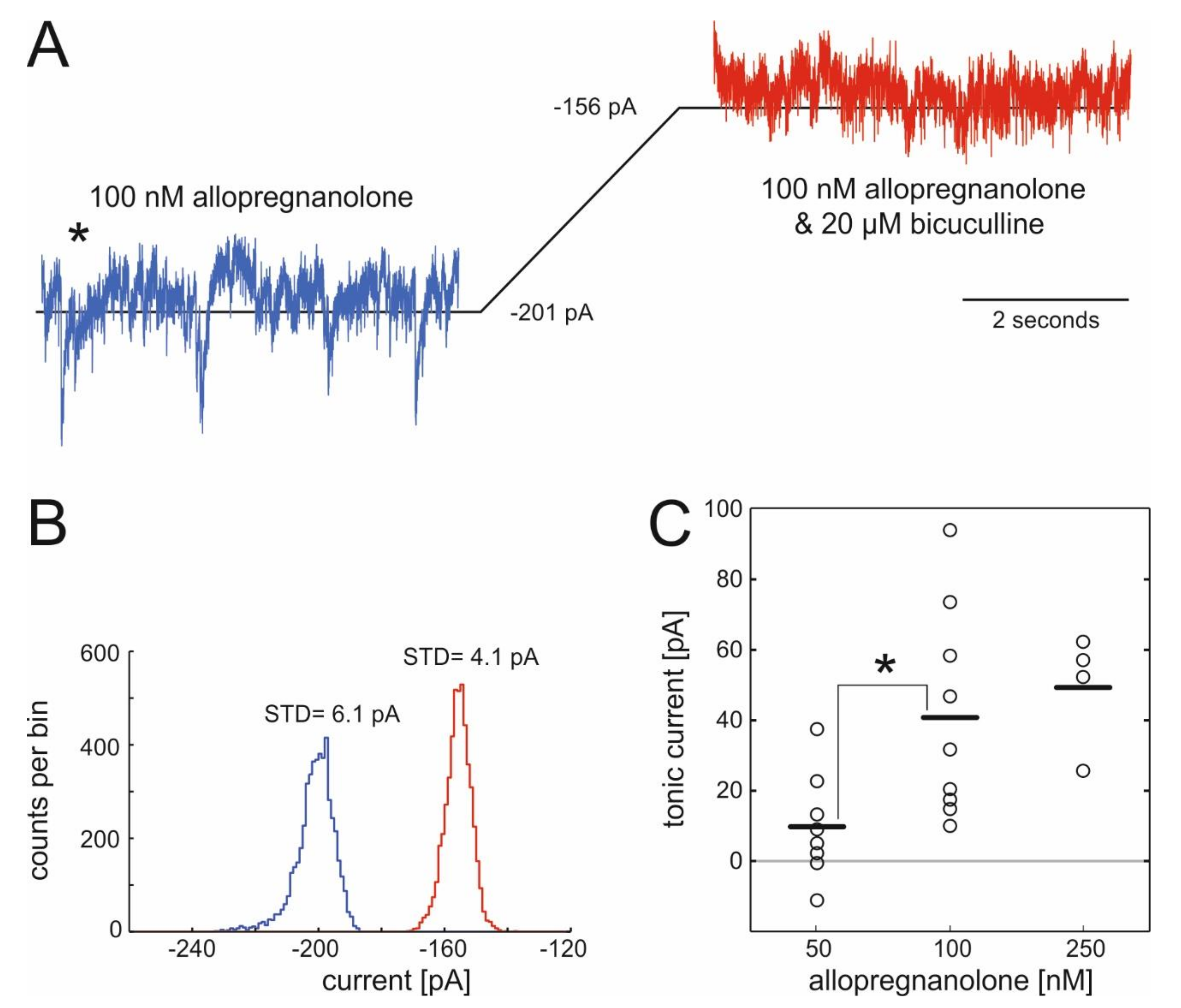
2.1. Actions of Allopregnanolone on Tonic GABAergic Inhibition
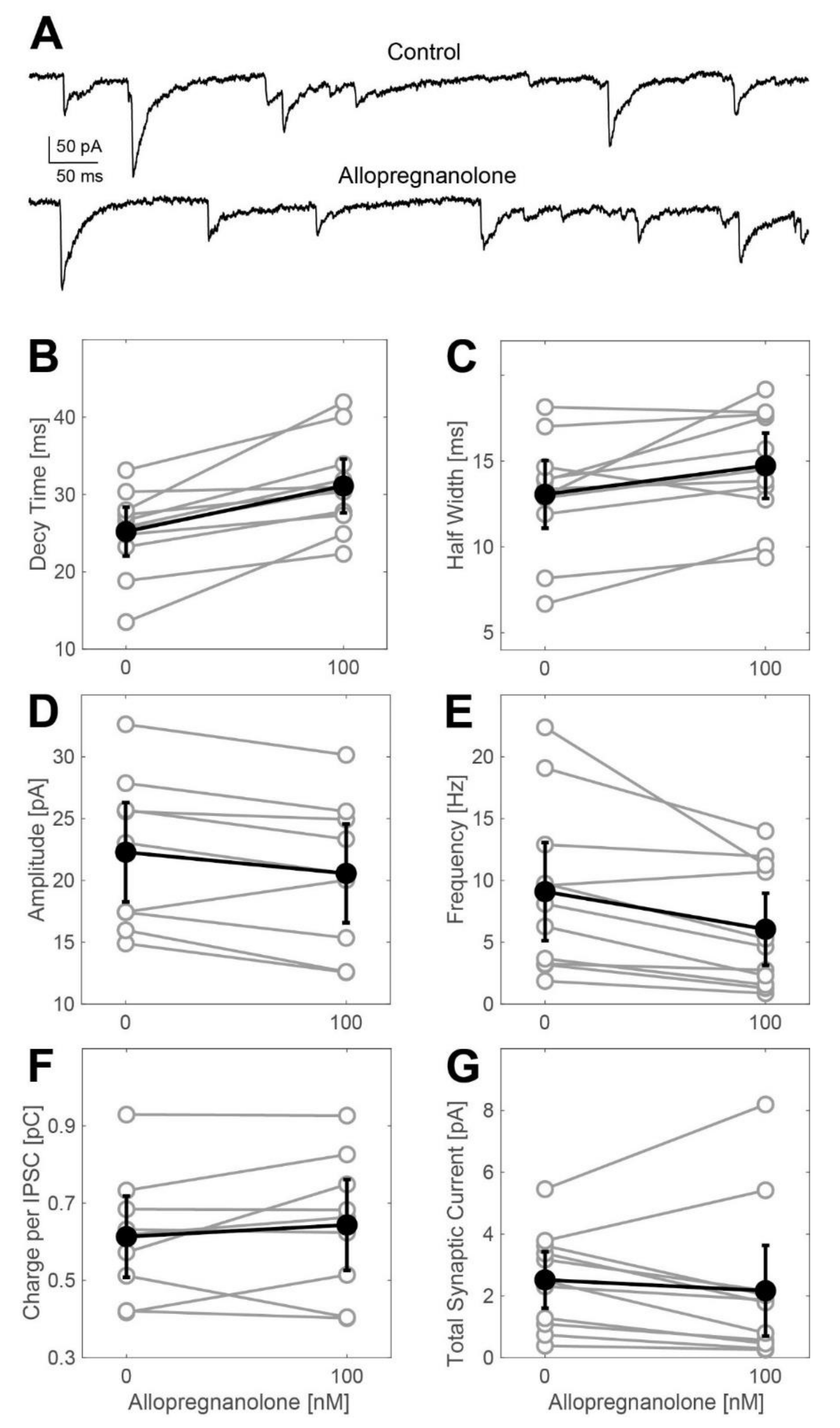
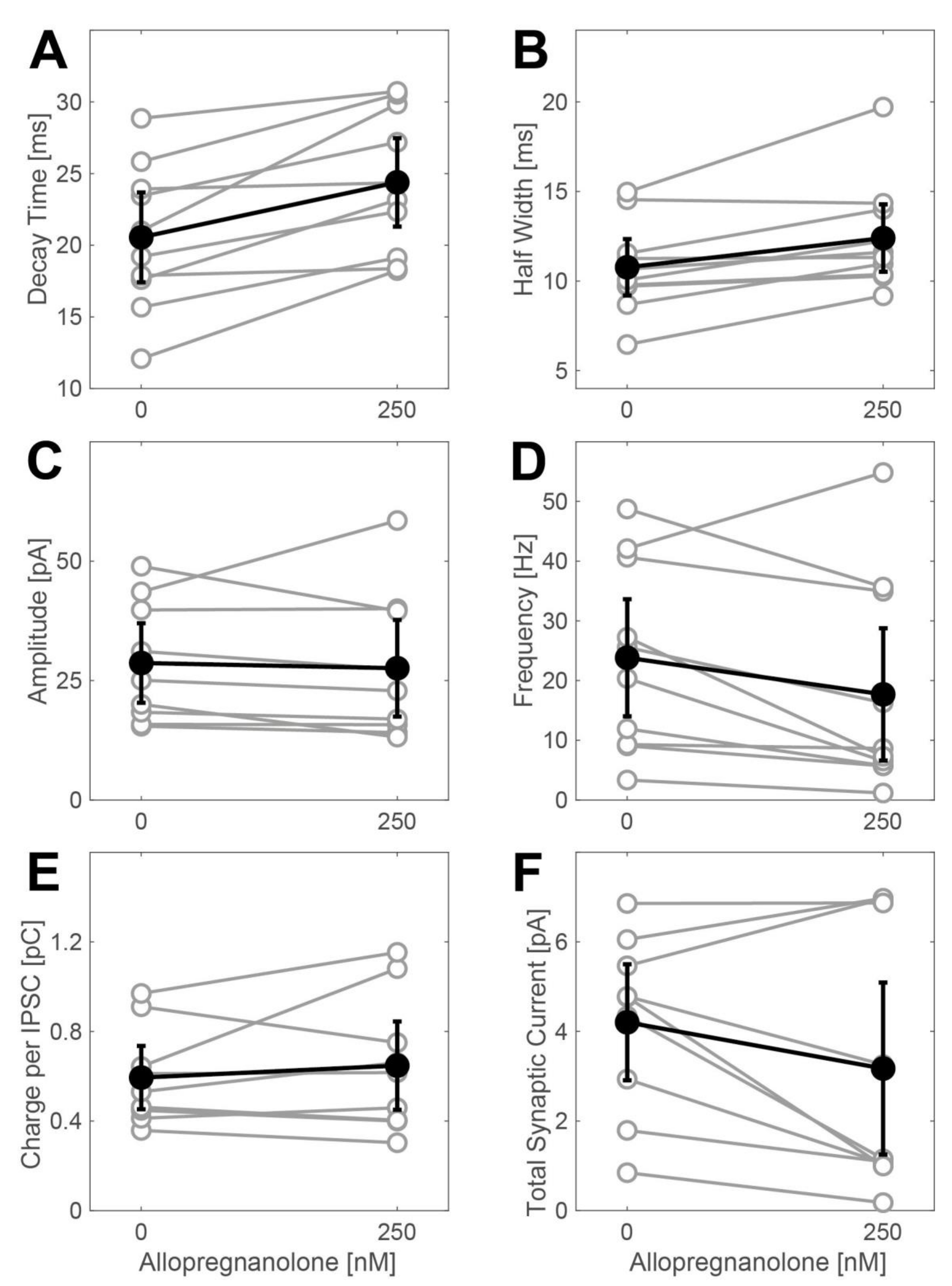
2.2. Actions of Allopregnanolone on Phasic GABAergic Inhibition
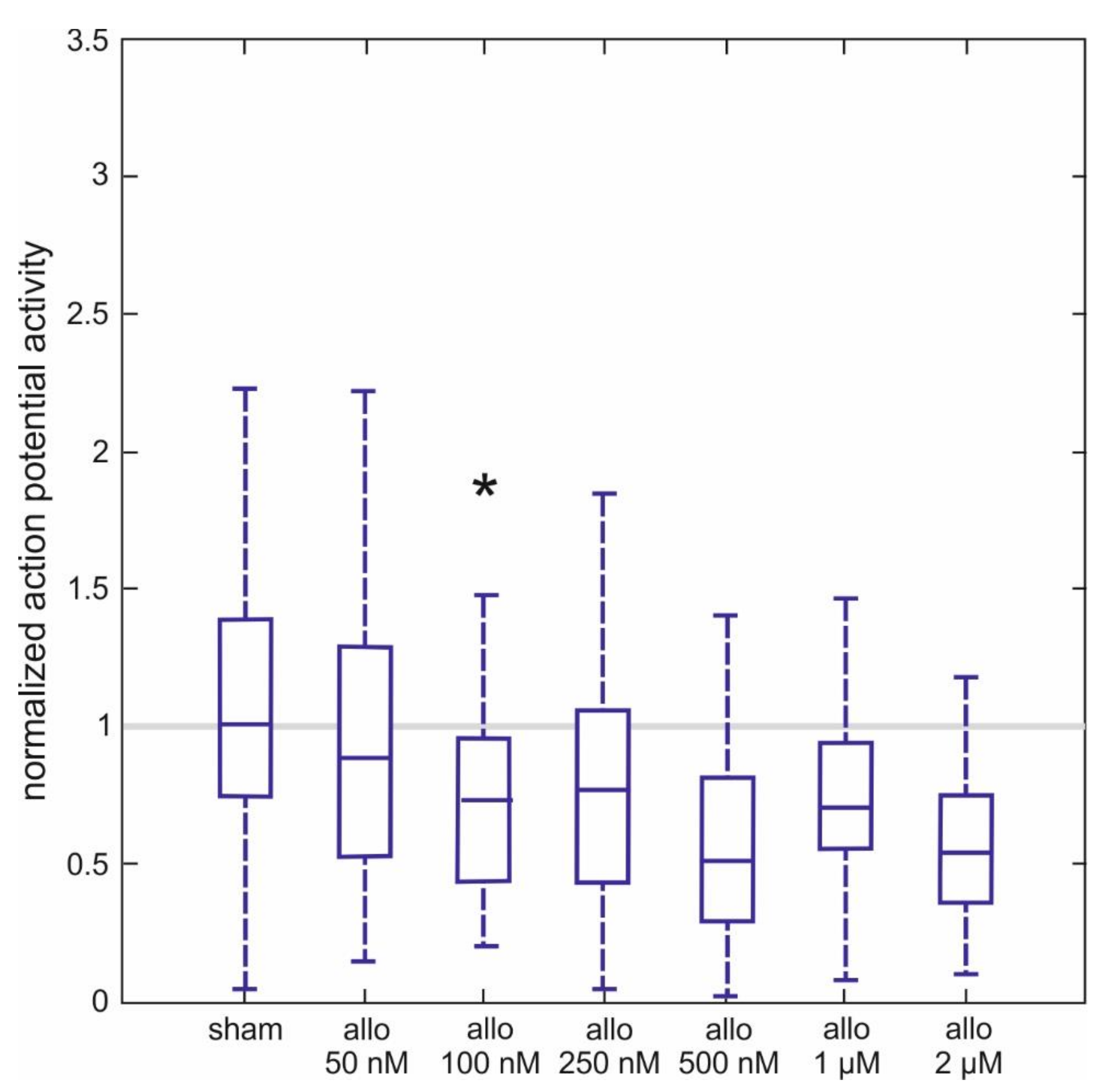
2.3. Effect of Allopregnanolone on Action Potential Firing of Spinal Neurons
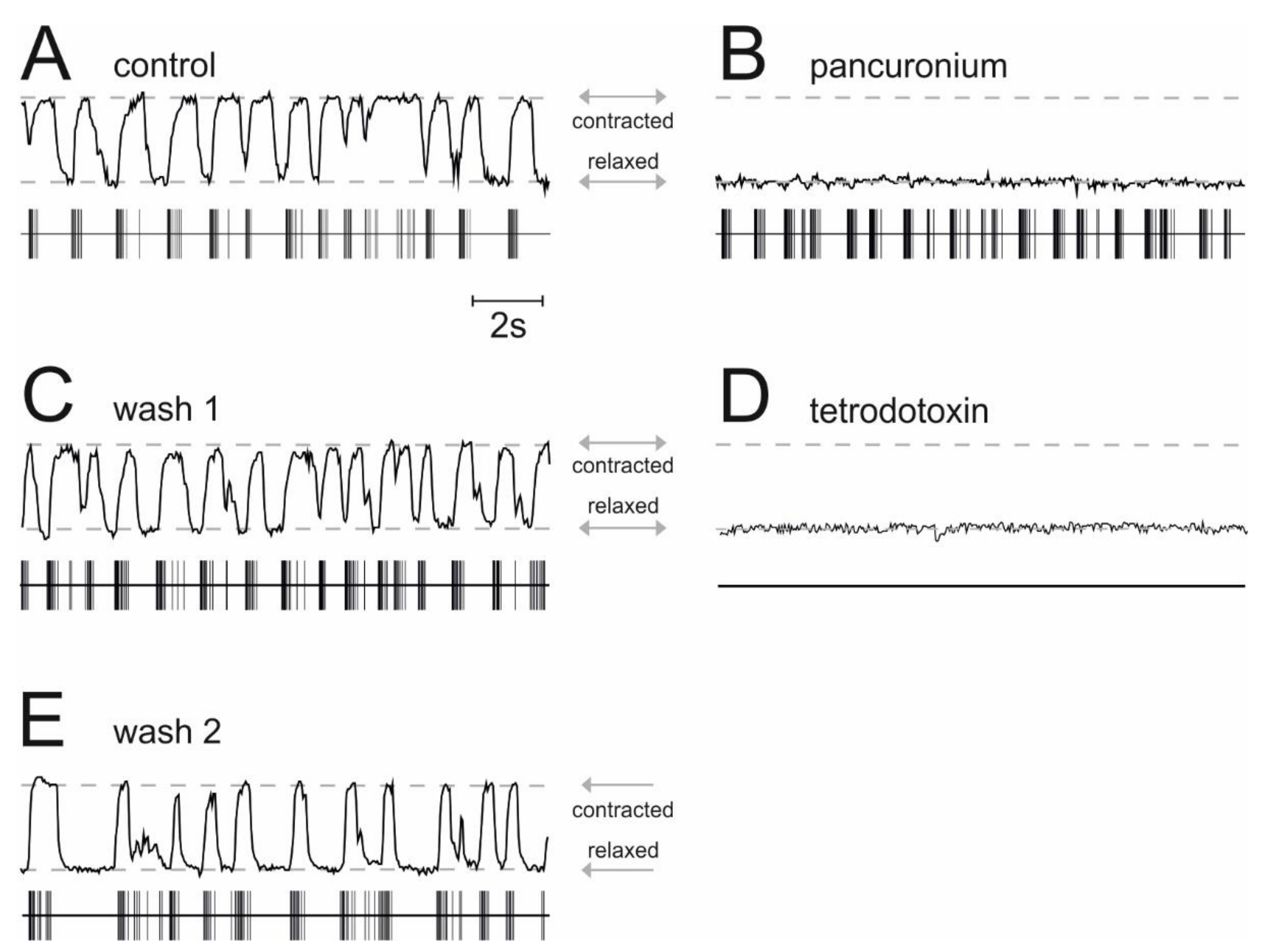
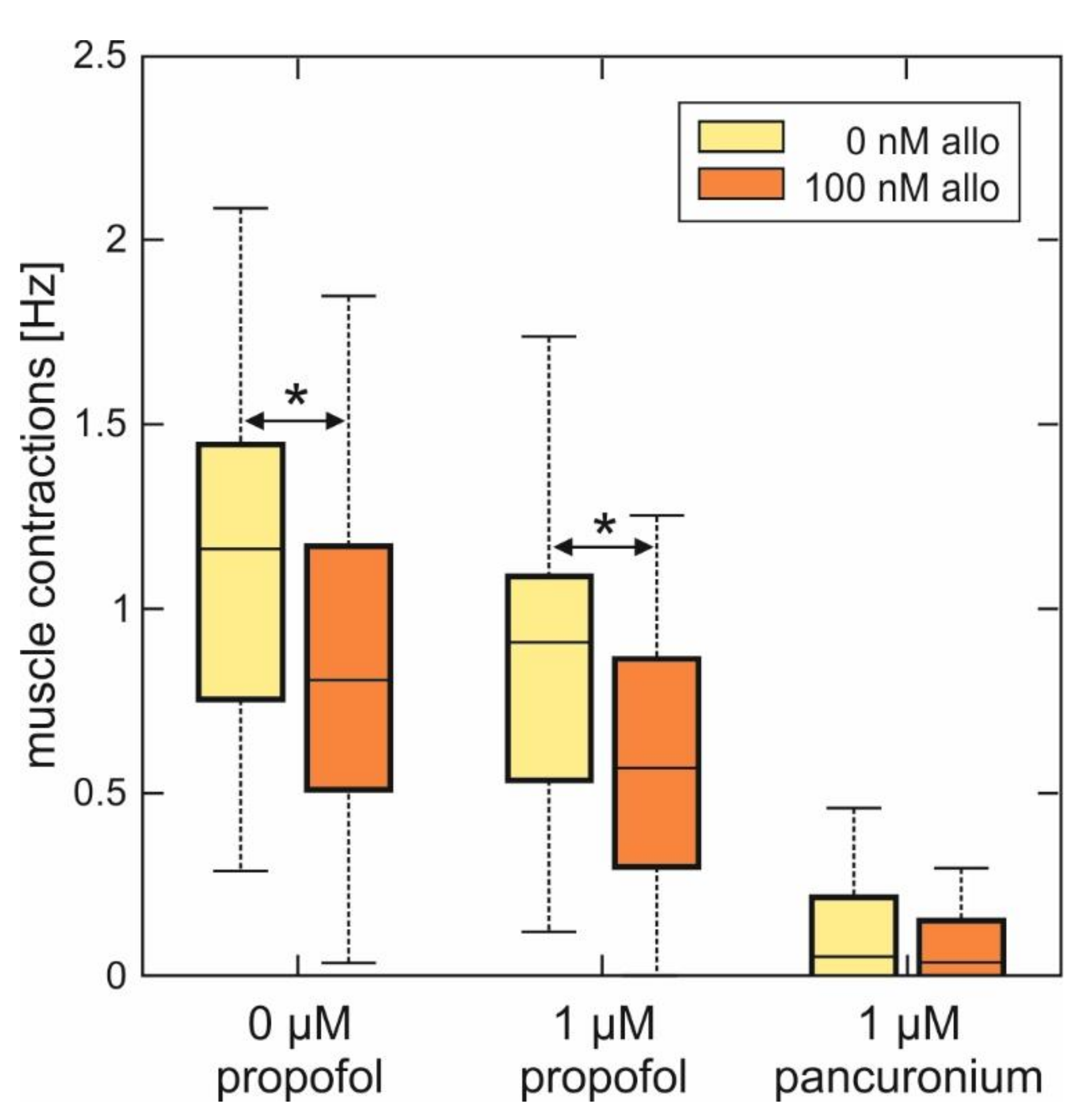
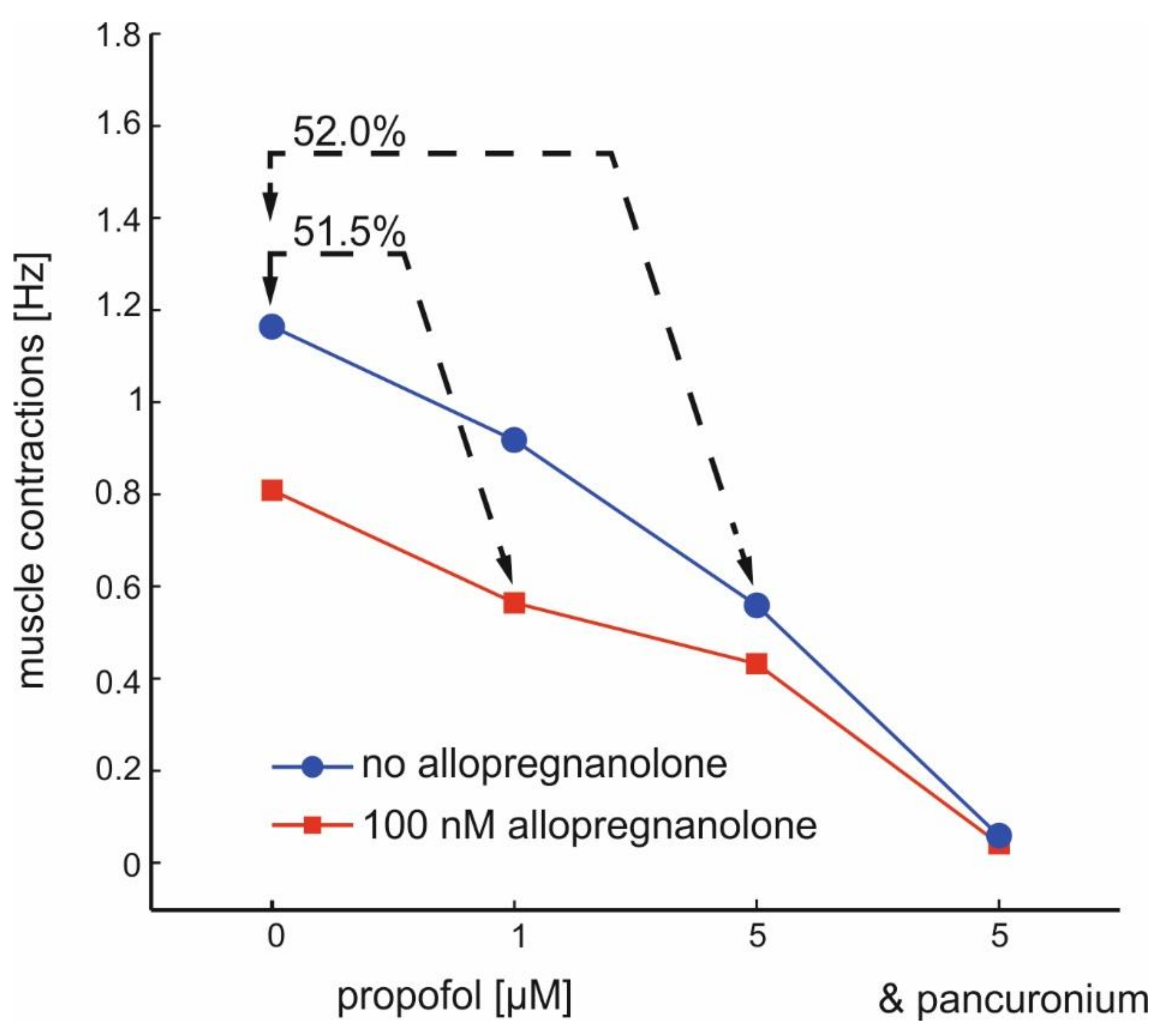
2.4. Actions of Allopregnanolone on Muscle Activity in Spinal Nerve and Muscle Co-Cultures
3. Discussion
3.1. Allopregnanolone Induces a Tonic GABAA-R-Mediated Current
3.2. Actions of Allopregnanolone on Muscle Contractions
3.3. Effects of Allopregnanolone on Synaptic GABAA Receptors Are of Minor Importance
3.4. Comparison of Spinal and Cortical Actions of Allopregnanolone
3.5. Limitations of the Present Study
3.6. Implications for Further Studies
4. Materials and Methods
4.1. Preparation of Nerve and Muscle Co-Cultures
4.2. Preparation and Application of Test Solutions
4.3. Recording and Analysis of Electrophysiological Data
4.4. Recording and Analysis of Muscle Activity
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALLO | allopregnanolone |
| GABA | γ-aminobutyric acid |
Appendix A

References
- Goodchild, C.S.; Serrao, J.M.; Kolosov, A.; Boyd, B.J. Alphaxalone Reformulated. Anesthesia Analg. 2015, 120, 1025–1031. [Google Scholar] [CrossRef]
- Monagle, J.; Siu, L.; Worrell, J.; Goodchild, C.S.; Serrao, J.M. A Phase 1c Trial Comparing the Efficacy and Safety of a New Aqueous Formulation of Alphaxalone with Propofol. Anesthesia Analg. 2015, 121, 914–924. [Google Scholar] [CrossRef] [Green Version]
- Antkowiak, B.; Rammes, G. GABA(A) receptor-targeted drug development -New perspectives in perioperative anesthesia. Expert Opin. Drug Discov. 2019, 14, 683–699. [Google Scholar] [CrossRef]
- Schumacher, M.; Mattern, C.; Ghoumari, A.; Oudinet, J.; Liere, P.; Labombarda, F.; Sitruk-Ware, R.; De Nicola, A.; Guennoun, R. Revisiting the roles of progesterone and allopregnanolone in the nervous system: Resurgence of the progesterone receptors. Prog. Neurobiol. 2014, 113, 6–39. [Google Scholar] [CrossRef]
- Lancel, M.; Faulhaber, J.; Schiffelholz, T.; Romeo, E.; Di Michele, F.; Holsboer, F.; Rupprecht, R. Allopregnanolone affects sleep in a benzodiazepine-like fashion. J. Pharmacol. Exp. Ther. 1997, 282, 1213–1218. [Google Scholar] [PubMed]
- Reddy, D.S.; Zeng, Y.-C. Differential anesthetic activity of ketamine and the GABAergic neurosteroid allopregnanolone in mice lacking progesterone receptor A and B subtypes. Methods Find. Exp. Clin. Pharmacol. 2007, 29, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Hosie, A.M.; Wilkins, M.E.; Da Silva, H.M.A.; Smart, T.G. Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites. Nature 2006, 444, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Forman, S.A.; Miller, K.W. Mapping General Anesthetic Sites in Heteromeric γ-Aminobutyric Acid Type A Receptors Reveals a Potential For Targeting Receptor Subtypes. Anesthesia Analg. 2016, 123, 1263–1273. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W.; Sieghart, W. GABAA receptors: Subtypes provide diversity of function and pharmacology. Neuropharmacology 2009, 56, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Farrant, M.; Nusser, Z. Variations on an inhibitory theme: Phasic and tonic activation of GABAA receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef]
- Brickley, S.G.; Mody, I. Extrasynaptic GABAA Receptors: Their Function in the CNS and Implications for Disease. Neuron 2012, 73, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonin, R.P.; Labrakakis, C.; Eng, D.G.; Whissell, P.D.; De Koninck, Y.; Orser, B.A. Pharmacological enhancement of δ-subunit-containing GABAA receptors that generate a tonic inhibitory conductance in spinal neurons attenuates acute nociception in mice. Pain 2011, 152, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Rampil, I.J.; Mason, P.; Singh, H. Anesthetic Potency (MAC) Is Independent of Forebrain Structures in the Rat. Anesthesiology 1993, 78, 707–712. [Google Scholar] [CrossRef]
- Antognini, J.F.; Wang, X.-W.; Piercy, M.; Carstens, E. Propofol directly depresses lumbar dorsal horn neuronal responses to noxious stimulation in goats. Can. J. Anesth. 2000, 47, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, B.T.; Kesselheim, A.S. Propofol as a transformative drug in anesthesia: Insights from key early investigators. Drug Discov. Today 2015, 20, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Drexler, B.; Balk, M.; Antkowiak, B. Synergistic Modulation of γ-Aminobutyric Acid Type A Receptor-Mediated Synaptic Inhibition in Cortical Networks by Allopregnanolone and Propofol. Anesthesia Analg. 2016, 123, 877–883. [Google Scholar] [CrossRef]
- Spenger, C.; Braschler, U.F.; Streit, J.; Lüscher, H.-R. An Organotypic Spinal Cord-Dorsal Root Ganglion-Skeletal Muscle Coculture of Embryonic Rat. I. The Morphological Correlates of the Spinal Reflex Arc. Eur. J. Neurosci. 1991, 3, 1037–1053. [Google Scholar] [CrossRef]
- Streit, J.; Spenger, C.; Lüscher, H.-R. An Organotypic Spinal Cord-Dorsal Root Ganglion-Skeletal Muscle Coculture of Embryonic Rat. II. Functional Evidence for the Formation of Spinal Reflex Arcs In Vitro. Eur. J. Neurosci. 1991, 3, 1054–1068. [Google Scholar] [CrossRef]
- Avossa, D.; Rosato-Siri, M.D.; Mazzarol, F.; Ballerini, L. Spinal circuits formation: A study of developmentally regulated markers in organotypic cultures of embryonic mouse spinal cord. Neuroscience 2003, 122, 391–405. [Google Scholar] [CrossRef]
- Drexler, B.; Hentschke, H.; Antkowiak, B.; Grasshoff, C. Organotypic cultures as tools for testing neuroactive drugs - link between in-vitro and in-vivo experiments. Curr. Med. Chem. 2010, 17, 4538–4550. [Google Scholar] [CrossRef]
- Drexler, B.; Thiermann, H.; Antkowiak, B.; Grasshoff, C. Effects of succinylcholine in an organotypic spinal cord-skeletal muscle coculture of embryonic mice. Chem. Interactions 2013, 206, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Weimer, I.; Worek, F.; Seeger, T.; Thiermann, H.; Eckle, V.; Grasshoff, C.; Antkowiak, B. Self-regeneration of neuromuscular function following soman and VX poisoning in spinal cord—Skeletal muscle cocultures. Toxicol. Lett. 2016, 244, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Eckle, V.-S.; Drexler, B.; Grasshoff, C.; Seeger, T.; Thiermann, H.; Antkowiak, B. Spinal cord-skeletal muscle cocultures detect muscle-relaxant action of botulinum neurotoxin A. ALTEX 2014, 31. [Google Scholar] [CrossRef] [Green Version]
- Drexler, B.; Seeger, T.; Grasshoff, C.; Thiermann, H.; Antkowiak, B. Long-term evaluation of organophosphate toxicity and antidotal therapy in co-cultures of spinal cord and muscle tissue. Toxicol. Lett. 2011, 206, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Jordan, L.M.; McVagh, J.R.; Noga, B.R.; Cabaj, A.M.; Majczyå„Ski, H.; Så‚Awiå„Ska, U.; Provencher, J.; Leblond, H.; Rossignol, S.; Majczyński, H.; et al. Cholinergic mechanisms in spinal locomotion—potential target for rehabilitation approaches. Front. Neural Circuits 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Eckle, V.S.; Rudolph, U.; Antkowiak, B.; Grasshoff, C. Propofol modulates phasic and tonic GABAergic currents in spinal ventral horn interneurones. Br. J. Anaesth. 2015, 114, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Carver, C.M.; Reddy, D.S.; Matthew, C.C.; Samba, R.D. Neurosteroid interactions with synaptic and extrasynaptic GABA(A) receptors: Regulation of subunit plasticity, phasic and tonic inhibition, and neuronal network excitability. Psychopharmacology 2013, 230, 151–188. [Google Scholar] [CrossRef]
- Minier, F.; Sigel, E. Positioning of the -subunit isoforms confers a functional signature to -aminobutyric acid type A receptors. Proc. Natl. Acad. Sci. USA 2004, 101, 7769–7774.29. [Google Scholar] [CrossRef] [Green Version]
- Andrés, C.; Aguilar, J.; González-Ramírez, R.; Elías-Viñas, D.; Felix, R.; Delgado-Lezama, R. Extrasynaptic α6 Subunit-Containing GABAA Receptors Modulate Excitability in Turtle Spinal Motoneurons. PLoS ONE 2014, 9, e115378. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.; Aguilar, J.; González-Ramírez, R.; Loeza-Alcocer, E.; Canto-Bustos, M.; Felix, R.; Delgado-Lezama, R. Tonic inhibition in spinal ventral horn interneurons mediated by α5 subunit-containing GABAA receptors. Biochem. Biophys. Res. Commun. 2011, 412, 26–31. [Google Scholar] [CrossRef]
- Delgado-Lezama, R.; Loeza-Alcocer, E.; Andrés, C.; Aguilar, J.; Guertin, A.P.; Felix, R. Extrasynaptic GABA(A) receptors in the brainstem and spinal cord: Structure and function. Curr. Pharm. Des. 2013, 19, 4485–4497. [Google Scholar] [CrossRef] [PubMed]
- Canto-Bustos, M.; Loeza-Alcocer, E.; Cuellar, C.A.; Osuna, P.; Elias-Viñas, D.; Granados-Soto, V.; Manjarrez, E.; Felix, R.; Delgado-Lezama, R. Tonically Active α5GABAA Receptors Reduce Motoneuron Excitability and Decrease the Monosynaptic Reflex. Front. Cell. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Reyes, J.E.; Salinas-Abarca, A.B.; Vidal-Cantú, G.C.; Raya-Tafolla, G.; Elias-Viñas, D.; Granados-Soto, V.; Delgado-Lezama, R. α5GABAA receptors play a pronociceptive role and avoid the rate-dependent depression of the Hoffmann reflex in diabetic neuropathic pain and reduce primary afferent excitability. Pain 2019, 160, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.K.; Delgado-Lezama, R.; Russo, R.E.; Lind, B.L.; Loeza-Alcocer, E.; Rath, M.F.; Fabbiani, G.; Schmitt, N.; Steinkellner, H.; Petersen, A.V.; et al. Spinal dorsal horn astrocytes release GABA in response to synaptic activation. J. Physiol. 2018, 596, 4983–4994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grasshoff, C.; Netzhammer, N.; Schweizer, J.; Antkowiak, B.; Hentschke, H. Depression of spinal network activity by thiopental: Shift from phasic to tonic GABAA receptor-mediated inhibition. Neuropharmacology 2008, 55, 793–802. [Google Scholar] [CrossRef]
- Seeger, T.; Worek, F.; Szinicz, L.; Thiermann, H. Reevaluation of indirect field stimulation technique to demonstrate oxime effectiveness in OP-poisoning in muscles in vitro. Toxicology 2007, 233, 209–213. [Google Scholar] [CrossRef]
- Rudolph, U.; Antkowiak, B. Molecular and neuronal substrates for general anaesthetics. Nat. Rev. Neurosci. 2004, 5, 709–720. [Google Scholar] [CrossRef]
- Di Cristo, G.; Wu, C.; Chattopadhyaya, B.; Ango, F.; Knott, G.; Welker, E.; Svoboda, K.; Huang, Z.J. Subcellular domain-restricted GABAergic innervation in primary visual cortex in the absence of sensory and thalamic inputs. Nat. Neurosci. 2004, 7, 1184–1186. [Google Scholar] [CrossRef]
- Li, G.-D.; Chiara, D.C.; Cohen, J.B.; Olsen, R.W. Neurosteroids Allosterically Modulate Binding of the Anesthetic Etomidate to γ-Aminobutyric Acid Type A Receptors. J. Biol. Chem. 2009, 284, 11771–11775. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, E.A.; Gentet, L.J.; Dempster, J.; Belelli, D. GABAA and glycine receptor-mediated transmission in rat lamina II neurones: Relevance to the analgesic actions of neuroactive steroids. J. Physiol. 2007, 583, 1021–1040. [Google Scholar] [CrossRef]
- Belelli, D.; Lambert, J.J. Neurosteroids: Endogenous regulators of the GABAA receptor. Nat. Rev. Neurosci. 2005, 6, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Harney, S.C.; Frenguelli, B.G.; Lambert, J.J. Phosphorylation influences neurosteroid modulation of synaptic GABAA receptors in rat CA1 and dentate gyrus neurones. Neuropharmacology 2003, 45, 873–883. [Google Scholar] [CrossRef]
- Braschler, U.; Iannone, A.; Spenger, C.; Streit, J.; Luscher, H.-R. A modified roller tube technique for organotypic cocultures of embryonic rat spinal cord, sensory ganglia and skeletal muscle. J. Neurosci. Methods 1989, 29, 121–129. [Google Scholar] [CrossRef]
- Gähwiler, B. Organotypic monolayer cultures of nervous tissue. J. Neurosci. Methods 1981, 4, 329–342. [Google Scholar] [CrossRef]
- Rosato-Siri, M.D.; Zoccolan, D.; Furlan, F.; Ballerini, L. Interneurone bursts are spontaneously associated with muscle contractions only during early phases of mouse spinal network development: A study in organotypic cultures. Eur. J. Neurosci. 2004, 20. [Google Scholar] [CrossRef]
- Gredell, J.A.; Turnquist, P.A.; Maciver, M.B.; Pearce, R.A. Determination of diffusion and partition coefficients of propofol in rat brain tissue: Implications for studies of drug action in vitro. Br. J. Anaesth. 2004, 93, 810–817. [Google Scholar] [CrossRef] [Green Version]
- Benkwitz, C.; Liao, M.; Laster, M.J.; Sonner, J.M.; Eger, E.I.; Pearce, R.A. Determination of the EC50 Amnesic Concentration of Etomidate and Its Diffusion Profile in Brain Tissue. Anesthesiology 2007, 106, 114–123. [Google Scholar] [CrossRef]
- Glykys, J.; Mody, I. The main source of ambient GABA responsible for tonic inhibition in the mouse hippocampus. J. Physiol. 2007, 582, 1163–1178. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drexler, B.; Grenz, J.; Grasshoff, C.; Antkowiak, B. Allopregnanolone Enhances GABAergic Inhibition in Spinal Motor Networks. Int. J. Mol. Sci. 2020, 21, 7399. https://doi.org/10.3390/ijms21197399
Drexler B, Grenz J, Grasshoff C, Antkowiak B. Allopregnanolone Enhances GABAergic Inhibition in Spinal Motor Networks. International Journal of Molecular Sciences. 2020; 21(19):7399. https://doi.org/10.3390/ijms21197399
Chicago/Turabian StyleDrexler, Berthold, Julia Grenz, Christian Grasshoff, and Bernd Antkowiak. 2020. "Allopregnanolone Enhances GABAergic Inhibition in Spinal Motor Networks" International Journal of Molecular Sciences 21, no. 19: 7399. https://doi.org/10.3390/ijms21197399
APA StyleDrexler, B., Grenz, J., Grasshoff, C., & Antkowiak, B. (2020). Allopregnanolone Enhances GABAergic Inhibition in Spinal Motor Networks. International Journal of Molecular Sciences, 21(19), 7399. https://doi.org/10.3390/ijms21197399