Resveratrol Rescue Indoxyl Sulfate-Induced Deterioration of Osteoblastogenesis via the Aryl Hydrocarbon Receptor /MAPK Pathway

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results
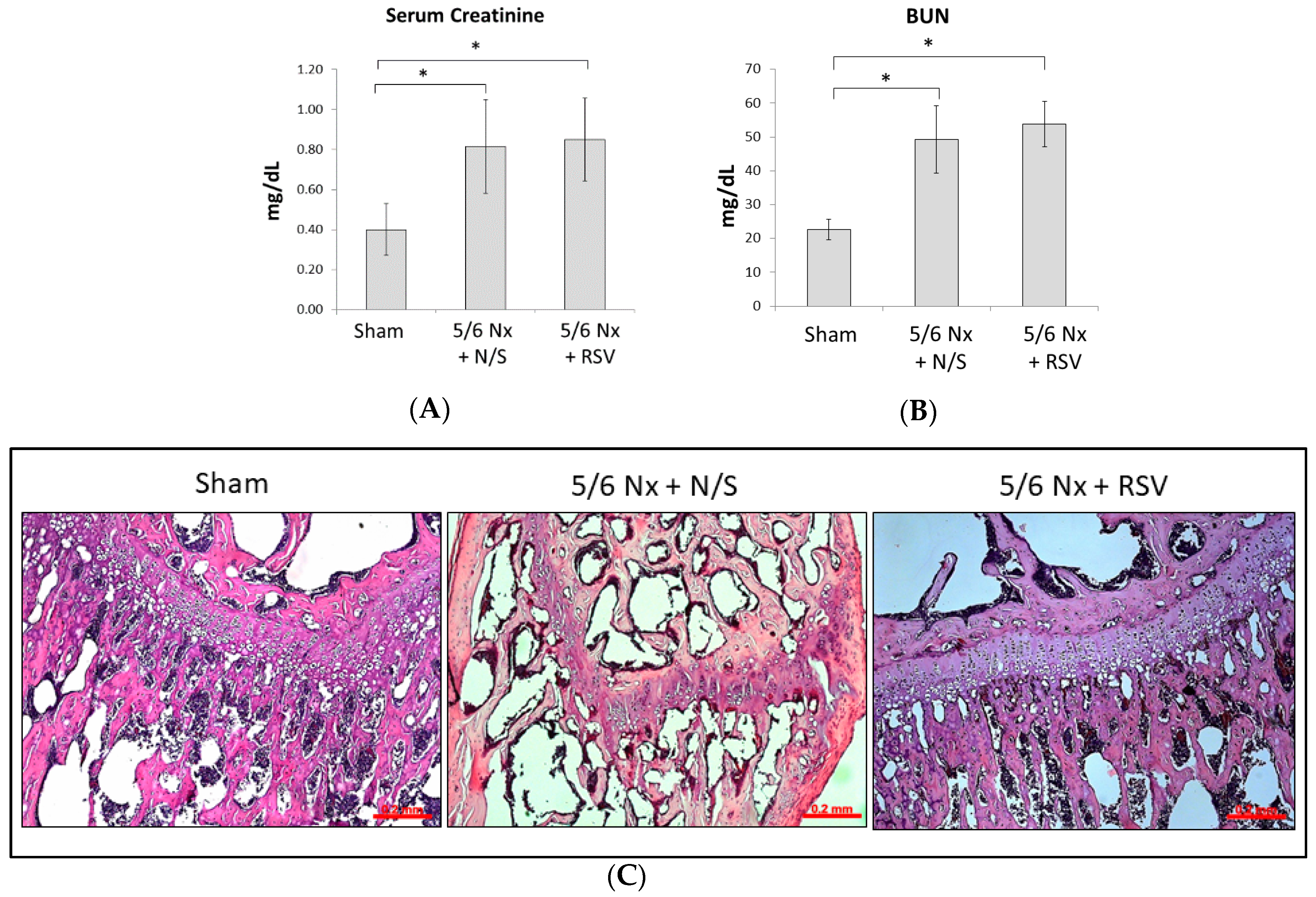
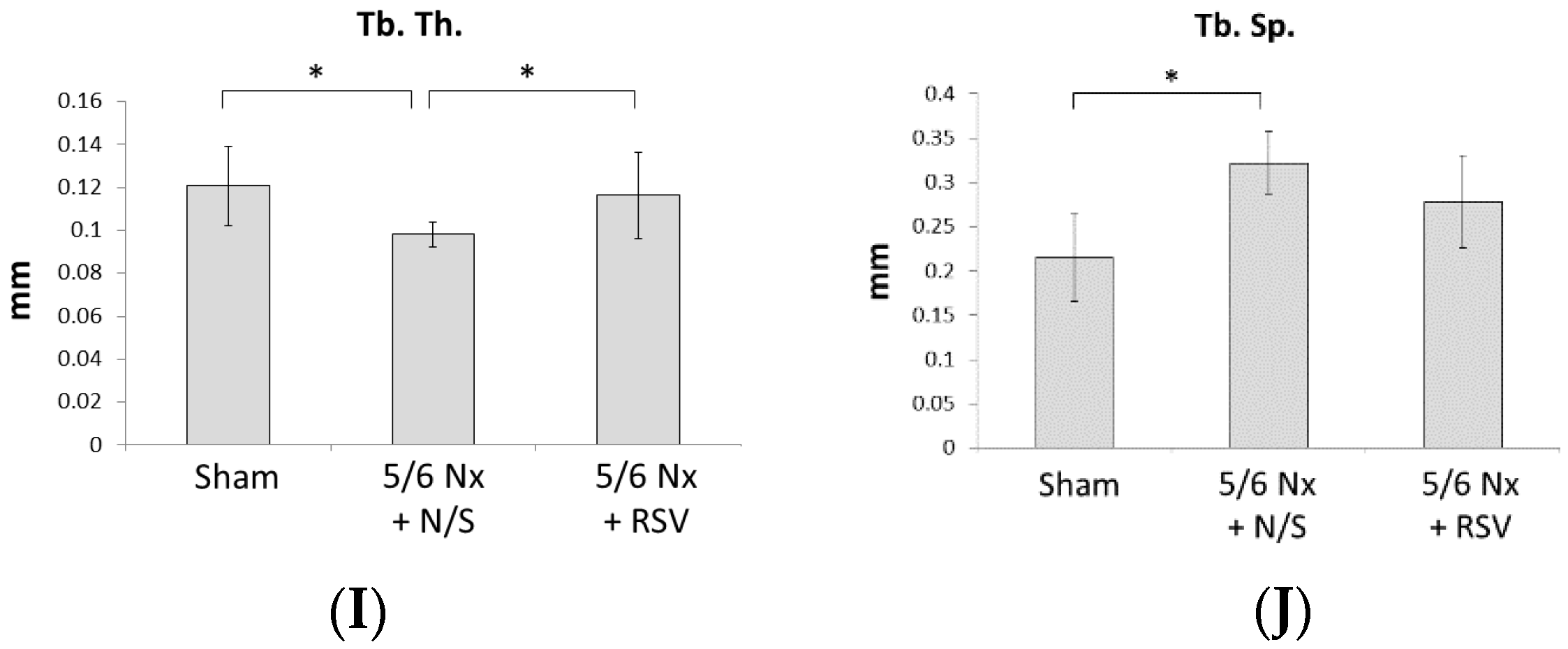
2.1. RSV Attenuates the Effect of Renal Function Insufficiency on Bone Formation In Vivo
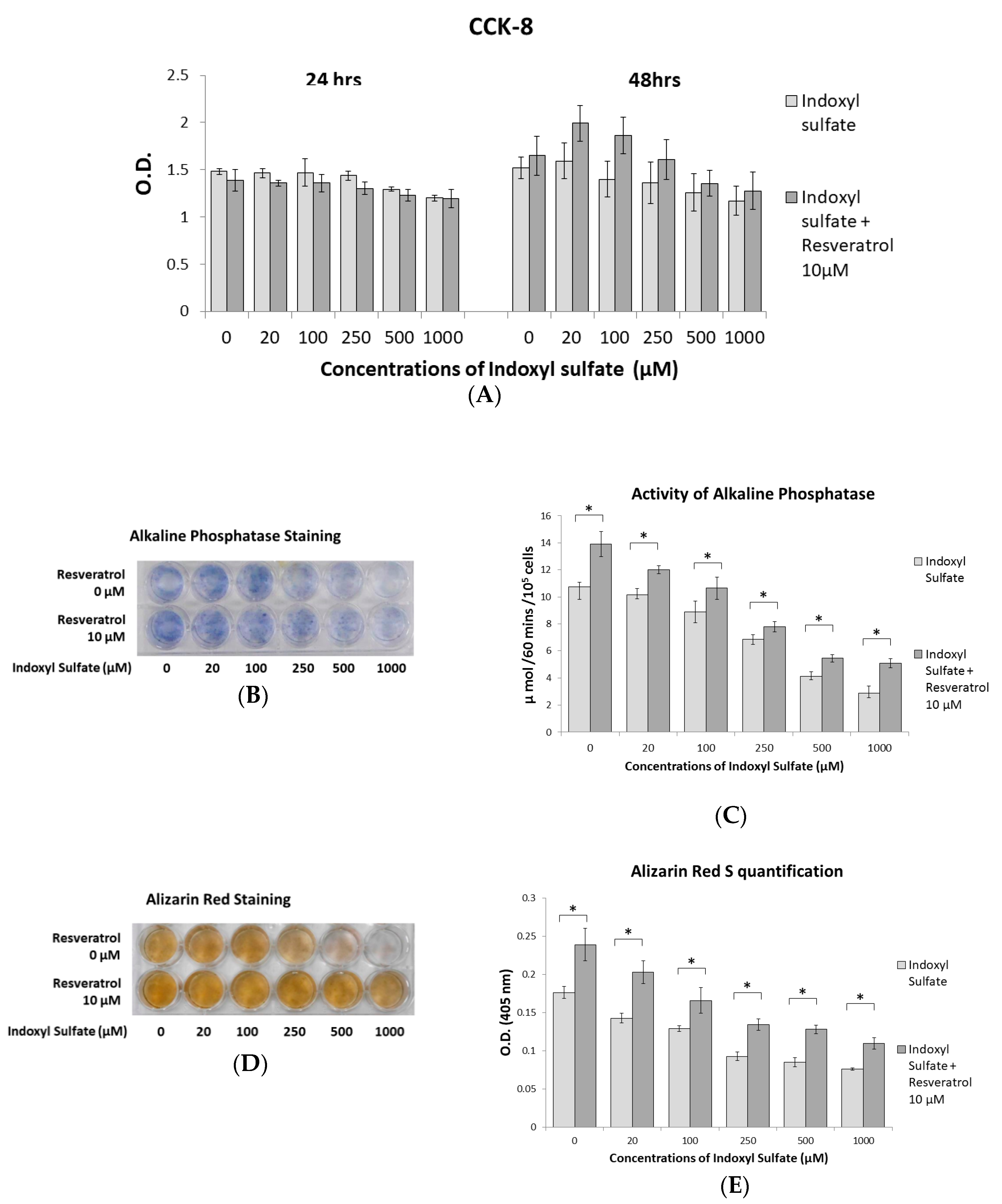
2.2. RSV Ameliorates Impaired Osteoblast Differentiation and Mineralization Induced by IS
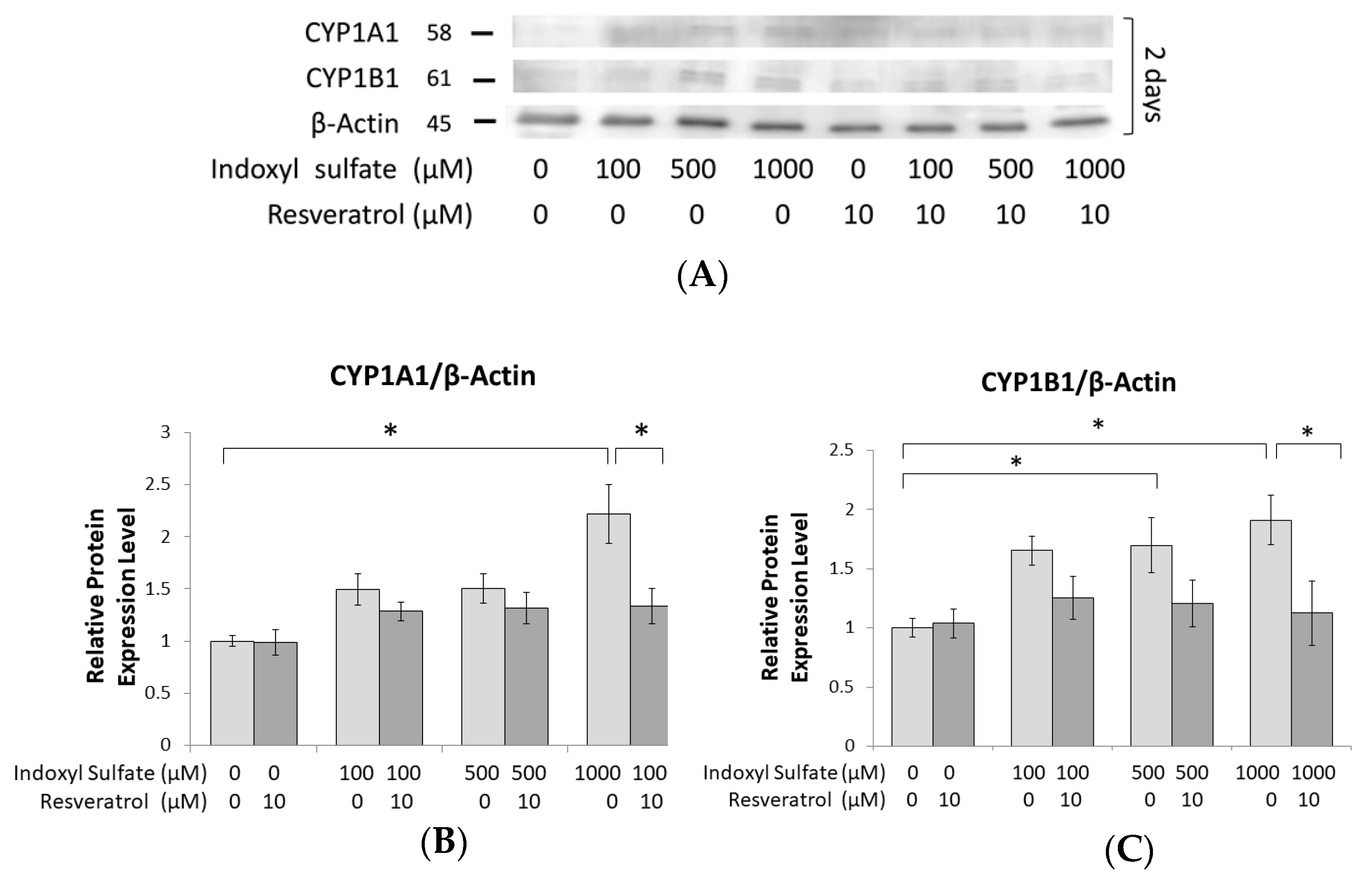
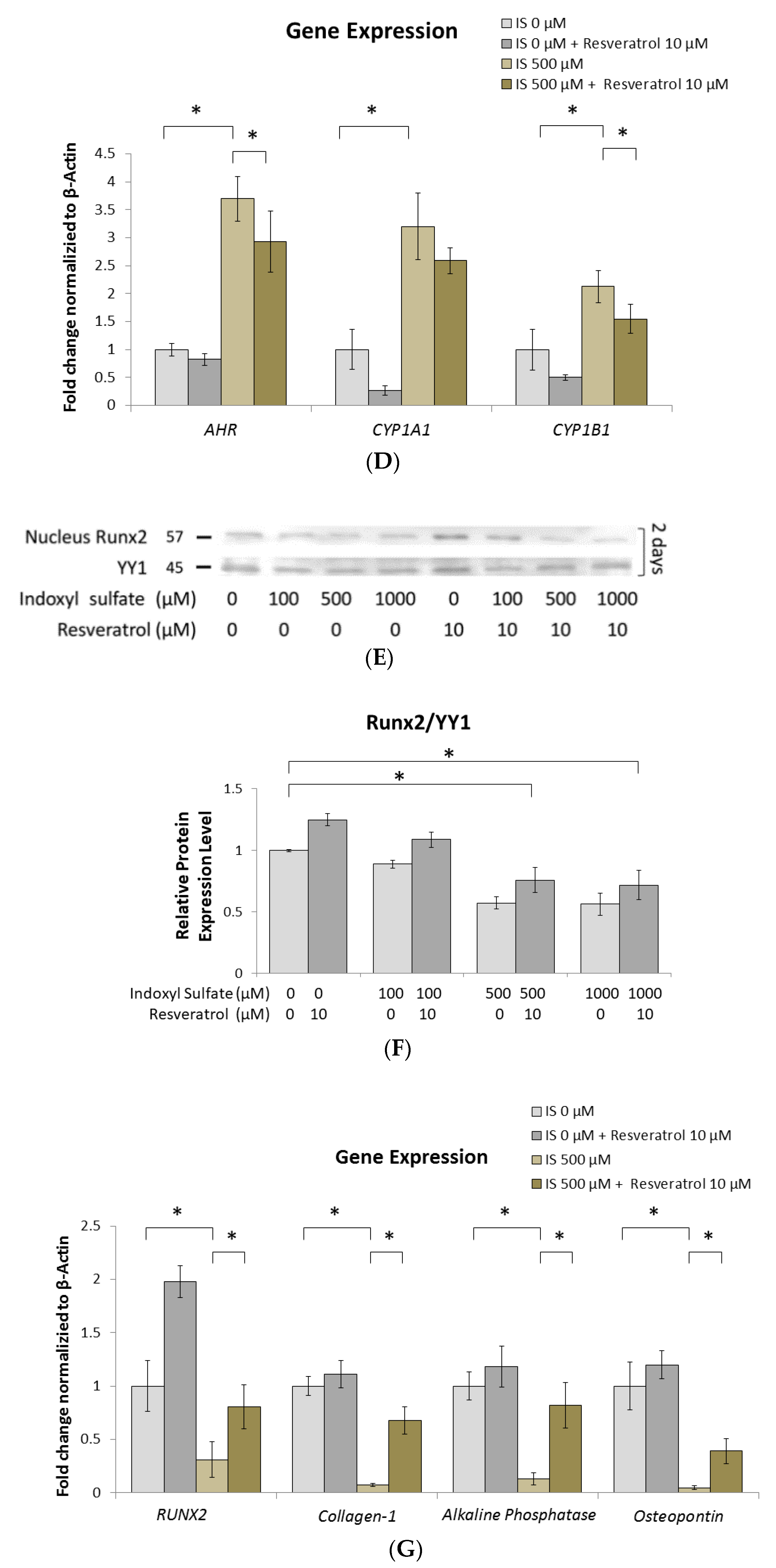
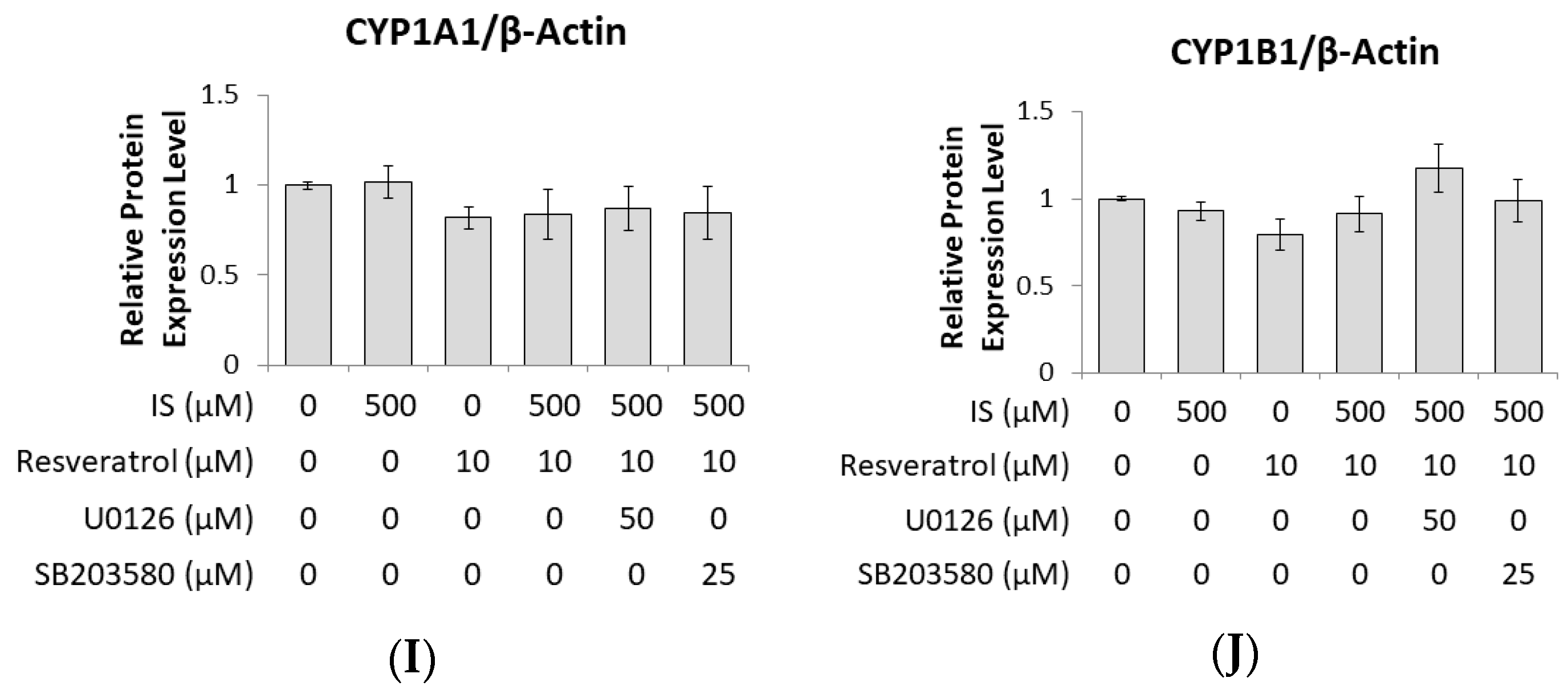
2.3. Prohibition of Osteogenesis Development through AhR Activated by IS Exposure
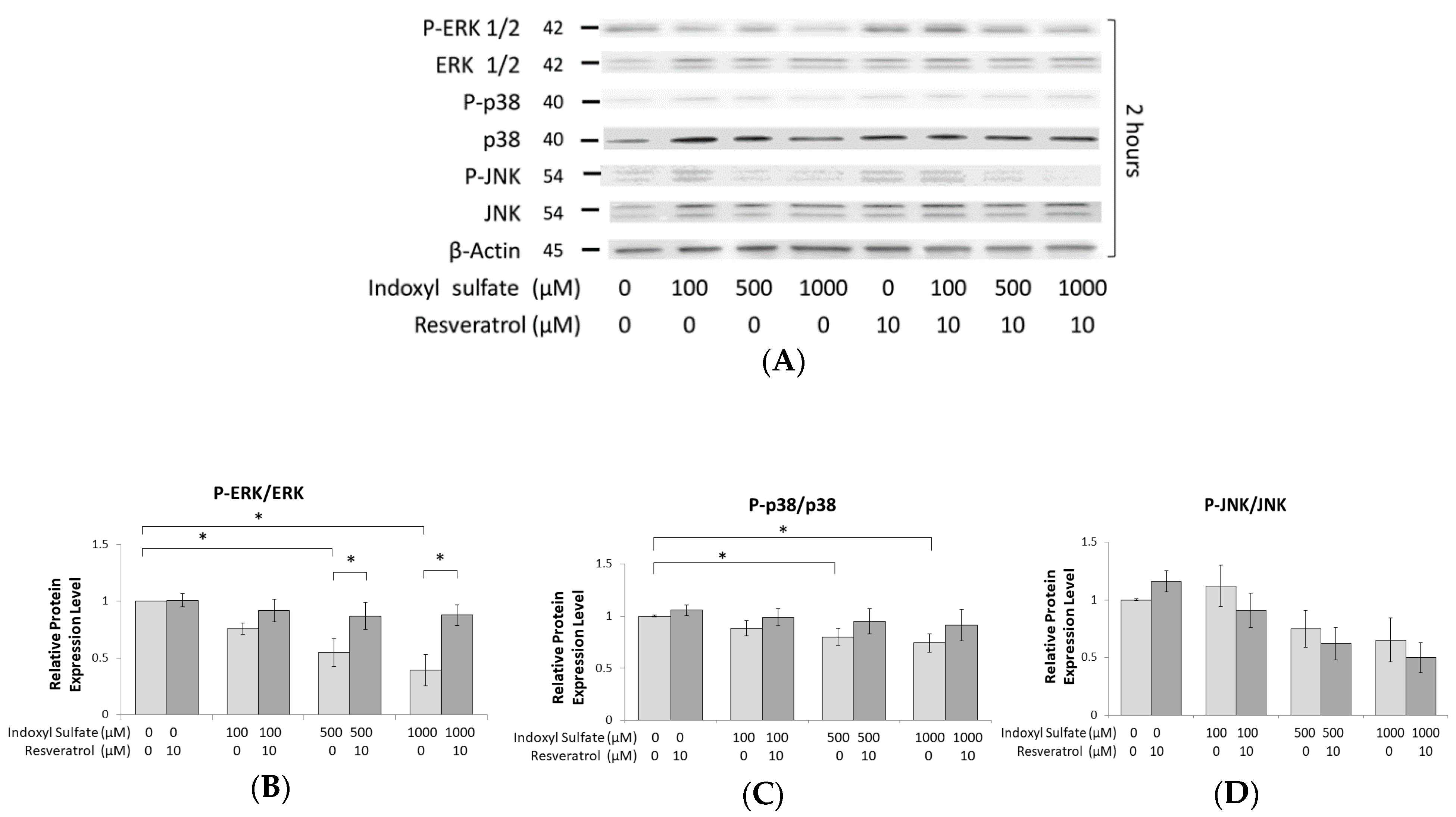
2.4. IS Inhibits the Phosphorylation of ERK1/2 and p38 MAPK
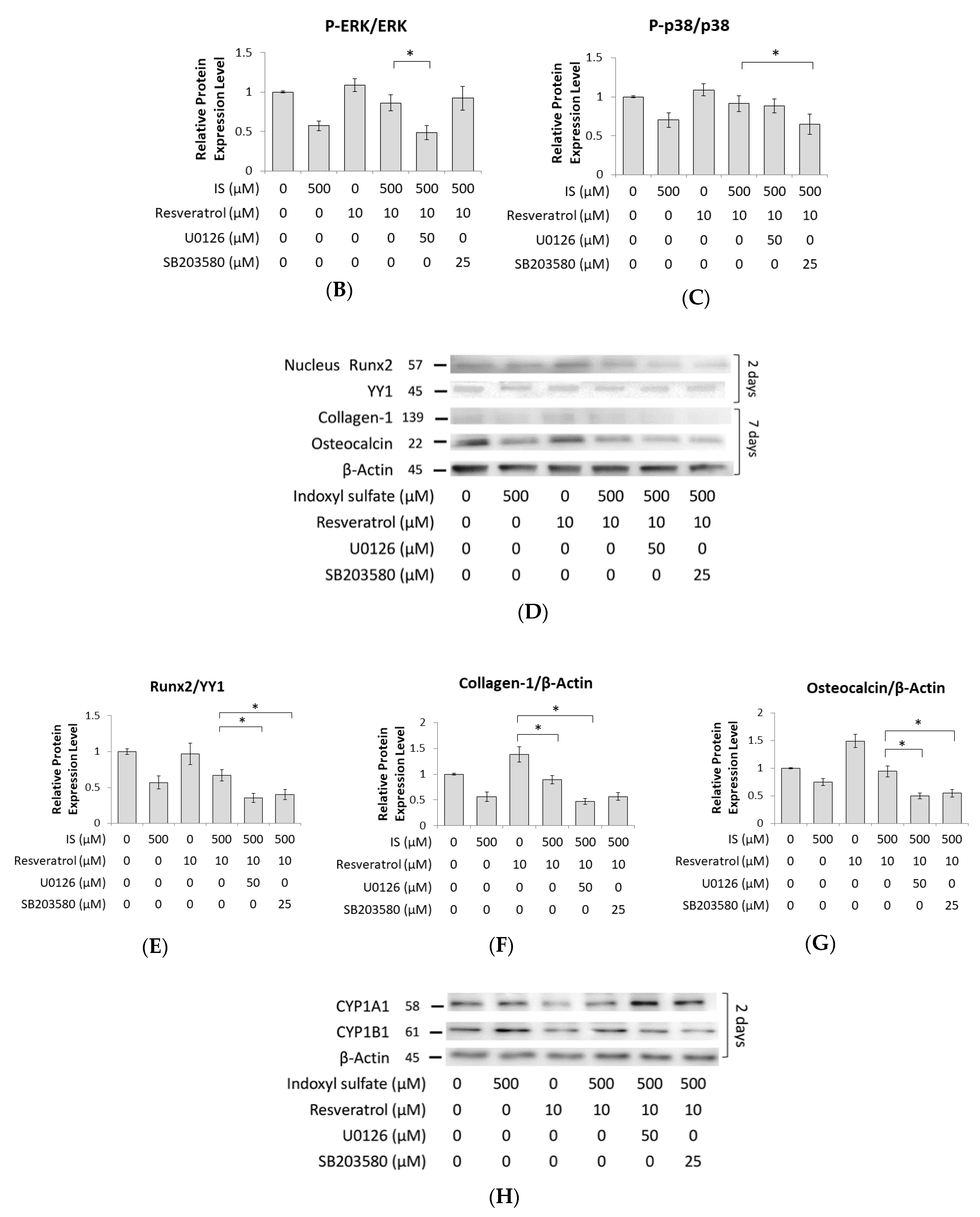
2.5. Role of ERK1/2 and p38 MAPK Signaling in Osteogenesis
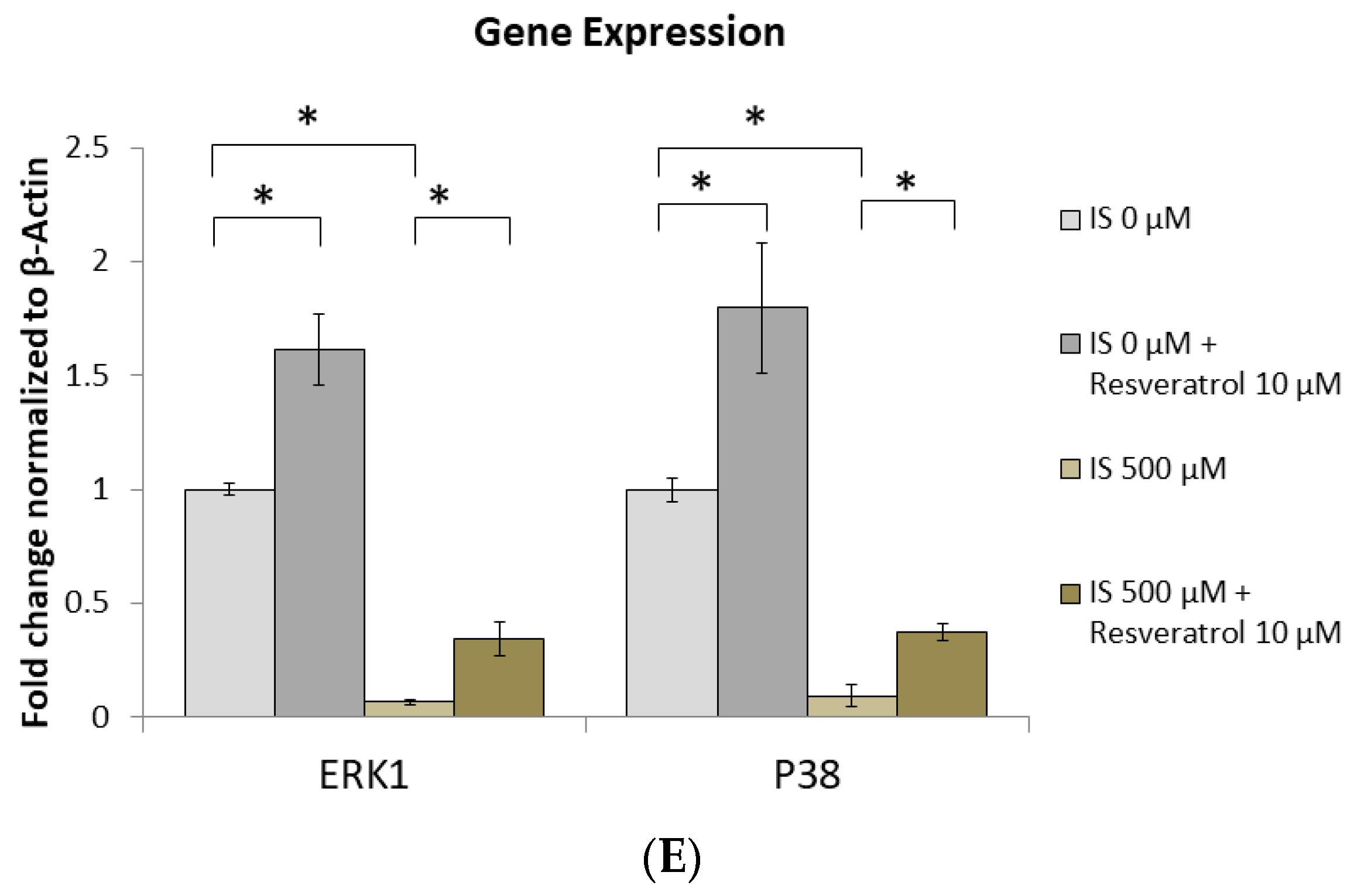
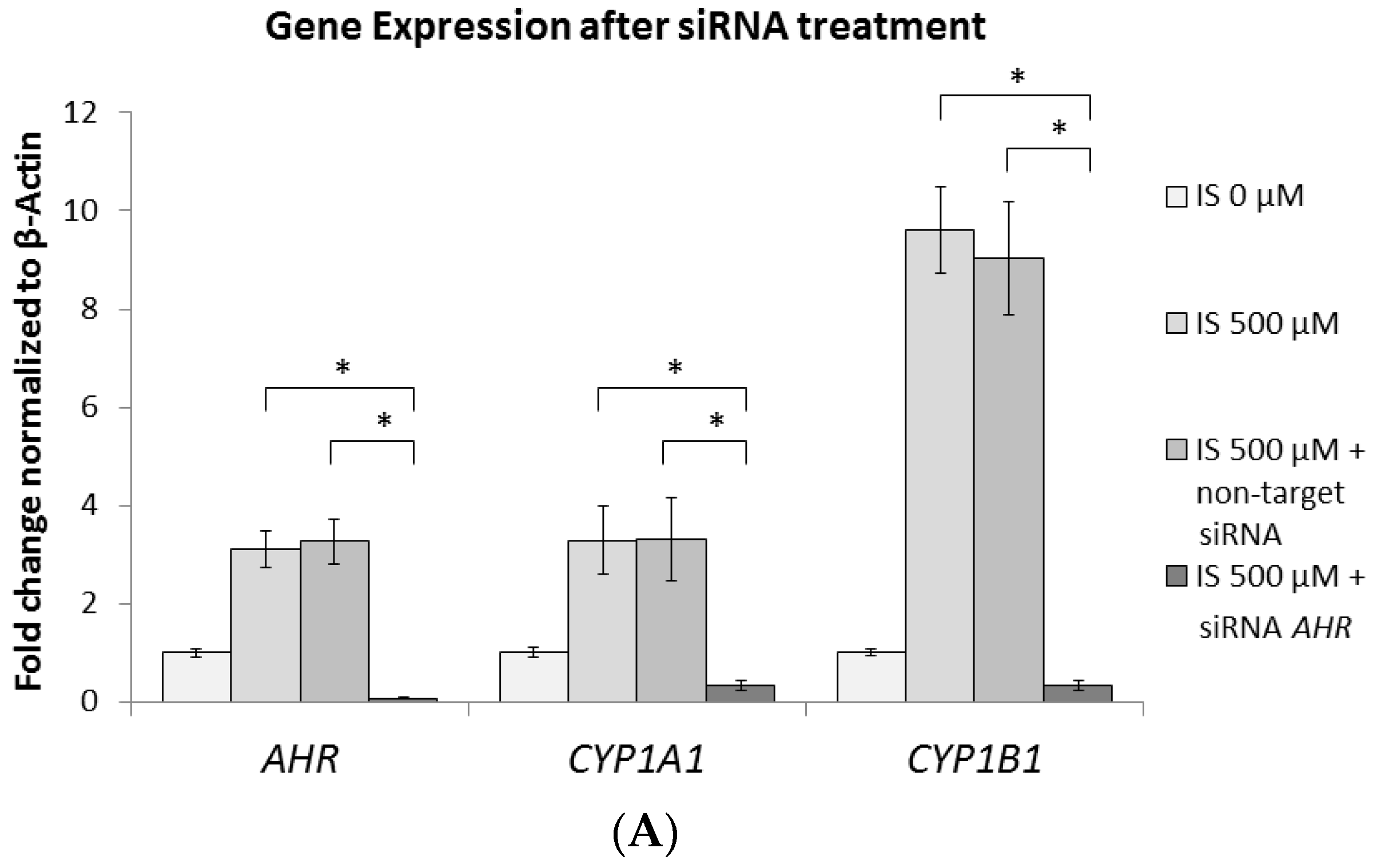
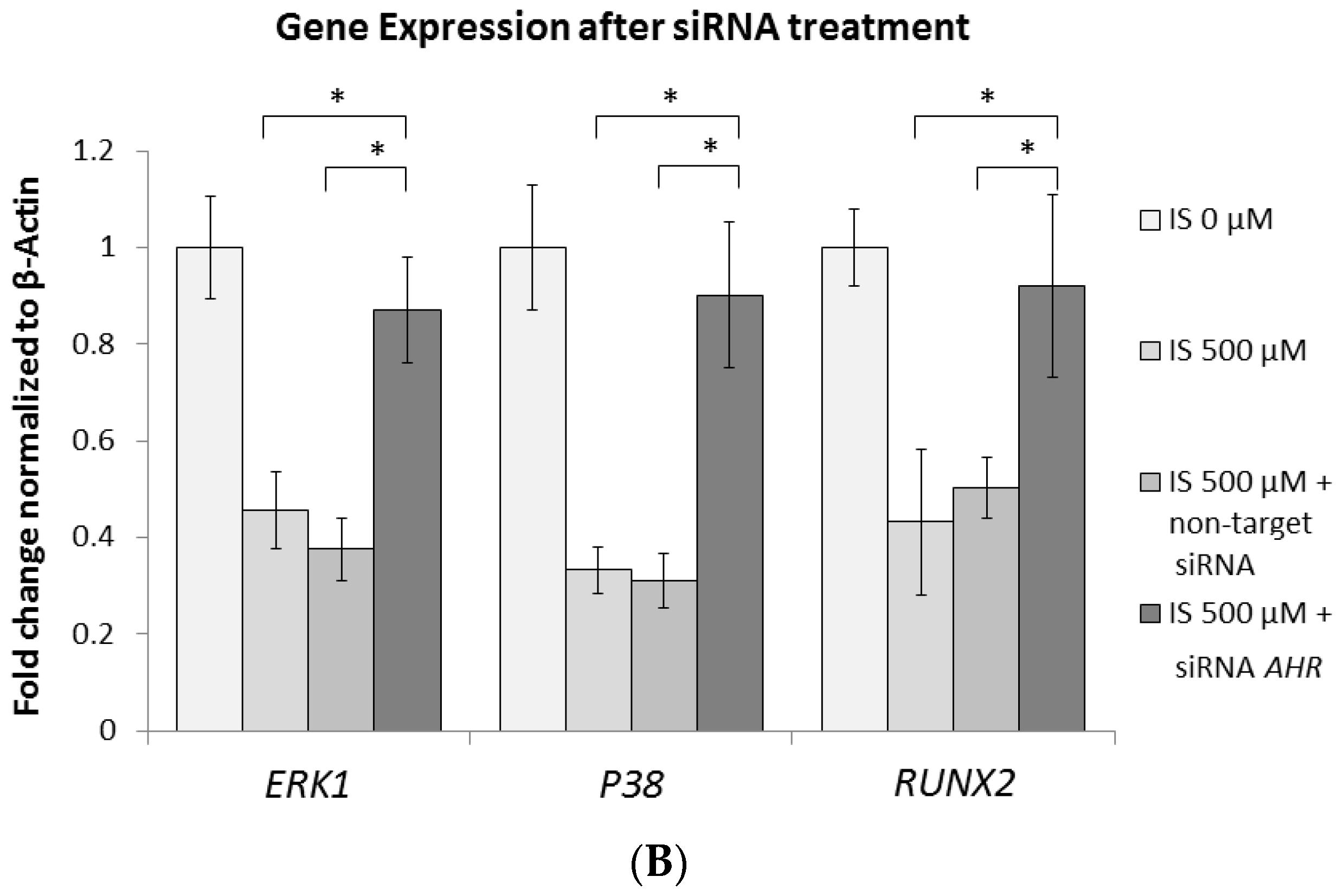
2.6. AhR Knockdown Rescues the Downstream Expression of ERK, p38 MAPK, and Runx2
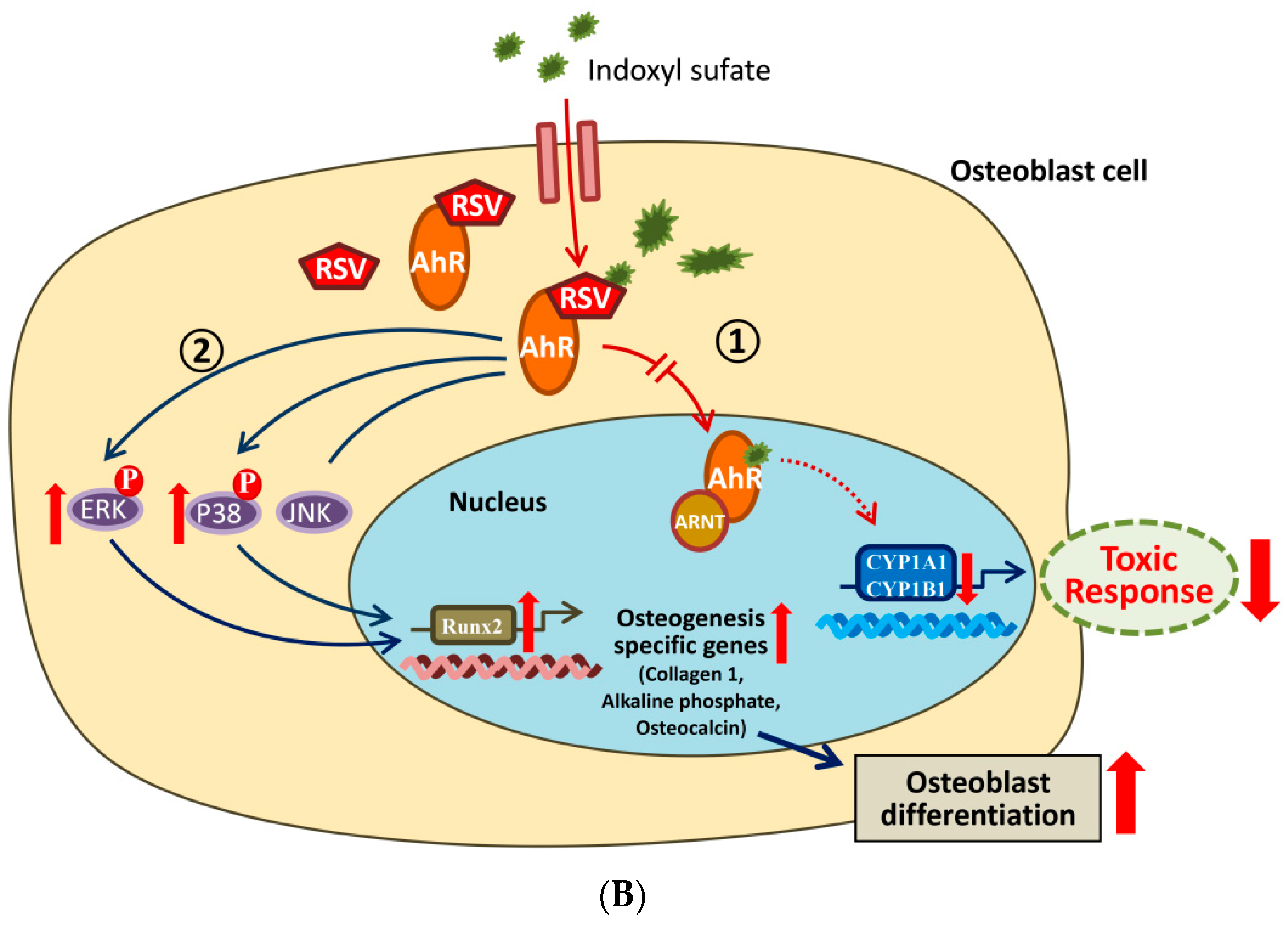
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. In Vivo Study
4.2.1. Serum Creatinine (Scr) and Blood Urea Nitrogen (BUN) Measurement
4.2.2. H&E and IHC Staining
4.2.3. Micro-CT Analysis
4.3. In Vitro Study
4.3.1. Primary Culture of Osteoblastic Cells
4.3.2. MC3T3-E1 Cell Line
4.3.3. Cell Viability
4.3.4. Alkaline Phosphatase Staining and Alkaline Phosphatase Activity
4.3.5. Alizarin Red Staining and Quantification Assay
4.3.6. Western Blot Analysis
4.3.7. qPCR and si-RNA AhR
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AhR | Aryl hydrocarbon Receptor |
| ARNT | Aromatic hydrocarbon Receptor Nuclear Transfer Protein |
| BMSCs | Bone Marrow-derived Stem Cells |
| BUN | Blood Urea Nitrogen |
| BV/TV | Bone Volume over Total Volume |
| CKD-MBD | Chronic Kidney Disease – Mineral and Bone Disorder |
| CYP1A1 | Cytochrome P450 family 1 Subfamily A Member 1 |
| CYP1B1 | Cytochrome P450 family 1 Subfamily B Member 1 |
| DAB | 3,3’-Diaminobenzidine |
| H&E staining | Hematoxylin and Eosin Staining |
| IHC staining | Immunohistochemical staining |
| IS | Indoxyl Sulfate |
| HRP | Horse Radish peroxidase |
| MAPK | Mitogen-activated Protein Kinase |
| N/S | Normal Saline |
| Nx | Nephrectomy |
| PTH | Parathyroid Hormone |
| RSV | Resveratrol |
| Runx2 | Runt-related Transcription Factor 2 |
| siRNA | Small Interfering RNA |
| Tb.N. | Trabecular Number |
| Tb.Sp. | Trabecular Separation |
| Tb.Th. | Trabecular Thickness |
| TCDD | 2,3,7,8-tetrachlorodibenzo-p-dioxin |
References
- Raff, A.C.; Meyer, T.W.; Hostetter, T.H. New insights into uremic toxicity. Curr. Opin. Nephrol. Hypertens. 2008, 17, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Adesso, S.; Popolo, A.; Bianco, G.; Sorrentino, R.; Pinto, A.; Autore, G.; Marzocco, S. The uremic toxin indoxyl sulphate enhances macrophage response to LPS. PLoS ONE 2013, 8, e76778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-C.; Tomino, Y.; Lu, K.-C. Impacts of indoxyl sulfate and p-cresol sulfate on chronic kidney disease and mitigating effects of AST-120. Toxins 2018, 10, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-C.; Shyu, J.-F.; Lim, P.S.; Fang, T.-C.; Lu, C.-L.; Zheng, C.-M.; Hou, Y.-C.; Wu, C.-C.; Lin, Y.-F.; Lu, K.-C. Concentration and duration of indoxyl sulfate exposure affects osteoclastogenesis by regulating NFATc1 via aryl hydrocarbon receptor. Int. J. Mol. Sci. 2020, 21, 3486. [Google Scholar] [CrossRef]
- Mozar, A.; Louvet, L.; Godin, C.; Mentaverri, R.; Brazier, M.; Kamel, S.; Massy, Z.A. Indoxyl sulphate inhibits osteoclast differentiation and function. Nephrol. Dial. Transplant. 2011, 27, 2176–2181. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-H.; Kwak, K.-A.; Gil, H.W.; Song, H.-Y.; Hong, S.Y. Indoxyl sulfate promotes apoptosis in cultured osteoblast cells. BMC Pharmacol. Toxicol. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Nii-Kono, T.; Iwasaki, Y.; Uchida, M.; Fujieda, A.; Hosokawa, A.; Motojima, M.; Yamato, H.; Kurokawa, K.; Fukagawa, M. Indoxyl sulfate induces skeletal resistance to parathyroid hormone in cultured osteoblastic cells. Kidney Int. 2007, 71, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Gehm, B.D.; McAndrews, J.M.; Chien, P.-Y.; Jameson, J.L. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc. Natl. Acad. Sci. 1997, 94, 14138–14143. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef] [Green Version]
- Most, J.; Penders, J.; Lucchesi, M.; Goossens, G.H.; E Blaak, E. Gut microbiota composition in relation to the metabolic response to 12-week combined polyphenol supplementation in overweight men and women. Eur. J. Clin. Nutr. 2017, 71, 1040–1045. [Google Scholar] [CrossRef]
- Chen, M.-L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.-D.; Zhang, Q.-Y.; Mi, M. Resveratrol attenuates trimethylamine-N-oxide (TMAO)-induced atherosclerosis by regulating TMAO synthesis and bile acid metabolism via remodeling of the gut Microbiota. mBio 2016, 7, e02210-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, H.; Yoshimura, M.; Saigo, C.; Komori, M.; Nomura, Y.; Yamamoto, Y.; Sagata, M.; Wakida, A.; Chuman, E.; Nishi, K.; et al. Hepatic sulfotransferase as a nephropreventing target by suppression of the uremic toxin indoxyl sulfate accumulation in ischemic acute kidney injury. Toxicol. Sci. 2014, 141, 206–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, R.K.; Jaiswal, N.; Bruder, S.P.; Mbalaviele, G.; Marshak, D.R.; Pittenger, M.F. Adult human mesenchymal stem cell differentiation to the osteogenic or adipogenic lineage is regulated by mitogen-activated protein kinase. J. Biol. Chem. 2000, 275, 9645–9652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.; Li, Y.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol enhances proliferation and osteoblastic differentiation in human mesenchymal stem cells via er-dependent erk1/2 activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Tou, L.; Quibria, N.; Alexander, J.M. Transcriptional regulation of the human runx2/cbfa1 gene promoter by bone morphogenetic protein-7. Mol. Cell. Endocrinol. 2003, 205, 121–129. [Google Scholar] [CrossRef]
- Ge, C.; Yang, Q.; Zhao, G.; Yu, H.; Kirkwood, K.L.; Franceschi, R.T. Interactions between extracellular signal-regulated kinase 1/2 and p38 map kinase pathways in the control of runx2 phosphorylation and transcriptional activity. J. Bone. Miner. Res. 2012, 27, 538–551. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.; Jiang, D.; Thomas, P.; Benson, M.D.; Guan, K.; Karsenty, G.; Franceschi, R.T. MAPK pathways activate and phosphorylate the osteoblast-specific transcription factor, Cbfa1. J. Biol. Chem. 2000, 275, 4453–4459. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, R.T.; Ge, C.; Xiao, G.; Roca, H.; Jiang, D. Transcriptional regulation of osteoblasts. Ann. N. Y. Acad. Sci. 2007, 1116, 196–207. [Google Scholar] [CrossRef]
- Stevens, E.A.; Mezrich, J.D.; Bradfield, C.A. The aryl hydrocarbon receptor: A perspective on potential roles in the immune system. Immunology 2009, 127, 299–311. [Google Scholar] [CrossRef]
- Murray, I.A.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat. Rev. Cancer 2014, 14, 801–814. [Google Scholar] [CrossRef]
- Schroeder, J.C.; DiNatale, B.C.; Murray, I.A.; Flaveny, C.A.; Liu, Q.; Laurenzana, E.M.; Lin, J.M.; Strom, S.C.; Omiecinski, C.J.; Amin, S.; et al. The uremic toxin 3-indoxyl sulfate is a potent endogenous agonist for the human aryl hydrocarbon receptor. Biochemistry 2010, 49, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Jiang, L.; Wan, B.; Zhang, W.; Yao, L.; Che, T.; Gan, C.; Su, N.; He, J.; Huang, J.; et al. The role of aryl hydrocarbon receptor in bone remodeling. Prog. Biophys. Mol. Biol. 2018, 134, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Korkalainen, M.; Kallio, E.; Olkku, A.; Nelo, K.; Ilvesaro, J.; Tuukkanen, J.; Mahonen, A.; Viluksela, M. Dioxins interfere with differentiation of osteoblasts and osteoclasts. Bone 2009, 44, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Ciolino, H.P.; Daschner, P.J.; Yeh, G.C. Resveratrol inhibits transcription of CYP1A1 in vitro by preventing activation of the aryl hydrocarbon receptor. Cancer Res. 1998, 58, 5707–5712. [Google Scholar]
- Tao, K.; Xiao, D.; Weng, J.; Xiong, A.; Kang, B.; Zeng, H. Berberine promotes bone marrow-derived mesenchymal stem cells osteogenic differentiation via canonical wnt/beta-catenin signaling pathway. Toxicol. Lett. 2016, 240, 68–80. [Google Scholar] [CrossRef]
- Qiu, J.; Li, J.; Mou, X.; Guo, W.; Wang, S.; Yu, X.; Ma, B.; Zhang, S.; Tang, W.; Sang, Y.; et al. Effects of graphene quantum dots on the self-renewal and differentiation of mesenchymal stem cells. Adv. Heal. Mater. 2016, 5, 702–710. [Google Scholar] [CrossRef]
- Gregory, C.A.; Gunn, W.G.; Peister, A.; Prockop, D.J. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef]
- Chmill, S.; Kadow, S.; Winter, M.; Weighardt, H.; Esser, C. 2,3,7,8-tetrachlorodibenzo-p-dioxin impairs stable establishment of oral tolerance in mice. Toxicol. Sci. 2010, 118, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Kazantseva, M.G.; Highton, J.; Stamp, L.K.; Hessian, P.A. Dendritic cells provide a potential link between smoking and inflammation in rheumatoid arthritis. Arthritis Res. Ther. 2012, 14, R208. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Sun, L.; Cao, J.; Yuen, T.; Lu, P.; Bab, I.; Leu, N.A.; Srinivasan, S.; Wagage, S.; Hunter, C.A.; et al. Smoke carcinogens cause bone loss through the aryl hydrocarbon receptor and induction of Cyp1 enzymes. Proc. Natl. Acad. Sci. 2013, 110, 11115–11120. [Google Scholar] [CrossRef] [Green Version]
- Ikuta, T.; Ohba, M.; Zouboulis, C.C.; Fujii-Kuriyama, Y.; Kawajiri, K. B lymphocyte-induced maturation protein 1 is a novel target gene of aryl hydrocarbon receptor. J. Dermatol. Sci. 2010, 58, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of proliferation, differentiation and functions of osteoblasts by Runx2. Int. J. Mol. Sci. 2019, 20, 1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akrami, R.; Skanderup, A.J.; Hoell, J.; Schultz, N.; Sander, C.; Larsson, E. Comprehensive analysis of long non-coding RNAs in ovarian cancer reveals global patterns and targeted DNA amplification. PLoS ONE 2013, 8, e80306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grafe, I.; Alexander, S.; Peterson, J.R.; Snider, T.N.; Levi, B.; Lee, B.; Mishina, Y. Tgf-beta family signaling in mesenchymal differentiation. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef]
- Rutkovskiy, A.; Stensløkken, K.-O.; Vaage, I.J. Osteoblast differentiation at a glance. Med Sci. Monit. Basic Res. 2016, 22, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Zheng, L.; Zhao, H.; Miao, J.; Sun, C.; Ren, N.; Wang, J.; Liu, H.; Tao, X. In vitro assessment of the differentiation potential of bone marrow-derived mesenchymal stem cells on genipin-chitosan conjugation scaffold with surface hydroxyapatite nanostructure for bone tissue engineering. Tissue Eng. Part A 2011, 17, 1341–1349. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Mou, X.; Qiu, J.; Wang, S.; Wang, D.; Sun, D.; Guo, W.; Li, D.; Kumar, A.; Yang, X.; et al. Surface charge regulation of osteogenic differentiation of mesenchymal stem cell on polarized ferroelectric crystal substrate. Adv. Heal. Mater. 2015, 4, 998–1003. [Google Scholar] [CrossRef]
- Ge, C.; Xiao, G.; Jiang, D.; Franceschi, R.T. Critical role of the extracellular signal-regulated kinase-MAPK pathway in osteoblast differentiation and skeletal development. J. Cell Biol. 2007, 176, 709–718. [Google Scholar] [CrossRef]
- Greenblatt, M.B.; Shim, J.-H.; Zou, W.; Sitara, D.; Schweitzer, M.; Hu, R.; Lotinun, S.; Sano, Y.; Baron, R.; Park, J.M.; et al. The p38 MAPK pathway is essential for skeletogenesis and bone homeostasis in mice. J. Clin. Investig. 2010, 120, 2457–2473. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Greenblatt, M.B.; Shim, J.-H.; Kant, S.; Zhai, B.; Lotinun, S.; Brady, N.J.; Hu, D.Z.; Gygi, S.P.; Baron, R.; et al. MLK3 regulates bone development downstream of the faciogenital dysplasia protein FGD1 in mice. J. Clin. Investig. 2011, 121, 4383–4392. [Google Scholar] [CrossRef]
- Olsen, B.R.; Reginato, A.M.; Wang, W. Bone development. Annu. Rev. Cell. Dev. Biol. 2000, 16, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Kazama, J.J.; Matsuo, K.; Iwasaki, Y.; Fukagawa, M. Chronic kidney disease and bone metabolism. J. Bone Miner. Metab. 2015, 33, 245–252. [Google Scholar] [CrossRef]
- Hirata, J.; Hirai, K.; Asai, H.; Matsumoto, C.; Inada, M.; Miyaura, C.; Yamato, H.; Watanabe-Akanuma, M. Indoxyl sulfate exacerbates low bone turnover induced by parathyroidectomy in young adult rats. Bone 2015, 79, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, H.M.; Pulkkinen, P.; Jämsä, T.; Koistinen, J.; Simanainen, U.; Tuomisto, J.; Tuukkanen, J.; Viluksela, M. Effects of in utero and lactational TCDD exposure on bone development in differentially sensitive rat lines. Toxicol. Sci. 2005, 85, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Hanieh, H.; Nakahama, T.; Kishimoto, T. The roles of aryl hydrocarbon receptor in immune responses. Int. Immunol. 2013, 25, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Jämsä, T.; Viluksela, M.; Tuomisto, J.T.; Tuomisto, J.; Tuukkanen, J. Effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin on bone in two rat strains with different aryl hydrocarbon receptor structures. J. Bone Miner. Res. 2001, 16, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.-Y.; Kondo, T.; Matsumoto, T.; Fujii-Kuriyama, Y.; Imai, Y. Aryl hydrocarbon receptor catabolic activity in bone metabolism is osteoclast dependent in vivo. Biochem. Biophys. Res. Commun. 2014, 450, 416–422. [Google Scholar] [CrossRef]
- Ryan, E.P.; Holz, J.D.; Mulcahey, M.; Sheu, T.-J.; A Gasiewicz, T.; Puzas, J.E. Environmental toxicants may modulate osteoblast differentiation by a mechanism involving the aryl hydrocarbon receptor. J. Bone Miner. Res. 2007, 22, 1571–1580. [Google Scholar] [CrossRef]
- Rodriguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK Signaling in osteoblast differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Henklová, P.; Vrzal, R.; Ulrichová, J.; Dvořák, Z. Role of mitogen-activated protein kinases in aryl hydrocarbon receptor signaling. Chem. Interactions 2008, 172, 93–104. [Google Scholar] [CrossRef]
- Puga, A.; Ma, C.; Marlowe, J.L. The aryl hydrocarbon receptor cross-talks with multiple signal transduction pathways. Biochem. Pharmacol. 2009, 77, 713–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, L.; Sallée, M.; Cerini, C.; Poitevin, S.; Gondouin, B.; Jourde-Chiche, N.; Fallague, K.; Brunet, P.; Calaf, R.; Dussol, B.; et al. The cardiovascular effect of the uremic solute indole-3 acetic acid. J. Am. Soc. Nephrol. 2014, 26, 876–887. [Google Scholar] [CrossRef] [PubMed]
- Addi, T.; Poitevin, S.; McKay, N.; El Mecherfi, K.E.; Kheroua, O.; Jourde-Chiche, N.; De Macedo, A.; Gondouin, B.; Cerini, C.; Brunet, P.; et al. Mechanisms of tissue factor induction by the uremic toxin indole-3 acetic acid through aryl hydrocarbon receptor/nuclear factor-kappa B signaling pathway in human endothelial cells. Arch. Toxicol. 2018, 93, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Du, Y.; Zhang, X.; Sun, Y.; Li, S.; Dou, Y.; Li, Z.; Yuan, H.; Zhao, W. The aryl hydrocarbon receptor suppresses osteoblast proliferation and differentiation through the activation of the ERK signaling pathway. Toxicol. Appl. Pharmacol. 2014, 280, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Assefa, E.G.; Yan, Q.; Gezahegn, S.B.; Salissou, M.T.M.; He, S.; Wu, N.; Zuo, X.; Ying, C. Role of resveratrol on indoxyl sulfate-induced endothelial hyperpermeability via aryl hydrocarbon receptor (ahr)/src-dependent pathway. Oxid. Med. Cell. Longev. 2019, 2019, 5847040. [Google Scholar] [CrossRef]
- Tan, Z.; Chang, X.; Puga, A.; Xia, Y. Activation of mitogen-activated protein kinases (MAPKs) by aromatic hydrocarbons: Role in the regulation of aryl hydrocarbon receptor (AHR) function. Biochem. Pharmacol. 2002, 64, 771–780. [Google Scholar] [CrossRef]
- Borsani, E.; Bonazza, V.; Buffoli, B.; Nocini, P.F.; Albanese, M.; Zotti, F.; Inchingolo, F.; Rezzani, R.; Rodella, L.F. Beneficial effects of concentrated growth factors and resveratrol on human osteoblasts in vitro treated with bisphosphonates. BioMed Res. Int. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gallo-Oller, G.; Ordoñez, R.; Dotor, J.; Ordoñez-Ciriza, R. A new background subtraction method for Western blot densitometry band quantification through image analysis software. J. Immunol. Methods 2018, 457, 1–5. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | Forward Primer 5′→3′ | Reverse Primer 5′→3′ |
|---|---|---|
| AHR | TTCTTAGGCTCAGCGTCAGCTA | GCAAATCCTGCCAGTCTCTGAT′ |
| Alkaline Phosphatase | GAGCGTCATCCCAGTGGAG | TAGCGGTTACTGTAGACACCC |
| Collagen 1 | GAGCGGAGAGTACTGGATCG | GCTTCTTTTCCTTGGGGTTC |
| CYP1A1 | GTCAGGACAGGAAGCTGGAC | GAGGCTCCACGAGATAGCAG |
| CYP1B1 | GCCACTATTACGGACATCTTCGG | ACAACCTGGTCCAACTCAGCCT |
| ERK1 | GACACCCCTGTCCTTTTGGATCTGGTCCTG | AAGCAGAGACCCCAGCAAAGTGAGAGAAG |
| Osteopontin | CCGCTCGAGACCATGAGATTGGCAGTGAT | CGGGATCCCTAGTTGACCTCAGAAGATG |
| p38 | GAAAGTGTAAAGCCAATTCCAGTGTTGGAC | GTCTGCCTCCTTCTGGGCTCCAAATGATTC |
| RUNX2 | TTCAACGATCTGAGATTTGTGGG | GGATGAGGAATGCGCCCTA |
| β-Actin | CCTCTATGCCAACACGTGC | CCTGCTTGCTGATCCACATC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-C.; Shyu, J.-F.; Lin, Y.-F.; Chiu, H.-W.; Lim, P.S.; Lu, C.-L.; Zheng, C.-M.; Hou, Y.-C.; Chen, P.-H.; Lu, K.-C. Resveratrol Rescue Indoxyl Sulfate-Induced Deterioration of Osteoblastogenesis via the Aryl Hydrocarbon Receptor /MAPK Pathway. Int. J. Mol. Sci. 2020, 21, 7483. https://doi.org/10.3390/ijms21207483
Liu W-C, Shyu J-F, Lin Y-F, Chiu H-W, Lim PS, Lu C-L, Zheng C-M, Hou Y-C, Chen P-H, Lu K-C. Resveratrol Rescue Indoxyl Sulfate-Induced Deterioration of Osteoblastogenesis via the Aryl Hydrocarbon Receptor /MAPK Pathway. International Journal of Molecular Sciences. 2020; 21(20):7483. https://doi.org/10.3390/ijms21207483
Chicago/Turabian StyleLiu, Wen-Chih, Jia-Fwu Shyu, Yuh-Feng Lin, Hui-Wen Chiu, Paik Seong Lim, Chien-Lin Lu, Cai-Mei Zheng, Yi-Chou Hou, Po-Han Chen, and Kuo-Cheng Lu. 2020. "Resveratrol Rescue Indoxyl Sulfate-Induced Deterioration of Osteoblastogenesis via the Aryl Hydrocarbon Receptor /MAPK Pathway" International Journal of Molecular Sciences 21, no. 20: 7483. https://doi.org/10.3390/ijms21207483
APA StyleLiu, W. -C., Shyu, J. -F., Lin, Y. -F., Chiu, H. -W., Lim, P. S., Lu, C. -L., Zheng, C. -M., Hou, Y. -C., Chen, P. -H., & Lu, K. -C. (2020). Resveratrol Rescue Indoxyl Sulfate-Induced Deterioration of Osteoblastogenesis via the Aryl Hydrocarbon Receptor /MAPK Pathway. International Journal of Molecular Sciences, 21(20), 7483. https://doi.org/10.3390/ijms21207483