Rationale, Relevance, and Limits of Stress-Induced Psychopathology in Rodents as Models for Psychiatry Research: An Introductory Overview



{kind=link}
{kind=link}
Abstract
:1. Introduction
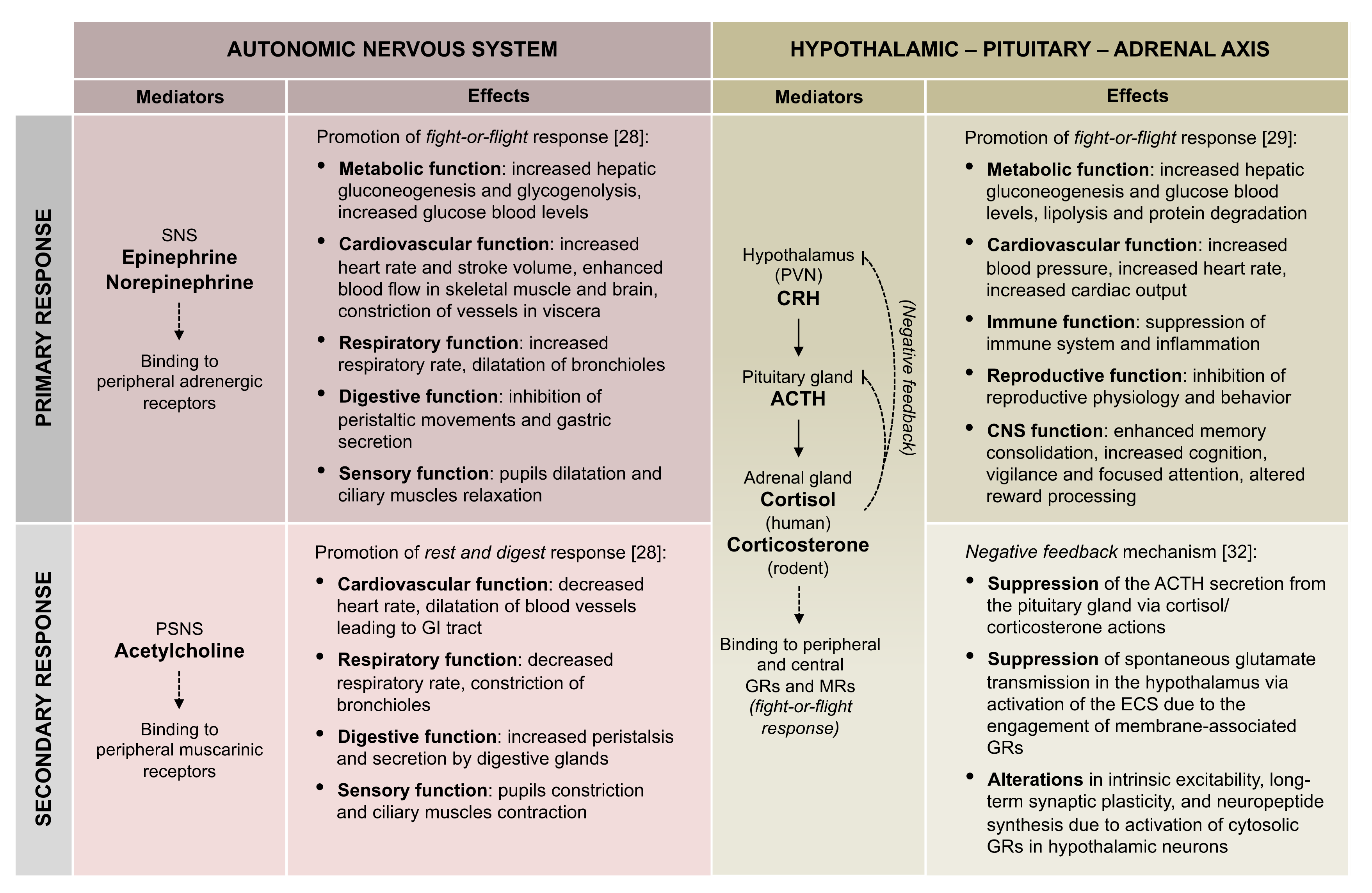
2. From Stress to Neuropsychiatric Disorders
3. Validity Assessment of Rodent Models of Environmental Stress
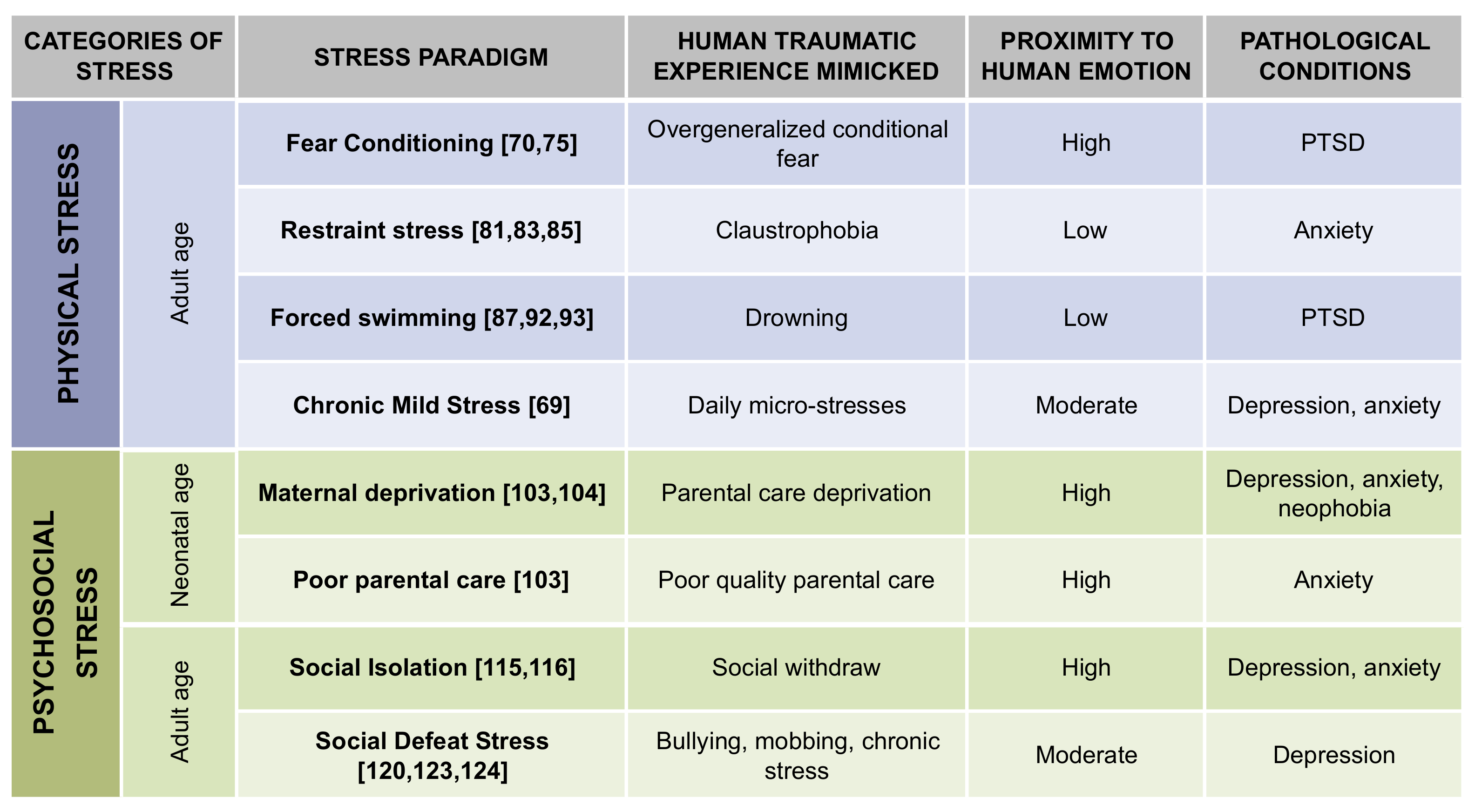
4. Animal Models of Stress, Feasibility, and Reliability as a Proxy for Mental Disorders
5. Physical Stress
5.1. Fear Conditioning
5.2. Restraint Stress
5.3. Forced Swimming
6. Chronic Mild Stress
7. Psychological Stress
7.1. Maternal Deprivation
7.2. Poor Parental Care
7.3. Social Isolation
7.4. Social Defeat Stress
8. Ethical Considerations
9. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- APA. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Burhan, A.M.; Marlatt, N.M.; Palaniyappan, L.; Anazodo, U.C.; Prato, F.S. Role of Hybrid Brain Imaging in Neuropsychiatric Disorders. Diagnostics 2015, 5, 577–614. [Google Scholar] [CrossRef] [Green Version]
- Pankow, A.; Katthagen, T.; Diner, S.; Deserno, L.; Boehme, R.; Kathmann, N.; Gleich, T.; Gaebler, M.; Walter, H.; Heinz, A.; et al. Aberrant Salience Is Related to Dysfunctional Self-Referential Processing in Psychosis. Schizophr. Bull. 2016, 42, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Grzenda, A.; Widge, A.S. Electroencephalographic Biomarkers for Predicting Antidepressant Response: New Methods, Old Questions. JAMA Psychiatry 2020, 77, 347. [Google Scholar] [CrossRef]
- Brambilla, P.; Nicoletti, M.A.; Harenski, K.; Sassi, R.B.; Mallinger, A.G.; Frank, E.; Kupfer, D.J.; Keshavan, M.S.; Soares, J.C. Anatomical MRI study of subgenual prefrontal cortex in bipolar and unipolar subjects. Neuropsychopharmacology 2002, 27, 792–799. [Google Scholar] [CrossRef] [Green Version]
- Brambilla, P.; Stanley, J.A.; Nicoletti, M.A.; Sassi, R.B.; Mallinger, A.G.; Frank, E.; Kupfer, D.; Keshavan, M.S.; Soares, J.C. 1H magnetic resonance spectroscopy investigation of the dorsolateral prefrontal cortex in bipolar disorder patients. J. Affect. Disord. 2005, 86, 61–67. [Google Scholar] [CrossRef]
- Maggioni, E.; Tana, M.G.; Arrigoni, F.; Zucca, C.; Bianchi, A.M. Constructing fMRI connectivity networks: A whole brain functional parcellation method for node definition. J. Neurosci. Methods 2014, 228, 86–99. [Google Scholar] [CrossRef]
- Maggioni, E.; DelVecchio, G.; Grottaroli, M.; Garzitto, M.; Piccin, S.; Bonivento, C.; Maieron, M.; D’Agostini, S.; Perna, G.; Balestrieri, M.; et al. Common and different neural markers in major depression and anxiety disorders: A pilot structural magnetic resonance imaging study. Psychiatry Res. Neuroimaging 2019, 290, 42–50. [Google Scholar] [CrossRef]
- Goodkind, M.; Eickhoff, S.B.; Oathes, D.J.; Jiang, Y.; Chang, A.; Jones-Hagata, L.B.; Ortega, B.N.; Zaiko, Y.V.; Roach, E.L.; Korgaonkar, M.S.; et al. Identification of a common neurobiological substrate for mental illness. JAMA Psychiatry 2015, 72, 305–315. [Google Scholar] [CrossRef]
- Sporns, O. From simple graphs to the connectome: Networks in neuroimaging. Neuroimage 2012, 62, 881–886. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; Bullmore, E.T.; Sporns, O. Comparative Connectomics. Trends Cogn. Sci. 2016, 20, 345–361. [Google Scholar]
- Menon, V. Large-scale brain networks and psychopathology: A unifying triple network model. Trends Cogn. Sci. 2011, 15, 483–506. [Google Scholar] [CrossRef]
- Hyman, S. Mental health: Depression needs large human-genetics studies. Nature 2014, 515, 189–191. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, P.F. Genetics of disease: Associations with depression. Nature 2015, 523, 539–540. [Google Scholar] [CrossRef] [Green Version]
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdellaoui, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.F.M.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [Green Version]
- Han, M.H.; Nestler, E.J. Neural Substrates of Depression and Resilience. Neurotherapeutics 2017, 14, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Caspi, A.; Moffitt, T.E. Gene-environment interactions in psychiatry: Joining forces with neuroscience. Nat. Rev. Neurosci. 2006, 7, 583–590. [Google Scholar] [CrossRef]
- Hollander, J.A.; Cory-Slechta, D.A.; Jacka, F.N.; Szabo, S.T.; Guilarte, T.R.; Bilbo, S.D.; Mattingly, C.J.; Moy, S.S.; Haroon, E.; Hornig, M.; et al. Beyond the looking glass: Recent advances in understanding the impact of environmental exposures on neuropsychiatric disease. Neuropsychopharmacology 2020, 45, 1086–1096. [Google Scholar] [CrossRef] [Green Version]
- Conway, F.; Brown, A.S. Maternal Immune Activation and Related Factors in the Risk of Offspring Psychiatric Disorders. Front. Psychiatry 2019, 10, 430. [Google Scholar] [CrossRef]
- Hay, D.F.; Pawlby, S.; Waters, C.S.; Sharp, D. Antepartum and postpartum exposure to maternal depression: Different effects on different adolescent outcomes. J. Child Psychol. Psychiatry 2008, 49, 1079–1088. [Google Scholar] [CrossRef]
- Fergusson, D.M.; Woodward, L.J.; Horwood, L.J. Maternal smoking during pregnancy and psychiatric adjustment in late adolescence. Arch. Gen. Psychiatry 1998, 55, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Allen, N.B.; Lewinsohn, P.M.; Seeley, J.R. Prenatal and perinatal influences on risk for psychopathology in childhood and adolescence. Dev. Psychopathol. 1998, 10, 513–529. [Google Scholar] [CrossRef]
- Jerrell, J.M.; McIntyre, R.S.; Tripathi, A. A cohort study of the prevalence and impact of comorbid medical conditions in pediatric bipolar disorder. J. Clin. Psychiatry 2010, 71, 1518–1525. [Google Scholar] [CrossRef]
- Wiwanitkit, V.; Jerrell, J.M.; McIntyre, R.S.; Tripathi, A. Cardiometabolic conditions in patients with Schizophrenia. Clin. Schizophr. Relat. Psychoses 2011, 5, 54. [Google Scholar] [CrossRef]
- McCoy, T.H.; Yu, S.; Hart, K.L.; Castro, V.M.; Brown, H.E.; Rosenquist, J.N.; Doyle, A.E.; Vuijk, P.J.; Cai, T.; Perlis, R.H. High Throughput Phenotyping for Dimensional Psychopathology in Electronic Health Records. Biol. Psychiatry 2018, 83, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Anderzhanova, E.; Kirmeier, T.; Wotjak, C.T. Animal models in psychiatric research: The RDoC system as a new framework for endophenotype-oriented translational neuroscience. Neurobiol. Stress 2017, 7, 47–56. [Google Scholar] [CrossRef]
- McEwen, B.S. Allostasis and allostatic load: Implications for neuropsychopharmacology. Neuropsychopharmacology 2000, 22, 108–124. [Google Scholar] [CrossRef]
- Purves, D.; Augustine, G.; Fitzpatrick, D.; Hall, W.; LaMantia, A.; White, L.; Mooney, R.; Platt, M. Neuroscience; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar]
- Jankord, R.; Herman, J.P. Limbic regulation of hypothalamo-pituitary-adrenocortical function during acute and chronic stress. Ann. N. Y. Acad. Sci. 2008, 1148, 64–73. [Google Scholar] [CrossRef]
- Jansen, A.S.; Nguyen, X.V.; Karpitskiy, V.; Mettenleiter, T.C.; Loewy, A.D. Central command neurons of the sympathetic nervous system: Basis of the fight-or-flight response. Science 1995, 270, 644–646. [Google Scholar] [CrossRef]
- Kim, J.S.; Iremonger, K.J. Temporally Tuned Corticosteroid Feedback Regulation of the Stress Axis. Trends Endocrinol. Metab. 2019, 30, 783–792. [Google Scholar] [CrossRef]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [Green Version]
- Lutz, B.; Marsicano, G.; Maldonado, R.; Hillard, C.J. The endocannabinoid system in guarding against fear, anxiety and stress. Nat. Rev. Neurosci. 2015, 16, 705–718. [Google Scholar] [CrossRef]
- deRoon-Cassini, T.A.; Stollenwerk, T.M.; Beatka, M.; Hillard, C.J. Meet Your Stress Management Professionals: The Endocannabinoids. Trends Mol. Med. 2020, in press. [Google Scholar] [CrossRef]
- Rusconi, F.; Rubino, T.; Battaglioli, E. Endocannabinoid-Epigenetic Cross-Talk: A Bridge toward Stress Coping. Int. J. Mol. Sci. 2020, 21, 6252. [Google Scholar] [CrossRef]
- Longaretti, A.; Forastieri, C.; Gabaglio, M.; Rubino, T.; Battaglioli, E.; Rusconi, F. Termination of acute stress response by the endocannabinoid system is regulated through LSD1-mediated transcriptional repression of 2-AG hydrolases ABHD6 and MAGL. J. Neurochem. 2020, 155, 98–110. [Google Scholar] [CrossRef]
- Rusconi, F.; Battaglioli, E. Acute Stress-Induced Epigenetic Modulations and Their Potential Protective Role Toward Depression. Front. Mol. Neurosci. 2018, 11, 184. [Google Scholar] [CrossRef]
- Rusconi, F.; Grillo, B.; Toffolo, E.; Mattevi, A.; Battaglioli, E. NeuroLSD1: Splicing-Generated Epigenetic Enhancer of Neuroplasticity. Trends Neurosci. 2017, 40, 28–38. [Google Scholar] [CrossRef]
- Rusconi, F.; Grillo, B.; Ponzoni, L.; Bassani, S.; Toffolo, E.; Paganini, L.; Mallei, A.; Braida, D.; Passafaro, M.; Popoli, M.; et al. LSD1 modulates stress-evoked transcription of immediate early genes and emotional behavior. Proc. Natl. Acad. Sci. USA 2016, 113, 3651–3656. [Google Scholar] [CrossRef] [Green Version]
- Prini, P.; Rusconi, F.; Zamberletti, E.; Gabaglio, M.; Penna, F.; Fasano, M.; Battaglioli, E.; Parolaro, D.; Rubino, T. Adolescent THC exposure in female rats leads to cognitive deficits through a mechanism involving chromatin modifications in the prefrontal cortex. J. Psychiatry Neurosci. 2018, 43, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Rusconi, F.; Battaglioli, E.; Venturin, M. Psychiatric Disorders and lncRNAs: A Synaptic Match. Int. J. Mol. Sci. 2020, 21, 3030. [Google Scholar] [CrossRef]
- Guggenhuber, S.; Romo-Parra, H.; Bindila, L.; Leschik, J.; Lomazzo, E.; Remmers, F.; Zimmermann, T.; Lerner, R.; Klugmann, M.; Pape, H.-C.; et al. Impaired 2-AG Signaling in Hippocampal Glutamatergic Neurons: Aggravation of Anxiety-Like Behavior and Unaltered Seizure Susceptibility. Int. J. Neuropsychopharmacol. 2015, 19, pyv091. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Shonesy, B.C.; Bluett, R.J.; Winder, D.G.; Colbran, R.J. The Anxiolytic Actions of 2-Arachidonoylglycerol: Converging Evidence From Two Recent Genetic Endocannabinoid Deficiency Models. Biol. Psychiatry 2016, 79, e78–e79. [Google Scholar] [CrossRef]
- McEwen, B.S. Effects of adverse experiences for brain structure and function. Biol. Psychiatry 2000, 48, 721–731. [Google Scholar] [CrossRef]
- McEwen, B.S. Mood disorders and allostatic load. Biol. Psychiatry 2003, 54, 200–207. [Google Scholar] [CrossRef]
- Copstead, L.-E.; Banasik, J. Pathophysiology; Saunders: Philadeplohia, PA, USA, 2012; p. 1216. [Google Scholar]
- Joëls, M.; Baram, T.Z. The neuro-symphony of stress. Nat. Rev. Neurosci. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Musazzi, L.; Tornese, P.; Sala, N.; Popoli, M. Acute or Chronic? A Stressful Question. Trends Neurosci. 2017, 40, 525–535. [Google Scholar] [CrossRef]
- Gold, P.W.; Machado-Vieira, R.; Pavlatou, M.G. Clinical and biochemical manifestations of depression: Relation to the neurobiology of stress. Neural Plast. 2015, 2015, 581976. [Google Scholar] [CrossRef]
- Gold, P.W. The organization of the stress system and its dysregulation in depressive illness. Mol. Psychiatry 2015, 20, 32–47. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bagot, R.C.; Labonté, B.; Peña, C.J.; Nestler, E.J. Epigenetic signaling in psychiatric disorders: Stress and depression. Dialogues Clin. Neurosci. 2014, 16, 281–295. [Google Scholar]
- Overstreet, D.H. Modeling depression in animal models. Methods Mol. Biol. 2012, 829, 125–144. [Google Scholar]
- Wang, Q.; Timberlake, M.A.; Prall, K.; Dwivedi, Y. The recent progress in animal models of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 77, 99–109. [Google Scholar] [CrossRef]
- Ménard, C.; Hodes, G.E.; Russo, S.J. Pathogenesis of depression: Insights from human and rodent studies. Neuroscience 2016, 321, 138–162. [Google Scholar] [CrossRef] [Green Version]
- Czéh, B.; Fuchs, E.; Wiborg, O.; Simon, M. Animal models of major depression and their clinical implications. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 293–310. [Google Scholar] [CrossRef]
- Belzung, C.; Lemoine, M. Criteria of validity for animal models of psychiatric disorders: Focus on anxiety disorders and depression. Biol. Mood Anxiety Disord. 2011, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- van der Staay, F.J.; Arndt, S.S.; Nordquist, R.E. Evaluation of animal models of neurobehavioral disorders. Behav. Brain Funct. 2009, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Gururajan, A.; Reif, A.; Cryan, J.F.; Slattery, D.A. The future of rodent models in depression research. Nat. Rev. Neurosci. 2019, 20, 686–701. [Google Scholar] [CrossRef]
- Richter-Levin, G.; Stork, O.; Schmidt, M.V. Animal models of PTSD: A challenge to be met. Mol. Psychiatry 2018, 24, 1135–1156. [Google Scholar] [CrossRef] [Green Version]
- Pryce, C.R.; Fuchs, E. Chronic psychosocial stressors in adulthood: Studies in mice, rats and tree shrews. Neurobiol. Stress 2017, 6, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Schöner, J.; Heinz, A.; Endres, M.; Gertz, K.; Kronenberg, G. Post-traumatic stress disorder and beyond: An overview of rodent stress models. J. Cell. Mol. Med. 2017, 21, 2248–2256. [Google Scholar] [CrossRef] [Green Version]
- Zanette, L.Y.; Hobbs, E.C.; Witterick, L.E.; MacDougall-Shackleton, S.A.; Clinchy, M. Predator-induced fear causes PTSD-like changes in the brains and behaviour of wild animals. Sci. Rep. 2019, 9, 11474. [Google Scholar] [CrossRef]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Schroeder, A.; Notaras, M.; Du, X.; Hill, R.A. On the Developmental Timing of Stress: Delineating Sex-Specific Effects of Stress across Development on Adult Behavior. Brain Sci. 2018, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, D.; Knoll, L.J.; Blakemore, S.J. Adolescence as a Sensitive Period of Brain Development. Trends Cogn. Sci. 2015, 19, 558–566. [Google Scholar]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [Green Version]
- Kaczkurkin, A.N.; Burton, P.C.; Chazin, S.M.; Manbeck, A.B.; Espensen-Sturges, T.; Cooper, S.E.; Sponheim, S.R.; Lissek, S. Neural Substrates of Overgeneralized Conditioned Fear in PTSD. Am. J. Psychiatry 2017, 174, 125–134. [Google Scholar] [CrossRef]
- Blacker, C.J.; Frye, M.A.; Morava, E.; Kozicz, T.; Veldic, M. A Review of Epigenetics of PTSD in Comorbid Psychiatric Conditions. Genes 2019, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Curzon, P.; Rustay, N.; Browman, K. Cued and Contextual Fear Conditioning for Rodents. In Methods of behaviour analysis in neuroscience; Buccafusco, J., Ed.; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Spadaro, P.A.; Flavell, C.R.; Widagdo, J.; Ratnu, V.S.; Troup, M.; Ragan, C.; Mattick, J.S.; Bredy, T.W. Long Noncoding RNA-Directed Epigenetic Regulation of Gene Expression Is Associated With Anxiety-like Behavior in Mice. Biol. Psychiatry 2015, 78, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Bali, A.; Jaggi, A.S. Electric foot shock stress: A useful tool in neuropsychiatric studies. Rev. Neurosci. 2015, 26, 655–677. [Google Scholar] [CrossRef]
- Gräff, J.; Joseph, N.F.; Horn, M.E.; Samiei, A.; Meng, J.; Seo, J.; Rei, D.; Bero, A.W.; Phan, T.X.; Wagner, F.; et al. Epigenetic priming of memory updating during reconsolidation to attenuate remote fear memories. Cell 2014, 156, 261–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Kim, S.Y.; Artis, S.; Molfese, D.L.; Schumacher, A.; Sweatt, J.D.; Paylor, R.E.; Lubin, F.D. Histone methylation regulates memory formation. J. Neurosci. 2010, 30, 3589–3599. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, J.M.; Malleret, G.; Touzani, K.; Vronskaya, S.; Ishii, S.; Kandel, E.R.; Barco, A. Chromatin acetylation, memory, and LTP are impaired in CBP+/- mice: A model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron 2004, 42, 947–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.-S.; Haggarty, S.J.; Giacometti, E.; Dannenberg, J.-H.; Joseph, N.; Gao, J.; Nieland, T.J.F.; Zhou, Y.; Wang, X.; Mazitschek, R.; et al. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature 2009, 459, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Casacchia, M.; Bianchini, V.; Mazza, M.; Pollice, R.; Roncone, R. Acute stress reactions and associated factors in the help-seekers after the L’Aquila earthquake. Psychopathology 2013, 46, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Pollice, R.; Bianchini, V.; Di Mauro, S.; Mazza, M.; Verni, L.; Roncone, R.; Casacchia, M. Cognitive function and clinical symptoms in first-episode psychosis and chronic schizophrenia before and after the 2009 L’Aquila earthquake. Early Interv. Psychiatry 2012, 6, 153–158. [Google Scholar] [CrossRef]
- Sadler, A.M.; Bailey, S.J. Repeated daily restraint stress induces adaptive behavioural changes in both adult and juvenile mice. Physiol. Behav. 2016, 167, 313–323. [Google Scholar] [CrossRef]
- Chiba, S.; Numakawa, T.; Ninomiya, M.; Richards, M.C.; Wakabayashi, C.; Kunugi, H. Chronic restraint stress causes anxiety- and depression-like behaviors, downregulates glucocorticoid receptor expression, and attenuates glutamate release induced by brain-derived neurotrophic factor in the prefrontal cortex. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 112–119. [Google Scholar] [CrossRef]
- Maghsoudi, N.; Ghasemi, R.; Ghaempanah, Z.; Ardekani, A.M.; Nooshinfar, E.; Tahzibi, A. Effect of Chronic Restraint Stress on HPA Axis Activity and Expression of BDNF and Trkb in the Hippocampus of Pregnant Rats: Possible Contribution in Depression during Pregnancy and Postpartum Period. Basic Clin. Neurosci. 2014, 5, 131–137. [Google Scholar]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477–501. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Zhou, Y.; Hu, Z.; Lou, J.; Song, W.; Li, J.; Liang, X.; Chen, C.; Wang, S.; Yang, B.; et al. 24-hour-restraint stress induces long-term depressive-like phenotypes in mice. Sci. Rep. 2016, 6, 32935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Hillard, C.J. Adaptations in endocannabinoid signaling in response to repeated homotypic stress: A novel mechanism for stress habituation. Eur. J. Neurosci. 2008, 27, 2821–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kara, N.Z.; Stukalin, Y.; Einat, H. Revisiting the validity of the mouse forced swim test: Systematic review and meta-analysis of the effects of prototypic antidepressants. Neurosci. Biobehav. Rev. 2018, 84, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.L.; Deak, T. A users guide to HPA axis research. Physiol. Behav. 2017, 178, 43–65. [Google Scholar] [CrossRef]
- Abel, E.L. Physiological correlates of the forced swim test in rats. Physiol. Behav. 1993, 54, 309–317. [Google Scholar] [CrossRef]
- Connor, T.J.; Kelly, J.P.; Leonard, B.E. Forced swim test-induced neurochemical endocrine, and immune changes in the rat. Pharmacol. Biochem. Behav. 1997, 58, 961–967. [Google Scholar] [CrossRef]
- Mul, J.D.; Zheng, J.; Goodyear, L.J. Validity Assessment of 5 Day Repeated Forced-Swim Stress to Model Human Depression in Young-Adult C57BL/6J and BALB/cJ Mice. eNeuro 2016, 3, e0201-16.2016. [Google Scholar] [CrossRef] [Green Version]
- de Kloet, E.R.; Molendijk, M.L. Coping with the Forced Swim Stressor: Towards Understanding an Adaptive Mechanism. Neural Plast. 2016, 2016, 6503162. [Google Scholar] [CrossRef] [Green Version]
- Slattery, D.A.; Cryan, J.F. Using the rat forced swim test to assess antidepressant-like activity in rodents. Nat. Protoc. 2012, 7, 1009–1014. [Google Scholar] [CrossRef]
- Chen, X.-N.; Meng, Q.-Y.; Bao, A.-M.; Swaab, D.F.; Wang, G.-H.; Zhou, J.-N. The involvement of retinoic acid receptor-alpha in corticotropin-releasing hormone gene expression and affective disorders. Biol. Psychiatry 2009, 66, 832–839. [Google Scholar] [CrossRef]
- Wang, S.S.; Yan, X.B.; Hofman, M.A.; Swaab, D.F.; Zhou, J.N. Increased expression level of corticotropin-releasing hormone in the amygdala and in the hypothalamus in rats exposed to chronic unpredictable mild stress. Neurosci. Bull. 2010, 26, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayensu, W.K.; Pucilowski, O.; Mason, G.A.; Overstreet, D.H.; Rezvani, A.H.; Janowsky, D.S. Effects of chronic mild stress on serum complement activity, saccharin preference, and corticosterone levels in Flinders lines of rats. Physiol. Behav. 1995, 57, 165–169. [Google Scholar] [CrossRef]
- Duniec, E.; Raz, M. Vitamins for the soul: John Bowlby’s thesis of maternal deprivation, biomedical metaphors and the deficiency model of disease. Hist. Psychiatry 2011, 22, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Bowlby, J. Maternal Care and Mental Health; World Health Organization: Geneva, Switzerland, 1951; pp. 355–534. [Google Scholar]
- Bowlby, J. A note on mother-child separation as a mental health hazard. Br. J. Med. Psychol. 1958, 31, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, J.S. The evolutionary ecology of attachment organization. Hum. Nat. 1996, 7, 1–37. [Google Scholar] [PubMed]
- Petti, T.A.; Salguero, C. A Manual of Clinical Practice and Consultation; American Psychiatric Publishing, Inc.: Arlington, VA, USA, 2006. [Google Scholar]
- Janetsian-Fritz, S.S.; Timme, N.M.; Timm, M.M.; McCane, A.M.; Baucum, A.J., II; O’Donnell, B.F.; Lapish, C.C. Maternal deprivation induces alterations in cognitive and cortical function in adulthood. Transl. Psychiatry 2018, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Marco, E.M.; Llorente, R.; López-Gallardo, M.; Mela, V.; Llorente-Berzal, Á.; Prada, C.; Viveros, M.P. The maternal deprivation animal model revisited. Neurosci. Biobehav. Rev. 2015, 51, 151–163. [Google Scholar] [CrossRef]
- Schmidt, M.V.; Enthoven, L.; Van Woezik, J.H.G.; Levine, S.; De Kloet, E.R.; Oitzl, M.S. The dynamics of the hypothalamic-pituitary-adrenal axis during maternal deprivation. J. Neuroendocrinol. 2004, 16, 52–57. [Google Scholar] [CrossRef]
- Kim, J.J.; Diamond, D.M. The stressed hippocampus, synaptic plasticity and lost memories. Nat. Rev. Neurosci. 2002, 3, 453–462. [Google Scholar] [CrossRef]
- Singh, A.; Misra, N. Loneliness, depression and sociability in old age. Ind. Psychiatry J. 2009, 18, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, R.; Shoib, S.; Shah, T.; Mushtaq, S. Relationship between loneliness, psychiatric disorders and physical health ? A review on the psychological aspects of loneliness. J. Clin. Diagn. Res. 2014, 8, WE01–WE04. [Google Scholar] [CrossRef] [PubMed]
- Orben, A.; Tomova, L.; Blakemore, S.J. The effects of social deprivation on adolescent development and mental health. Lancet Child Adolesc. Health 2020, 4, 634–640. [Google Scholar] [CrossRef]
- Sourander, A.; Klomek, A.B.; Ikonen, M.; Lindroos, J.; Luntamo, T.; Koskelainen, M.; Ristkari, T.; Helenius, H. Psychosocial risk factors associated with cyberbullying among adolescents: A population-based study. Arch. Gen. Psychiatry 2010, 67, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Teo, A.R.; Gaw, A.C. Hikikomori, a Japanese culture-bound syndrome of social withdrawal?: A proposal for DSM-5. J. Nerv. Ment. Dis. 2010, 198, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Malagón-Amor, Á.; Martín-López, L.M.; Córcoles, D.; González, A.; Bellsolà, M.; Teo, A.R.; Bulbena, A.; Pérez, V.; Bergé, D. Family Features of Social Withdrawal Syndrome (Hikikomori). Front. Psychiatry 2020, 11, 138. [Google Scholar] [CrossRef] [Green Version]
- Yanguas, J.; Pinazo-Henandis, S.; Tarazona-Santabalbina, F.J. The complexity of loneliness. Acta Biomed. 2018, 89, 302–314. [Google Scholar]
- Koike, H.; Ibi, D.; Mizoguchi, H.; Nagai, T.; Nitta, A.; Takuma, K.; Nabeshima, T.; Yoneda, Y.; Yamada, K. Behavioral abnormality and pharmacologic response in social isolation-reared mice. Behav. Brain Res. 2009, 202, 114–121. [Google Scholar] [CrossRef]
- Kim, J.W.; Kirkpatrick, B. Social isolation in animal models of relevance to neuropsychiatric disorders. Biol. Psychiatry 1996, 40, 918–922. [Google Scholar] [CrossRef]
- Ieraci, A.; Mallei, A.; Popoli, M. Social Isolation Stress Induces Anxious-Depressive-Like Behavior and Alterations of Neuroplasticity-Related Genes in Adult Male Mice. Neural Plast. 2016, 2016, 6212983. [Google Scholar] [CrossRef] [Green Version]
- Boero, G.; Pisu, M.G.; Biggio, F.; Muredda, L.; Carta, G.; Banni, S.; Paci, E.; Follesa, P.; Concas, A.; Porcu, P.; et al. Impaired glucocorticoid-mediated HPA axis negative feedback induced by juvenile social isolation in male rats. Neuropharmacology 2018, 133, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Gillen, P.A.; Sinclair, M.; Kernohan, W.G.; Begley, C.M.; Luyben, A.G. Interventions for prevention of bullying in the workplace. Cochrane Database Syst. Rev. 2017, 1, CD009778. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.J.; Kendall, G.E.; Burns, S.K.; Schonert-Reichl, K.A.; Kane, R.T. Measuring 8 to 12 year old children’s self-report of power imbalance in relation to bullying: Development of the Scale of Perceived Power Imbalance. BMC Public Health 2019, 19, 1046. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.A.; Covington, H.E.; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 2011, 6, 1183–1191. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Zhou, X.-Y.; Yang, L.-N.; Wang, H.-Y.; Zhang, Y.-Q.; Pu, J.-C.; Liu, L.-X.; Gui, S.-W.; Zeng, L.; Chen, J.-J.; et al. Social defeat stress causes depression-like behavior with metabolite changes in the prefrontal cortex of rats. PLoS ONE 2017, 12, e0176725. [Google Scholar] [CrossRef] [Green Version]
- Keeney, A.; Jessop, D.S.; Harbuz, M.; Marsden, C.A.; Hogg, S.; Blackburn-Munro, R.E. Differential effects of acute and chronic social defeat stress on hypothalamic-pituitary-adrenal axis function and hippocampal serotonin release in mice. J. Neuroendocrinol. 2006, 18, 330–338. [Google Scholar] [CrossRef]
- Bagot, R.C.; Parise, E.M.; Pena, C.J.; Zhang, H.X.; Maze, I.; Chaudhury, D.; Persaud, B.; Cachope, R.; Bolaños-Guzmán, C.A.; Cheer, J.F.; et al. Ventral hippocampal afferents to the nucleus accumbens regulate susceptibility to depression. Nat. Commun. 2015, 6, 7062. [Google Scholar] [CrossRef]
- Gerosa, L.; Grillo, B.; Forastieri, C.; Longaretti, A.; Toffolo, E.; Mallei, A.; Bassani, S.; Popoli, M.; Battaglioli, E.; Rusconi, F. SRF and SRFΔ5 Splicing Isoform Recruit Corepressor LSD1/KDM1A Modifying Structural Neuroplasticity and Environmental Stress Response. Mol. Neurobiol. 2019, 57, 393–407. [Google Scholar] [CrossRef]
- Anacker, C.; Luna, V.M.; Stevens, G.S.; Millette, A.; Shores, R.; Jimenez, J.C.; Chen, B.; Hen, R. Hippocampal neurogenesis confers stress resilience by inhibiting the ventral dentate gyrus. Nature 2018, 559, 98–102. [Google Scholar] [CrossRef]
- Tsankova, N.M.; Berton, O.; Renthal, W.; Kumar, A.; Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef]
- Toyoda, A. Social defeat models in animal science: What we have learned from rodent models. Anim. Sci. J. 2017, 88, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archibald, K.; Clotworthy, M. Comment on ‘The ethics of animal research’ by Festing & Wilkinson. EMBO Rep. 2007, 8, 794–796. [Google Scholar] [PubMed]
- Festing, S.; Wilkinson, R. The ethics of animal research. Talking Point on the use of animals in scientific research. EMBO Rep. 2007, 8, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S. Biomarkers for assessing population and individual health and disease related to stress and adaptation. Metabolism 2015, 64, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Gray, J.; Nasca, C. Recognizing Resilience: Learning from the Effects of Stress on the Brain. Neurobiol. Stress 2015, 1, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-Y.; Jing, D.; Bath, K.G.; Ieraci, A.; Khan, T.; Siao, C.-J.; Herrera, D.G.; Toth, M.; Yang, C.; McEwen, B.S.; et al. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 2006, 314, 140–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Italia, M.; Forastieri, C.; Longaretti, A.; Battaglioli, E.; Rusconi, F. Rationale, Relevance, and Limits of Stress-Induced Psychopathology in Rodents as Models for Psychiatry Research: An Introductory Overview. Int. J. Mol. Sci. 2020, 21, 7455. https://doi.org/10.3390/ijms21207455
Italia M, Forastieri C, Longaretti A, Battaglioli E, Rusconi F. Rationale, Relevance, and Limits of Stress-Induced Psychopathology in Rodents as Models for Psychiatry Research: An Introductory Overview. International Journal of Molecular Sciences. 2020; 21(20):7455. https://doi.org/10.3390/ijms21207455
Chicago/Turabian StyleItalia, Maria, Chiara Forastieri, Alessandra Longaretti, Elena Battaglioli, and Francesco Rusconi. 2020. "Rationale, Relevance, and Limits of Stress-Induced Psychopathology in Rodents as Models for Psychiatry Research: An Introductory Overview" International Journal of Molecular Sciences 21, no. 20: 7455. https://doi.org/10.3390/ijms21207455
APA StyleItalia, M., Forastieri, C., Longaretti, A., Battaglioli, E., & Rusconi, F. (2020). Rationale, Relevance, and Limits of Stress-Induced Psychopathology in Rodents as Models for Psychiatry Research: An Introductory Overview. International Journal of Molecular Sciences, 21(20), 7455. https://doi.org/10.3390/ijms21207455