p38β and Cancer: The Beginning of the Road

 , , ,
, , , 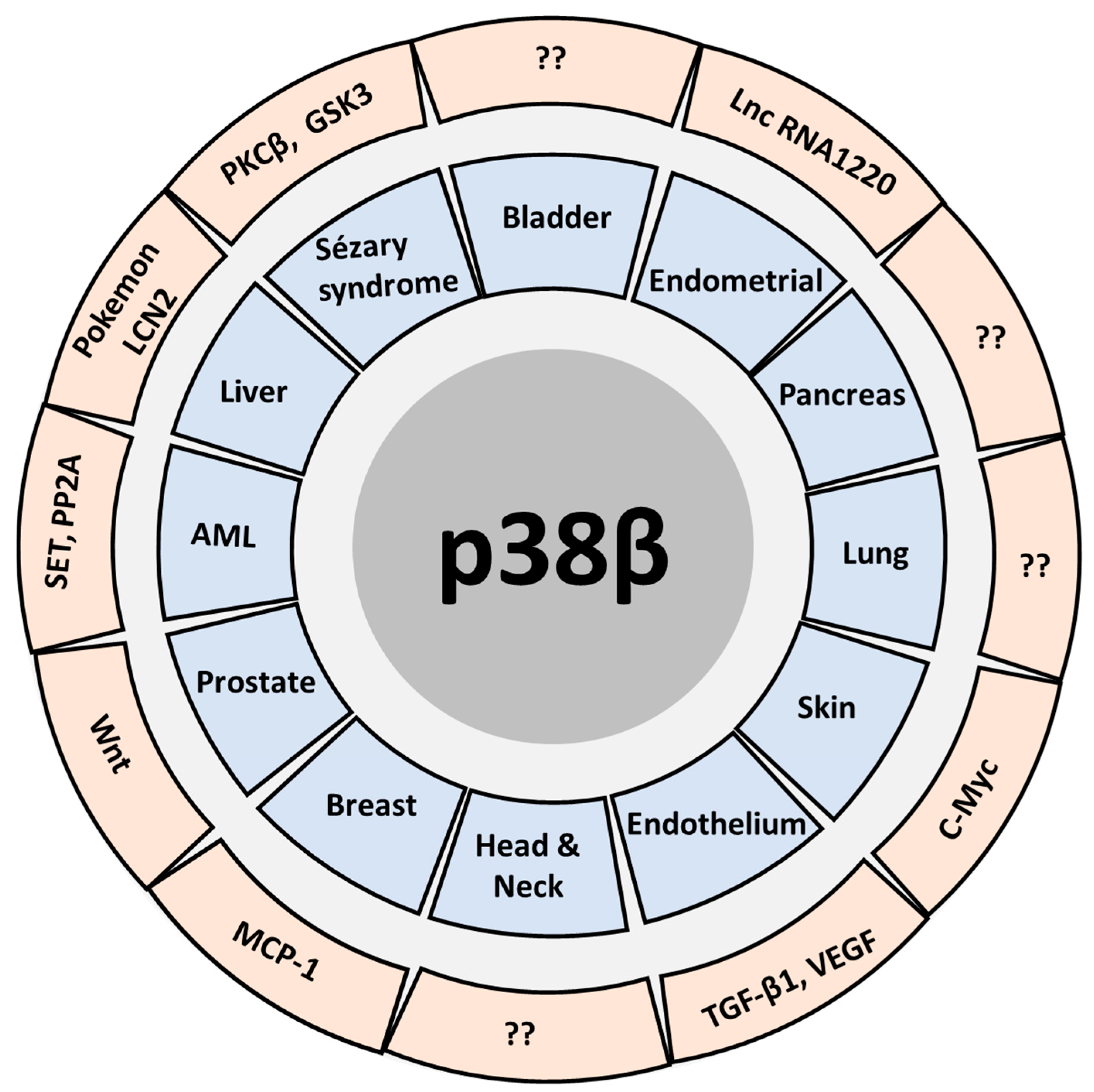{kind=link}
Abstract
:1. Introduction
2. The p38MAPK Family
3. p38β
4. p38β and Cancer
5. Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, E.K.; Choi, E.-J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Morrison, D.K.; Davis, R.J. Regulation of MAP kinase signaling modules by scaffold proteins in mammals. Annu. Rev. Cell Dev. Biol. 2003, 19, 91–118. [Google Scholar] [CrossRef] [PubMed]
- Peti, W.; Page, R. Molecular basis of MAP kinase regulation. Protein Sci. 2013, 22, 1698–1710. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulombe, P.; Meloche, S. Atypical mitogen-activated protein kinases: Structure, regulation and functions. Biochim. Biophys. Acta 2007, 1773, 1376–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, W.-L.; Duke, C.J.; Abe, M.K.; Kaplan, E.L.; Gomes, S.; Rosner, M.R. ERK7 expression and kinase activity is regulated by the ubiquitin-proteosome pathway. J. Biol. Chem. 2004, 279, 23073–23081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishitani, T.; Ishitani, S. Nemo-like kinase, a multifaceted cell signaling regulator. Cell. Signal. 2013, 25, 190–197. [Google Scholar] [CrossRef]
- Abe, M.K.; Saelzler, M.P.; Espinosa, R.; Kahle, K.T.; Hershenson, M.B.; Le Beau, M.M.; Rosner, M.R. ERK8, a new member of the mitogen-activated protein kinase family. J. Biol. Chem. 2002, 277, 16733–16743. [Google Scholar] [CrossRef] [Green Version]
- Yokota, T.; Wang, Y. p38 MAP kinases in the heart. Gene 2016, 575, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Wu, J.; Silke, J. An overview of mammalian p38 mitogen-activated protein kinases, central regulators of cell stress and receptor signaling. F1000Res 2020, 9, 653. [Google Scholar] [CrossRef]
- Remy, G.; Risco, A.M.; Iñesta-Vaquera, F.A.; González-Terán, B.; Sabio, G.; Davis, R.J.; Cuenda, A. Differential activation of p38MAPK isoforms by MKK6 and MKK3. Cell. Signal. 2010, 22, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Dérijard, B.; Raingeaud, J.; Barrett, T.; Wu, I.H.; Han, J.; Ulevitch, R.J.; Davis, R.J. Independent human MAP-kinase signal transduction pathways defined by MEK and MKK isoforms. Science 1995, 267, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Limón, A.; Joaquin, M.; Caballero, M.; Posas, F.; de Nadal, E. The p38 Pathway: From Biology to Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador, J.M.; Mittelstadt, P.R.; Guszczynski, T.; Copeland, T.D.; Yamaguchi, H.; Appella, E.; Fornace, A.J.; Ashwell, J.D. Alternative p38 activation pathway mediated by T cell receptor-proximal tyrosine kinases. Nat. Immunol. 2005, 6, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Peregrin, S.; Jurado-Pueyo, M.; Campos, P.M.; Sanz-Moreno, V.; Ruiz-Gomez, A.; Crespo, P.; Mayor, F.; Murga, C. Phosphorylation of p38 by GRK2 at the docking groove unveils a novel mechanism for inactivating p38MAPK. Curr. Biol. 2006, 16, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; McDonnell, P.C.; Gum, R.J.; Hand, A.T.; Lee, J.C.; Young, P.R. Novel homologues of CSBP/p38 MAP kinase: Activation, substrate specificity and sensitivity to inhibition by pyridinyl imidazoles. Biochem. Biophys. Res. Commun. 1997, 235, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Young, P.R.; McLaughlin, M.M.; Kumar, S.; Kassis, S.; Doyle, M.L.; McNulty, D.; Gallagher, T.F.; Fisher, S.; McDonnell, P.C.; Carr, S.A.; et al. Pyridinyl imidazole inhibitors of p38 mitogen-activated protein kinase bind in the ATP site. J. Biol. Chem. 1997, 272, 12116–12121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, V.; Nahidino, P.; Forster, M.; Laufer, S.A. An updated patent review of p38 MAP kinase inhibitors (2014–2019). Expert Opin. Ther. Pat. 2020, 30, 453–466. [Google Scholar] [CrossRef]
- Trempolec, N.; Dave-Coll, N.; Nebreda, A.R. SnapShot: p38 MAPK substrates. Cell 2013, 152, 924–924.e1. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Chen, C.; Li, Z.; Guo, W.; Gegner, J.A.; Lin, S.; Han, J. Characterization of the structure and function of a new mitogen-activated protein kinase (p38beta). J. Biol. Chem. 1996, 271, 17920–17926. [Google Scholar] [CrossRef] [Green Version]
- Stein, B.; Yang, M.X.; Young, D.B.; Janknecht, R.; Hunter, T.; Murray, B.W.; Barbosa, M.S. p38-2, a novel mitogen-activated protein kinase with distinct properties. J. Biol. Chem. 1997, 272, 19509–19517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.B.; Cameron, P.M.; O’Keefe, S.J.; Frantz-Wattley, B.; Thompson, J.; O’Neill, E.A.; Tennis, T.; Liu, L.; Becker, J.W.; Scapin, G. The three-dimensional structure of MAP kinase p38beta: Different features of the ATP-binding site in p38beta compared with p38alpha. Acta Crystallogr. D Biol. Crystallogr. 2009, 65, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Hale, K.K.; Trollinger, D.; Rihanek, M.; Manthey, C.L. Differential expression and activation of p38 mitogen-activated protein kinase alpha, beta, gamma, and delta in inflammatory cell lineages. J. Immunol. 1999, 162, 4246–4252. [Google Scholar] [PubMed]
- Johnston, S.J.; Ahmad, D.; Aleskandarany, M.A.; Kurozumi, S.; Nolan, C.C.; Diez-Rodriguez, M.; Green, A.R.; Rakha, E.A. Co-expression of nuclear P38 and hormone receptors is prognostic of good long-term clinical outcome in primary breast cancer and is linked to upregulation of DNA repair. BMC Cancer 2018, 18, 1027. [Google Scholar] [CrossRef] [Green Version]
- Enslen, H.; Raingeaud, J.; Davis, R.J. Selective activation of p38 mitogen-activated protein (MAP) kinase isoforms by the MAP kinase kinases MKK3 and MKK6. J. Biol. Chem. 1998, 273, 1741–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keesler, G.A.; Bray, J.; Hunt, J.; Johnson, D.A.; Gleason, T.; Yao, Z.; Wang, S.W.; Parker, C.; Yamane, H.; Cole, C.; et al. Purification and activation of recombinant p38 isoforms alpha, beta, gamma, and delta. Protein Expr. Purif. 1998, 14, 221–228. [Google Scholar] [CrossRef]
- Beenstock, J.; Ben-Yehuda, S.; Melamed, D.; Admon, A.; Livnah, O.; Ahn, N.G.; Engelberg, D. The p38β Mitogen-activated Protein Kinase Possesses an Intrinsic Autophosphorylation Activity, Generated by a Short Region Composed of the α-G Helix and MAPK Insert. J. Biol. Chem. 2014, 289, 23546–23556. [Google Scholar] [CrossRef] [Green Version]
- Beenstock, J.; Melamed, D.; Mooshayef, N.; Mordechay, D.; Garfinkel, B.P.; Ahn, N.G.; Admon, A.; Engelberg, D. p38β Mitogen-Activated Protein Kinase Modulates Its Own Basal Activity by Autophosphorylation of the Activating Residue Thr180 and the Inhibitory Residues Thr241 and Ser261. Mol. Cell. Biol. 2016, 36, 1540–1554. [Google Scholar] [CrossRef] [Green Version]
- del Barco Barrantes, I.; Coya, J.M.; Maina, F.; Arthur, J.S.C.; Nebreda, A.R. Genetic analysis of specific and redundant roles for p38alpha and p38beta MAPKs during mouse development. Proc. Natl. Acad. Sci. USA 2011, 108, 12764–12769. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.H.; Porras, A.; Alonso, G.; Jones, M.; Vintersten, K.; Panelli, S.; Valladares, A.; Perez, L.; Klein, R.; Nebreda, A.R. Essential role of p38alpha MAP kinase in placental but not embryonic cardiovascular development. Mol. Cell 2000, 6, 109–116. [Google Scholar] [CrossRef]
- Beardmore, V.A.; Hinton, H.J.; Eftychi, C.; Apostolaki, M.; Armaka, M.; Darragh, J.; McIlrath, J.; Carr, J.M.; Armit, L.J.; Clacher, C.; et al. Generation and characterization of p38beta (MAPK11) gene-targeted mice. Mol. Cell. Biol. 2005, 25, 10454–10464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaestel, M. MAPKAP kinases-MKs-two’s company, three’s a crowd. Nat. Rev. Mol. Cell Biol. 2006, 7, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Llopis, A.; Salvador, N.; Ercilla, A.; Guaita-Esteruelas, S.; Barrantes, I.D.B.; Gupta, J.; Gaestel, M.; Davis, R.J.; Nebreda, A.R.; Agell, N. The stress-activated protein kinases p38α/β and JNK1/2 cooperate with Chk1 to inhibit mitotic entry upon DNA replication arrest. Cell Cycle 2012, 11, 3627–3637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifton, A.D.; Young, P.R.; Cohen, P. A comparison of the substrate specificity of MAPKAP kinase-2 and MAPKAP kinase-3 and their activation by cytokines and cellular stress. FEBS Lett. 1996, 392, 209–214. [Google Scholar] [CrossRef] [Green Version]
- New, L.; Jiang, Y.; Zhao, M.; Liu, K.; Zhu, W.; Flood, L.J.; Kato, Y.; Parry, G.C.; Han, J. PRAK, a novel protein kinase regulated by the p38 MAP kinase. EMBO J. 1998, 17, 3372–3384. [Google Scholar] [CrossRef] [Green Version]
- New, L.; Jiang, Y.; Han, J. Regulation of PRAK subcellular location by p38 MAP kinases. Mol. Biol. Cell 2003, 14, 2603–2616. [Google Scholar] [CrossRef] [Green Version]
- Saurin, A.T.; Durgan, J.; Cameron, A.J.; Faisal, A.; Marber, M.S.; Parker, P.J. The regulated assembly of a PKCepsilon complex controls the completion of cytokinesis. Nat. Cell Biol. 2008, 10, 891–901. [Google Scholar] [CrossRef]
- Pon, J.R.; Marra, M.A. MEF2 transcription factors: Developmental regulators and emerging cancer genes. Oncotarget 2015, 7, 2297–2312. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-H.; Galanis, A.; Sharrocks, A.D. Targeting of p38 Mitogen-Activated Protein Kinases to MEF2 Transcription Factors. Mol. Cell. Biol. 1999, 19, 4028–4038. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Jiang, Y.; Li, Z.; Kravchenko, V.V.; Ulevitch, R.J. Activation of the transcription factor MEF2C by the MAP kinase p38 in inflammation. Nature 1997, 386, 296–299. [Google Scholar] [CrossRef]
- Ho, I.C.; Kim, J.H.; Rooney, J.W.; Spiegelman, B.M.; Glimcher, L.H. A potential role for the nuclear factor of activated T cells family of transcriptional regulatory proteins in adipogenesis. Proc. Natl. Acad. Sci. USA 1998, 95, 15537–15541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushdid, P.B.; Osinska, H.; Waclaw, R.R.; Molkentin, J.D.; Yutzey, K.E. NFATc3 and NFATc4 are required for cardiac development and mitochondrial function. Circ. Res. 2003, 92, 1305–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.T.C.; Xiong, Q.; Enslen, H.; Davis, R.J.; Chow, C.-W. Phosphorylation of NFATc4 by p38 mitogen-activated protein kinases. Mol. Cell. Biol. 2002, 22, 3892–3904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.T.C.; Yu, R.Y.L.; Agadir, A.; Gao, G.-J.; Campos-Gonzalez, R.; Tournier, C.; Chow, C.-W. Integration of Protein Kinases mTOR and Extracellular Signal-Regulated Kinase 5 in Regulating Nucleocytoplasmic Localization of NFATc4. Mol. Cell. Biol. 2008, 28, 3489–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaulian, E.; Karin, M. AP-1 as a regulator of cell life and death. Nat. Cell Biol. 2002, 4, E131–E136. [Google Scholar] [CrossRef]
- Tanos, T.; Marinissen, M.J.; Leskow, F.C.; Hochbaum, D.; Martinetto, H.; Gutkind, J.S.; Coso, O.A. Phosphorylation of c-Fos by members of the p38 MAPK family. Role in the AP-1 response to UV light. J. Biol. Chem. 2005, 280, 18842–18852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenda, A.; Cohen, P.; Buée-Scherrer, V.; Goedert, M. Activation of stress-activated protein kinase-3 (SAPK3) by cytokines and cellular stresses is mediated via SAPKK3 (MKK6); comparison of the specificities of SAPK3 and SAPK2 (RK/p38). EMBO J. 1997, 16, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Mahlknecht, U.; Will, J.; Varin, A.; Hoelzer, D.; Herbein, G. Histone deacetylase 3, a class I histone deacetylase, suppresses MAPK11-mediated activating transcription factor-2 activation and represses TNF gene expression. J. Immunol. 2004, 173, 3979–3990. [Google Scholar] [CrossRef] [Green Version]
- Kaneto, H.; Matsuoka, T. Role of pancreatic transcription factors in maintenance of mature β-cell function. Int. J. Mol. Sci. 2015, 16, 6281–6297. [Google Scholar] [CrossRef] [Green Version]
- Sii-Felice, K.; Pouponnot, C.; Gillet, S.; Lecoin, L.; Girault, J.-A.; Eychène, A.; Felder-Schmittbuhl, M.-P. MafA transcription factor is phosphorylated by p38 MAP kinase. FEBS Lett. 2005, 579, 3547–3554. [Google Scholar] [CrossRef] [Green Version]
- Simone, C.; Forcales, S.V.; Hill, D.A.; Imbalzano, A.N.; Latella, L.; Puri, P.L. p38 pathway targets SWI-SNF chromatin-remodeling complex to muscle-specific loci. Nat. Genet. 2004, 36, 738–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forcales, S.V.; Albini, S.; Giordani, L.; Malecova, B.; Cignolo, L.; Chernov, A.; Coutinho, P.; Saccone, V.; Consalvi, S.; Williams, R.; et al. Signal-dependent incorporation of MyoD–BAF60c into Brg1-based SWI/SNF chromatin-remodelling complex. EMBO J. 2012, 31, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Page, J.L.; Wang, X.; Sordillo, L.M.; Johnson, S.E. MEKK1 signaling through p38 leads to transcriptional inactivation of E47 and repression of skeletal myogenesis. J. Biol. Chem. 2004, 279, 30966–30972. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado, A.; Lafarga, V.; Cheung, P.C.F.; Dolado, I.; Llanos, S.; Cohen, P.; Nebreda, A.R. A new p38 MAP kinase-regulated transcriptional coactivator that stimulates p53-dependent apoptosis. EMBO J. 2007, 26, 2115–2126. [Google Scholar] [CrossRef]
- Cuadrado, A.; Corrado, N.; Perdiguero, E.; Lafarga, V.; Muñoz-Canoves, P.; Nebreda, A.R. Essential role of p18Hamlet/SRCAP-mediated histone H2A.Z chromatin incorporation in muscle differentiation. EMBO J. 2010, 29, 2014–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanovas, O.; Jaumot, M.; Paules, A.-B.; Agell, N.; Bachs, O. P38SAPK2 phosphorylates cyclin D3 at Thr-283 and targets it for proteasomal degradation. Oncogene 2004, 23, 7537–7544. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Shi, Y.; Wang, Z.; Song, Z.; Zhu, M.; Cai, Q.; Chen, T. Serum starvation induces H2AX phosphorylation to regulate apoptosis via p38 MAPK pathway. FEBS Lett. 2008, 582, 2703–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briata, P.; Forcales, S.V.; Ponassi, M.; Corte, G.; Chen, C.-Y.; Karin, M.; Puri, P.L.; Gherzi, R. p38-dependent phosphorylation of the mRNA decay-promoting factor KSRP controls the stability of select myogenic transcripts. Mol. Cell 2005, 20, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Derynck, R. Direct activation of TACE-mediated ectodomain shedding by p38 MAP kinase regulates EGF receptor-dependent cell proliferation. Mol. Cell 2010, 37, 551–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuma, Y.; Campbell, D.G.; Cuenda, A. Identification of glycogen synthase as a new substrate for stress-activated protein kinase 2b/p38beta. Biochem. J. 2004, 379, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.-N.; Wang, X.-K.; Wu, S.-Q.; Lu, J.; Zheng, M.; Wang, Y.-H.; Zhou, H.; Zhang, H.; Han, J. Phosphorylation of Raptor by p38beta participates in arsenite-induced mammalian target of rapamycin complex 1 (mTORC1) activation. J. Biol. Chem. 2011, 286, 31501–31511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Fu, Y.; Liang, Y.; Song, H.; Yao, Y.; Wu, P.; Yao, Y.; Pan, Y.; Wen, X.; Ma, L.; et al. Suppression of MAPK11 or HIPK3 reduces mutant Huntingtin levels in Huntington’s disease models. Cell Res. 2017, 27, 1441–1465. [Google Scholar] [CrossRef]
- Zhang, S.; Weinheimer, C.; Courtois, M.; Kovacs, A.; Zhang, C.E.; Cheng, A.M.; Wang, Y.; Muslin, A.J. The role of the Grb2-p38 MAPK signaling pathway in cardiac hypertrophy and fibrosis. J. Clin. Investig. 2003, 111, 833–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, B.; Chaube, U.; Patel, B.M. Beneficial Effect of Silymarin in Pressure Overload Induced Experimental Cardiac Hypertrophy. Cardiovasc. Toxicol. 2019, 19, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Raingeaud, J.; Kaneki, M.; Weichselbaum, R.; Davis, R.J.; Kufe, D.; Kharbanda, S. Activation of p38 mitogen-activated protein kinase by c-Abl-dependent and -independent mechanisms. J. Biol. Chem. 1996, 271, 23775–23779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, J.; del Barco Barrantes, I.; Igea, A.; Sakellariou, S.; Pateras, I.S.; Gorgoulis, V.G.; Nebreda, A.R. Dual function of p38α MAPK in colon cancer: Suppression of colitis-associated tumor initiation but requirement for cancer cell survival. Cancer Cell 2014, 25, 484–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitos-Faleato, J.; Real, S.M.; Gutierrez-Prat, N.; Villanueva, A.; Llonch, E.; Drosten, M.; Barbacid, M.; Nebreda, A.R. Requirement for epithelial p38α in KRAS-driven lung tumor progression. Proc. Natl. Acad. Sci. USA 2020, 117, 2588–2596. [Google Scholar] [CrossRef]
- Sahu, V.; Mohan, A.; Dey, S. p38 MAP kinases: Plausible diagnostic and prognostic serum protein marker of non small cell lung cancer. Exp. Mol. Pathol. 2019, 107, 118–123. [Google Scholar] [CrossRef]
- Roseweir, A.K.; Halcrow, E.S.; Chichilo, S.; Powell, A.G.; McMillan, D.C.; Horgan, P.G.; Edwards, J. ERK and p38MAPK combine to improve survival in patients with BRAF mutant colorectal cancer. Br. J. Cancer 2018, 119, 323–329. [Google Scholar] [CrossRef]
- Igea, A.; Nebreda, A.R. The Stress Kinase p38α as a Target for Cancer Therapy. Cancer Res. 2015, 75, 3997–4002. [Google Scholar] [CrossRef] [Green Version]
- Del Reino, P.; Alsina-Beauchamp, D.; Escós, A.; Cerezo-Guisado, M.I.; Risco, A.; Aparicio, N.; Zur, R.; Fernandez-Estévez, M.; Collantes, E.; Montans, J.; et al. Pro-oncogenic role of alternative p38 mitogen-activated protein kinases p38γ and p38δ, linking inflammation and cancer in colitis-associated colon cancer. Cancer Res. 2014, 74, 6150–6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zur, R.; Garcia-Ibanez, L.; Nunez-Buiza, A.; Aparicio, N.; Liappas, G.; Escós, A.; Risco, A.; Page, A.; Saiz-Ladera, C.; Alsina-Beauchamp, D.; et al. Combined deletion of p38γ and p38δ reduces skin inflammation and protects from carcinogenesis. Oncotarget 2015, 6, 12920–12935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenda, A.; Sanz-Ezquerro, J.J. p38γ and p38δ: From Spectators to Key Physiological Players. Trends Biochem. Sci. 2017, 42, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, F.; Zhang, N.; Cao, D.; Liu, M.; Tan, Y.; Jiang, Y. p38β, A novel regulatory target of Pokemon in hepatic cells. Int. J. Mol. Sci. 2013, 14, 13511–13524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Sánchez, G.S.; Pita-Grisanti, V.; Quiñones-Díaz, B.; Gumpper, K.; Cruz-Monserrate, Z.; Vivas-Mejía, P.E. Biological Functions and Therapeutic Potential of Lipocalin 2 in Cancer. Int. J. Mol. Sci. 2020, 21, 4365. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Chaudhary, N.; Shah, S.; Braggs, C.; Sawant, A.; Vaz, S.; Thorat, R.; Gupta, S.; Dalal, S.N. Plakophilin3 loss leads to an increase in lipocalin2 expression, which is required for tumour formation. Exp. Cell Res. 2018, 369, 251–265. [Google Scholar] [CrossRef]
- Duperret, E.K.; Dahal, A.; Ridky, T.W. Focal-adhesion-independent integrin-αv regulation of FAK and c-Myc is necessary for 3D skin formation and tumor invasion. J. Cell Sci. 2015, 128, 3997–4013. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, G.; Terushkin, V.; Wolff, M.J.; Zhang, X.; Valacca, C.; Poggio, P.; Pintucci, G.; Mignatti, P. TGF-β1 induces endothelial cell apoptosis by shifting VEGF activation of p38(MAPK) from the prosurvival p38β to proapoptotic p38α. Mol. Cancer Res. 2012, 10, 605–614. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Pedram, A.; Razandi, M.; Levin, E.R. Estrogen prevents cardiomyocyte apoptosis through inhibition of reactive oxygen species and differential regulation of p38 kinase isoforms. J. Biol. Chem. 2006, 281, 6760–6767. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Janjanam, J.; Singh, N.K.; Rao, G.N. A new role for cofilin in retinal neovascularization. J. Cell. Sci. 2016, 129, 1234–1249. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Sin, K.W.T.; Ding, H.; Doan, H.A.; Gao, S.; Miao, H.; Wei, Y.; Wang, Y.; Zhang, G.; Li, Y.-P. p38β MAPK mediates ULK1-dependent induction of autophagy in skeletal muscle of tumor-bearing mice. Cell Stress 2018, 2, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Yao, J.; Zhuang, L.; Wang, D.; Han, J.; Lam, E.W.-F.; TCGA Research Network; Gan, B. The FoxO-BNIP3 axis exerts a unique regulation of mTORC1 and cell survival under energy stress. Oncogene 2014, 33, 3183–3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Xiang, H.-B.; Ye, D.-W.; Tian, X.-B. Inhibitory effects of intrathecal p38β antisense oligonucleotide on bone cancer pain in rats. Int. J. Clin. Exp. Pathol. 2014, 7, 7690–7698. [Google Scholar] [PubMed]
- Gao, J.; Chen, S.; Lin, S.; Han, H. Effect of music therapy on pain behaviors in rats with bone cancer pain. J. BUON 2016, 21, 466–472. [Google Scholar] [PubMed]
- Singh, A.K.; Pandey, R.; Gill, K.; Singh, R.; Saraya, A.; Chauhan, S.S.; Yadav, S.; Pal, S.; Singh, N.; Dey, S. p38β MAP kinase as a therapeutic target for pancreatic cancer. Chem. Biol. Drug Des. 2012, 80, 266–273. [Google Scholar] [CrossRef]
- Yao, Z.; Xu, R.; Yuan, L.; Xu, M.; Zhuang, H.; Li, Y.; Zhang, Y.; Lin, N. Circ_0001955 facilitates hepatocellular carcinoma (HCC) tumorigenesis by sponging miR-516a-5p to release TRAF6 and MAPK11. Cell Death Dis. 2019, 10, 945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Yuan, X.; Wang, D.; Barakat, B.; Williams, E.D.; Hannigan, G.E. Selective regulation of p38β protein and signaling by integrin-linked kinase mediates bladder cancer cell migration. Oncogene 2014, 33, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Browne, A.J.; Göbel, A.; Thiele, S.; Hofbauer, L.C.; Rauner, M.; Rachner, T.D. p38 MAPK regulates the Wnt inhibitor Dickkopf-1 in osteotropic prostate cancer cells. Cell Death Dis. 2016, 7, e2119. [Google Scholar] [CrossRef] [Green Version]
- Grossi, V.; Lucarelli, G.; Forte, G.; Peserico, A.; Matrone, A.; Germani, A.; Rutigliano, M.; Stella, A.; Bagnulo, R.; Loconte, D.; et al. Loss of STK11 expression is an early event in prostate carcinogenesis and predicts therapeutic response to targeted therapy against MAPK/p38. Autophagy 2015, 11, 2102–2113. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kong, C.; Wu, C.; Wang, Y.; Xu, B.; Liang, S.; Ying, X. Knocking down of LINC01220 inhibits proliferation and induces apoptosis of endometrial carcinoma through silencing MAPK11. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Meng, S.; Xu, Q.; Wang, X.; Wang, J.; Gong, R.; Song, Y.; Duan, Y.; Zhang, Y. Gene expression profiling of lung adenocarcinoma in Xuanwei, China. Eur. J. Cancer Prev. 2016, 25, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, I.; Riera, P.; Andrés, M.; Altés, A.; Majem, M.; Blanco, R.; Capdevila, L.; Barba, A.; Barnadas, A.; Salazar, J. Prognostic effect of VEGF gene variants in metastatic non-small-cell lung cancer patients. Angiogenesis 2019, 22, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Planchard, D.; Camara-Clayette, V.; Dorvault, N.; Soria, J.-C.; Fouret, P. p38 Mitogen-activated protein kinase signaling, ERCC1 expression, and viability of lung cancer cells from never or light smoker patients. Cancer 2012, 118, 5015–5025. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; He, J.; Liu, Z.; Xu, J.; Yi, S.F.; Liu, H.; Yang, J. MAPK11 in breast cancer cells enhances osteoclastogenesis and bone resorption. Biochimie 2014, 106, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Luo, Q.; Xiao, Y.; Li, H.; Kong, L.; Ren, G. The implication from RAS/RAF/ERK signaling pathway increased activation in epirubicin treated triple negative breast cancer. Oncotarget 2017, 8, 108249–108260. [Google Scholar] [CrossRef] [Green Version]
- Sahu, V.; Nigam, L.; Agnihotri, V.; Gupta, A.; Shekhar, S.; Subbarao, N.; Bhaskar, S.; Dey, S. Diagnostic Significance of p38 Isoforms (p38α, p38β, p38γ, p38δ) in Head and Neck Squamous Cell Carcinoma: Comparative Serum Level Evaluation and Design of Novel Peptide Inhibitor Targeting the Same. Cancer Res. Treat. 2019, 51, 313–325. [Google Scholar] [CrossRef]
- Cristóbal, I.; Garcia-Orti, L.; Cirauqui, C.; Cortes-Lavaud, X.; García-Sánchez, M.A.; Calasanz, M.J.; Odero, M.D. Overexpression of SET is a recurrent event associated with poor outcome and contributes to protein phosphatase 2A inhibition in acute myeloid leukemia. Haematologica 2012, 97, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Arriazu, E.; Vicente, C.; Pippa, R.; Peris, I.; Martínez-Balsalobre, E.; García-Ramírez, P.; Marcotegui, N.; Igea, A.; Alignani, D.; Rifón, J.; et al. A new regulatory mechanism of protein phosphatase 2A activity via SET in acute myeloid leukemia. Blood Cancer J. 2020, 10, 3. [Google Scholar] [CrossRef]
- Arriazu, E.; Pippa, R.; Odero, M.D. Protein Phosphatase 2A as a Therapeutic Target in Acute Myeloid Leukemia. Front. Oncol. 2016, 6, 78. [Google Scholar] [CrossRef] [Green Version]
- Bliss-Moreau, M.; Coarfa, C.; Gunaratne, P.H.; Guitart, J.; Krett, N.L.; Rosen, S.T. Identification of p38β as a therapeutic target for the treatment of Sézary syndrome. J. Investig. Dermatol. 2015, 135, 599–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, S.-Y.; Cheng, S.-Y.; Ng, S.-C.; Aneja, R.; Chen, C.-J.; Huang, C.-Y.; Kuo, W.-W. Roles of p38α and p38β mitogen-activated protein kinase isoforms in human malignant melanoma A375 cells. Int. J. Mol. Med. 2019, 44, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Chen, D.; Song, S.; Fu, X.; Wei, Y.; Lu, J.; Wang, L.; Wang, J. A genetic variation of the p38β promoter region is correlated with an increased risk of sporadic colorectal cancer. Oncol. Lett. 2013, 6, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimberg, J.; Olsen, R.S.; Skarstedt, M.; Löfgren, S.; Zar, N.; Matussek, A. Polymorphism of the p38β gene in patients with colorectal cancer. Oncol. Lett. 2014, 8, 1093–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Sun, D.; Zhao, S.; Xiong, H.; Fang, J. Screening of potential diagnostic markers and therapeutic targets against colorectal cancer. Onco Targets Ther. 2015, 8, 1691–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, J.; Igea, A.; Papaioannou, M.; Lopez-Casas, P.P.; Llonch, E.; Hidalgo, M.; Gorgoulis, V.G.; Nebreda, A.R. Pharmacological inhibition of p38 MAPK reduces tumor growth in patient-derived xenografts from colon tumors. Oncotarget 2015, 6, 8539–8551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelberg, D. Stress-activated protein kinases-tumor suppressors or tumor initiators? Semin. Cancer Biol. 2004, 14, 271–282. [Google Scholar] [CrossRef]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef]
- Loesch, M.; Chen, G. The p38 MAPK stress pathway as a tumor suppressor or more? Front. Biosci. A J. Virtual Libr. 2008, 13, 3581–3593. [Google Scholar] [CrossRef] [Green Version]
- Bulavin, D.V.; Phillips, C.; Nannenga, B.; Timofeev, O.; Donehower, L.A.; Anderson, C.W.; Appella, E.; Fornace, A.J. Inactivation of the Wip1 phosphatase inhibits mammary tumorigenesis through p38 MAPK-mediated activation of the p16(Ink4a)-p19(Arf) pathway. Nat. Genet. 2004, 36, 343–350. [Google Scholar] [CrossRef]
- Frey, M.R.; Dise, R.S.; Edelblum, K.L.; Polk, D.B. p38 kinase regulates epidermal growth factor receptor downregulation and cellular migration. EMBO J. 2006, 25, 5683–5692. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Leng, J.; Bian, D.; Mahanivong, C.; Carpenter, K.A.; Pan, Z.K.; Han, J.; Huang, S. Rac1-MKK3-p38-MAPKAPK2 pathway promotes urokinase plasminogen activator mRNA stability in invasive breast cancer cells. J. Biol. Chem. 2002, 277, 48379–48385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Cano, J.; Roche, O.; Cimas, F.J.; Pascual-Serra, R.; Ortega-Muelas, M.; Fernández-Aroca, D.M.; Sánchez-Prieto, R. p38MAPK and Chemotherapy: We Always Need to Hear Both Sides of the Story. Front. Cell Dev. Biol. 2016, 4, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cánovas, B.; Igea, A.; Sartori, A.A.; Gomis, R.R.; Paull, T.T.; Isoda, M.; Pérez-Montoyo, H.; Serra, V.; González-Suárez, E.; Stracker, T.H.; et al. Targeting p38α Increases DNA Damage, Chromosome Instability, and the Anti-tumoral Response to Taxanes in Breast Cancer Cells. Cancer Cell 2018, 33, 1094–1110.e8. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Roy, S.; Rana, A.; Akhter, Y.; Hande, M.P.; Banerjee, B. The role of p38 MAPK pathway in p53 compromised state and telomere mediated DNA damage response. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 836, 89–97. [Google Scholar] [CrossRef]
- Sui, X.; Kong, N.; Ye, L.; Han, W.; Zhou, J.; Zhang, Q.; He, C.; Pan, H. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014, 344, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Munshi, A.; Ramesh, R. Mitogen-activated protein kinases and their role in radiation response. Genes Cancer 2013, 4, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-H.; Liu, F.-H.; Lee, Y.-L.; Huang, H.-M. Interferon-alpha induces the growth inhibition of human T-cell leukaemia line Jurkat through p38alpha and p38beta. J. Biochem. 2010, 147, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, S.; Xiang, J.; Huang, S.; Lin, A. Induction of apoptosis by SB202190 through inhibition of p38beta mitogen-activated protein kinase. J. Biol. Chem. 1998, 273, 16415–16420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelfadil, E.; Cheng, Y.-H.; Bau, D.-T.; Ting, W.-J.; Chen, L.-M.; Hsu, H.-H.; Lin, Y.-M.; Chen, R.-J.; Tsai, F.-J.; Tsai, C.-H.; et al. Thymoquinone induces apoptosis in oral cancer cells through p38β inhibition. Am. J. Chin. Med. 2013, 41, 683–696. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roche, O.; Fernández-Aroca, D.M.; Arconada-Luque, E.; García-Flores, N.; Mellor, L.F.; Ruiz-Hidalgo, M.J.; Sánchez-Prieto, R. p38β and Cancer: The Beginning of the Road. Int. J. Mol. Sci. 2020, 21, 7524. https://doi.org/10.3390/ijms21207524
Roche O, Fernández-Aroca DM, Arconada-Luque E, García-Flores N, Mellor LF, Ruiz-Hidalgo MJ, Sánchez-Prieto R. p38β and Cancer: The Beginning of the Road. International Journal of Molecular Sciences. 2020; 21(20):7524. https://doi.org/10.3390/ijms21207524
Chicago/Turabian StyleRoche, Olga, Diego M. Fernández-Aroca, Elena Arconada-Luque, Natalia García-Flores, Liliana F. Mellor, María José Ruiz-Hidalgo, and Ricardo Sánchez-Prieto. 2020. "p38β and Cancer: The Beginning of the Road" International Journal of Molecular Sciences 21, no. 20: 7524. https://doi.org/10.3390/ijms21207524
APA StyleRoche, O., Fernández-Aroca, D. M., Arconada-Luque, E., García-Flores, N., Mellor, L. F., Ruiz-Hidalgo, M. J., & Sánchez-Prieto, R. (2020). p38β and Cancer: The Beginning of the Road. International Journal of Molecular Sciences, 21(20), 7524. https://doi.org/10.3390/ijms21207524