Proteomic Characterization of Urinary Extracellular Vesicles from Kidney-Transplanted Patients Treated with Calcineurin Inhibitors


 ,
, 
Abstract
:1. Introduction
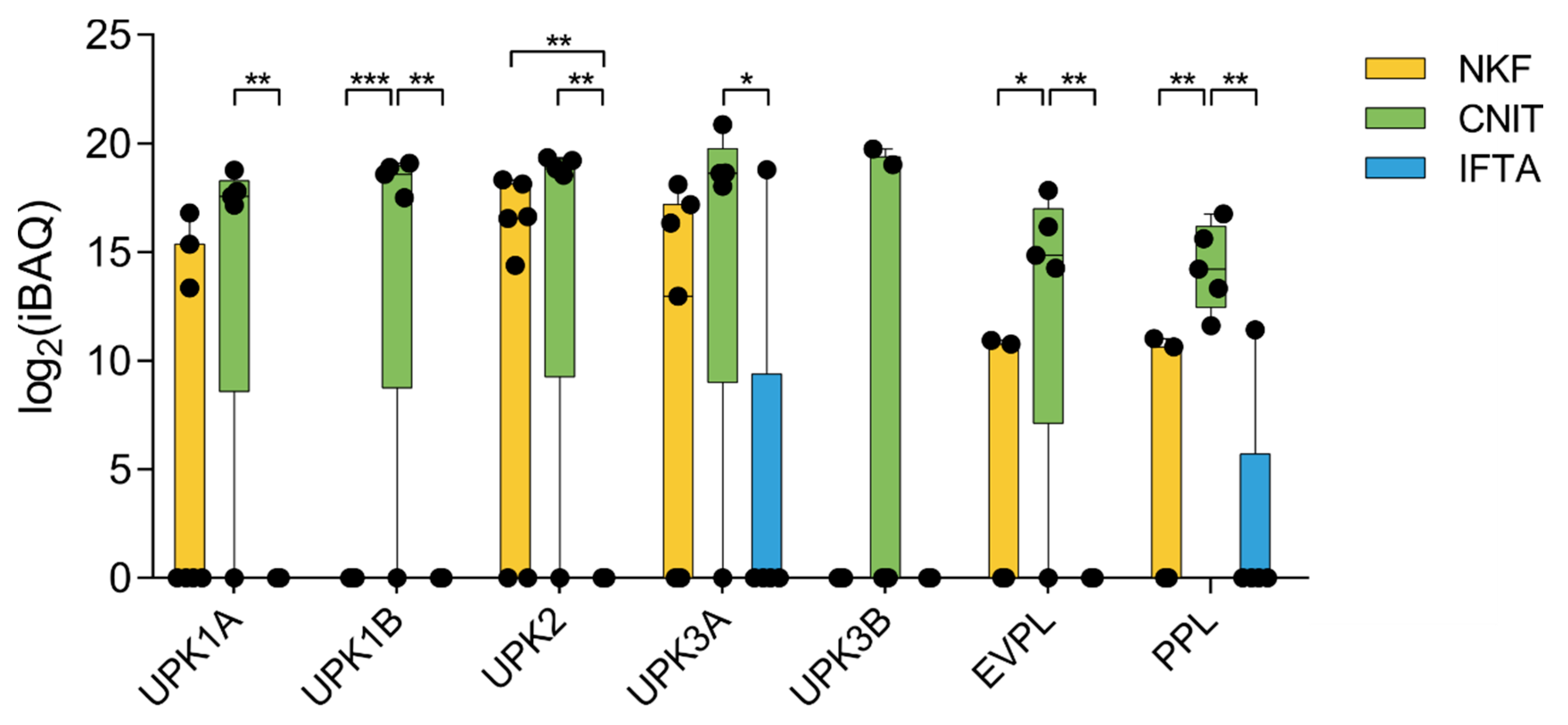
2. Results
2.1. Patients and Samples
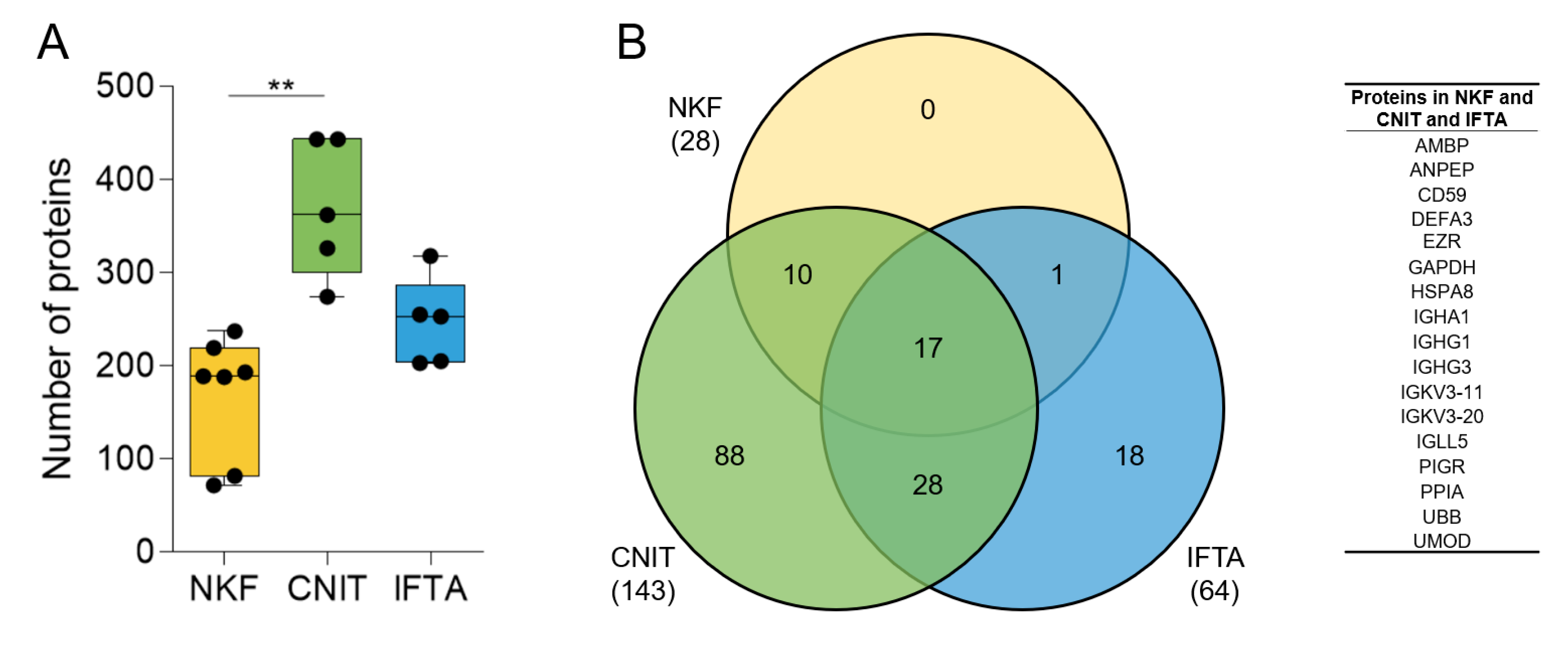
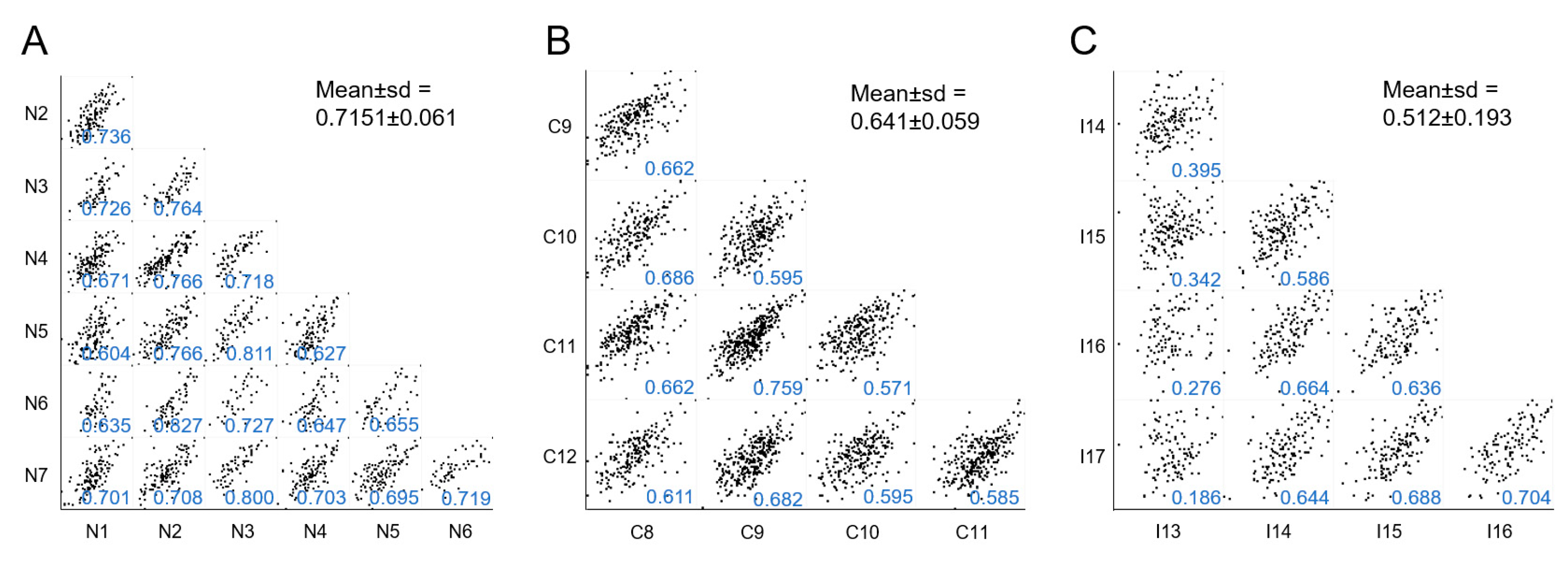
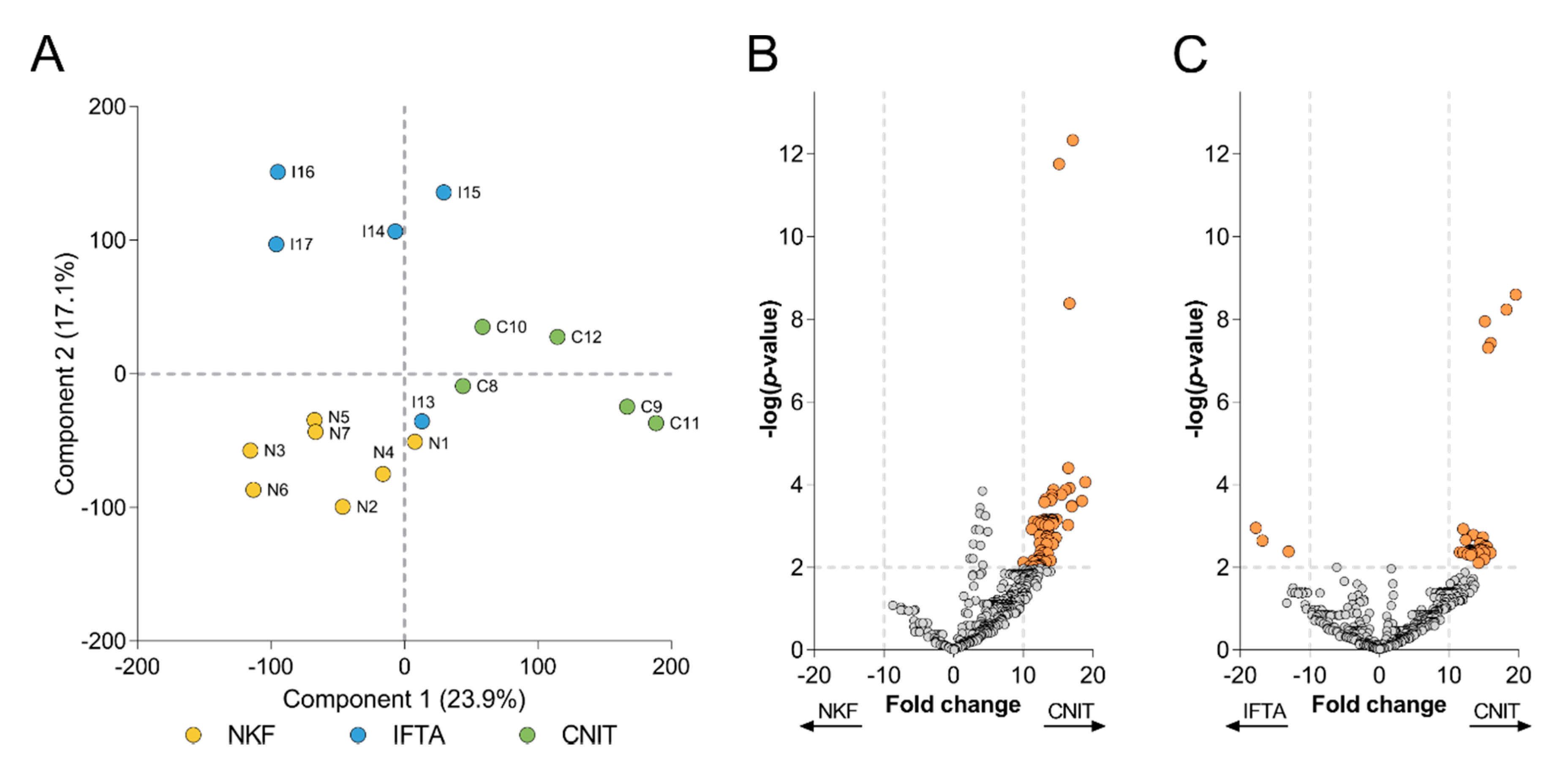
2.2. Global Analysis of the uEV Proteome
2.3. Differentially Expressed Proteins
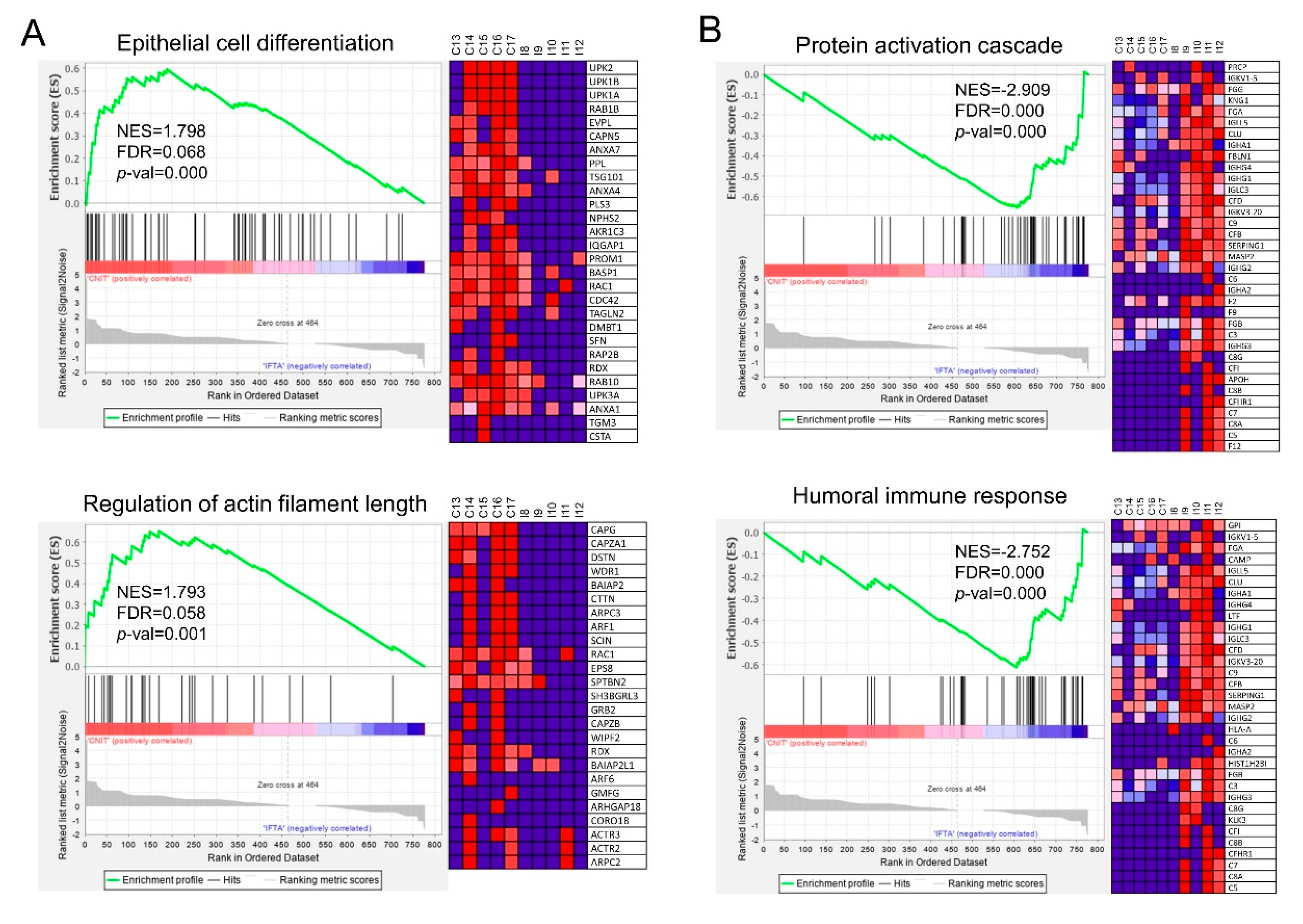
2.4. Biological Processes Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Patients and Pre-Processing of Samples
4.2. Samples Processing and uEV Isolation
4.3. Mass-Spectrometry
4.4. Proteomics and Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| aCNIT | acute calcineurin inhibitors toxicity |
| ANOVA | Analysis of variance |
| BP | Biological Process |
| CC | Cellular Component |
| cCNIT | chronic calcineurin inhibitors toxicity |
| CD | cluster of differentiation |
| CNI | calcineurin inhibitors |
| CNIT | calcineurin inhibitors toxicity |
| CSA | cyclosporine A |
| DM | diabetes mellitus type 2 |
| ES | enrichment score |
| F | female |
| FDR | false discovery rate |
| G | grade |
| GO | Gene Ontology |
| GSEA | Gene Set Enrichment Analysis |
| HT | arterial hypertension |
| iBAQ | Intensity-based absolute quantification |
| IFTA | interstitial fibrosis and tubular atrophy |
| IL | interleukin |
| IL | interleukin |
| IL2RA | interleukin 2 receptor antagonists |
| IS | immunosuppressive treatment |
| LC-MS/MS | liquid chromatography tandem mass-spectrometry |
| LD | living donor |
| M | male |
| MFI | mean fluorescence intensity |
| MMF | mycophenolate mofetil |
| MSigDB | Molecular Signatures Database |
| NES | normalized enrichment score |
| NKF | normal kidney function |
| PBS | phosphate-buffered saline |
| PCA | Principal Component Analysis |
| PR | prednisone |
| rATG | rabbit anti-thymocyte globulin |
| RT | Renal transplantation |
| SEC | size-exclusion chromatography |
| TAC | tacrolimus |
| uEV | Urinary Extracellular Vesicles |
References
- Heusler, K.; Pletscher, A. The controversial early history of cyclosporin. Swiss Med. Wkly. 2001, 131, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Calne, R. Cyclosporine as a milestone in immunosuppression. Transplant. Proc. 2004, 36, 13S–15S. [Google Scholar] [CrossRef]
- Kino, T.; Hatanaka, H.; Hashimoto, M.; Nishiyama, M.; Goto, T.; Okuhara, M.; Kohsaka, M.; Aoki, H.; Imanaka, H. FK-506, a novel immunosuppressant isolated from a Streptomyces. I. Fermentation, isolation, and physico-chemical and biological characteristics. J. Antibiot. 1987, 40, 1249–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatanaka, H.; Iwami, M.; Kino, T.; Goto, T.; Okuhara, M. FR-900520 and FR-900523, novel immunosuppressants isolated from a Streptomyces. I. Taxonomy of the producing strain. J. Antibiot. 1988, 41, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Starzl, T.E.; Todo, S.; Fung, J.; Demetris, A.J.; Venkataramman, R.; Jain, A. FK 506 for liver, kidney, and pancreas transplantation. Lancet 1989, 2, 1000–1004. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Martínez, S.; Redondo, J.M. Inhibitors of the calcineurin/NFAT pathway. Curr. Med. Chem. 2004, 11, 997–1007. [Google Scholar] [CrossRef]
- Andreoni, K.A.; Brayman, K.L.; Guidinger, M.K.; Sommers, C.M.; Sung, R.S. Kidney and pancreas transplantation in the United States, 1996–2005. Am. J. Transplant. 2007, 7, 1359–1375. [Google Scholar] [CrossRef] [Green Version]
- Azzi, J.R.; Sayegh, M.H.; Mallat, S.G. Calcineurin inhibitors: 40 years later, can’t live without …. J. Immunol. 2013, 191, 5785–5791. [Google Scholar] [CrossRef] [Green Version]
- Issa, N.; Kukla, A.; Ibrahim, H.N. Calcineurin inhibitor nephrotoxicity: A review and perspective of the evidence. Am. J. Nephrol. 2013, 37, 602–612. [Google Scholar] [CrossRef]
- Liptak, P.; Ivanyi, B. Primer: Histopathology of calcineurin-inhibitor toxicity in renal allografts. Nat. Clin. Pract. Nephrol. 2006, 2, 398–404. [Google Scholar] [CrossRef]
- Charney, D.A.; Bhaskaran, M.; Molmenti, E. Calcineurin inhibitor toxicity in a renal transplant recipient. Clin. Kidney J. 2009, 2, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Fogo, A.B.; Lusco, M.A.; Najafian, B.; Alpers, C.E. AJKD Atlas of renal pathology: Osmotic tubular injury. Am. J. Kidney Dis. 2017, 69, e11–e12. [Google Scholar] [CrossRef] [PubMed]
- Nankivell, B.J.; Fenton-Lee, C.A.; Kuypers, D.R.; Cheung, E.; Allen, R.D.; O’Connell, P.J.; Chapman, J.R. Effect of histological damage on long-term kidney transplant outcome. Transplantation 2001, 71, 515–523. [Google Scholar] [CrossRef]
- Leal, R.; Tsapepas, D.; Crew, R.J.; Dube, G.K.; Ratner, L.; Batal, I. Pathology of calcineurin and mammalian target of rapamycin inhibitors in kidney transplantation. Kidney Int. Rep. 2018, 3, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naesens, M.; Lerut, E.; Damme, B.V.; Vanrenterghem, Y.; Kuypers, D.R.J. Tacrolimus exposure and evolution of renal allograft histology in the first year after transplantation. Am. J. Transplant. 2007, 7, 2114–2123. [Google Scholar] [CrossRef] [PubMed]
- Serón, D.; Moreso, F.; Fulladosa, X.; Hueso, M.; Carrera, M.; Grinyó, J.M. Reliability of chronic allograft nephropathy diagnosis in sequential protocol biopsies. Kidney Int. 2002, 61, 727–733. [Google Scholar] [CrossRef] [Green Version]
- Kershner, R.P.; Fitzsimmons, W.E. Relationship of FK506 whole blood concentrations and efficacy and toxicity after liver and kidney transplantation. Transplantation 1996, 62, 920–926. [Google Scholar] [CrossRef]
- Hesselink, D.A.; Bouamar, R.; van Gelder, T. The pharmacogenetics of calcineurin inhibitor-related nephrotoxicity. Therap. Drug Monit. 2010, 32, 387–393. [Google Scholar] [CrossRef]
- Thölking, G.; Schütte-Nütgen, K.; Schmitz, J.; Rovas, A.; Dahmen, M.; Bautz, J.; Jehn, U.; Pavenstädt, H.; Heitplatz, B.; Van Marck, V.; et al. A low tacrolimus concentration/dose ratio increases the risk for the development of acute calcineurin inhibitor-induced nephrotoxicity. J. Clin. Med. 2019, 8, 1586. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Cosio, F.G.; Amer, H.; Grande, J.P.; Larson, T.S.; Stegall, M.D.; Griffin, M.D. Comparison of low versus high tacrolimus levels in kidney transplantation: Assessment of efficacy by protocol biopsies. Transplantation 2007, 83, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Ekberg, H.; Grinyó, J.; Nashan, B.; Vanrenterghem, Y.; Vincenti, F.; Voulgari, A.; Truman, M.; Nasmyth-Miller, C.; Rashford, M. Cyclosporine sparing with mycophenolate mofetil, daclizumab and corticosteroids in renal allograft recipients: The CAESAR Study. Am. J. Transplant. 2007, 7, 560–570. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Wu, J.; Zhu, J.; Lubman, D.M. Comparison of an Optimized Ultracentrifugation Method versus Size-Exclusion Chromatography for Isolation of Exosomes from Human Serum. J. Proteome Res. 2018, 17, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Benedikter, B.J.; Bouwman, F.G.; Vajen, T.; Heinzmann, A.C.A.; Grauls, G.; Mariman, E.C.; Wouters, E.F.M.; Savelkoul, P.H.; Lopez-Iglesias, C.; Koenen, R.R.; et al. Ultrafiltration combined with size exclusion chromatography efficiently isolates extracellular vesicles from cell culture media for compositional and functional studies. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Monguió-Tortajada, M.; Carreras-Planella, L.; Franquesa, M.; Beyer, K.; Borràs, F.E. Size-Exclusion Chromatography-based isolation minimally alters Extracellular Vesicles’ characteristics compared to precipitating agents. Sci. Rep. 2016, 6, 33641. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, A.; Haller, H.; Schmitt, R.; Schiffer, M.; Koenecke, C.; Strassburg, C.; Lehner, F.; Gottlieb, J.; Bara, C.; Becker, J.U.; et al. Biopsy-diagnosed renal disease in patients after transplantation of other organs and tissues. Am. J. Transplant. 2010, 10, 2017–2025. [Google Scholar] [CrossRef]
- Wang, X.; Wilkinson, R.; Kildey, K.; Potriquet, J.; Mulvenna, J.; Lobb, R.J.; Möller, A.; Cloonan, N.; Mukhopadhyay, P.; Kassianos, A.J.; et al. Unique molecular profile of exosomes derived from primary human proximal tubular epithelial cells under diseased conditions. J. Extracell. Vesicles 2017, 6, 1314073. [Google Scholar] [CrossRef]
- Khanna, A.; Plummer, M.; Bromberek, C.; Bresnahan, B.; Hariharan, S. Expression of TGF-beta and fibrogenic genes in transplant recipients with tacrolimus and cyclosporine nephrotoxicity. Kidney Int. 2002, 62, 2257–2263. [Google Scholar] [CrossRef] [Green Version]
- Farris, A.B.; Colvin, R.B. Renal interstitial fibrosis: Mechanisms and evaluation. Curr. Opin. Nephrol. Hypertens. 2012, 21, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Gewin, L.; Zent, R. How does TGF-β mediate tubulointerstitial fibrosis? Semin. Nephrol. 2012, 32, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Grgic, I.; Duffield, J.S.; Humphreys, B.D. The origin of interstitial myofibroblasts in chronic kidney disease. Pediatr. Nephrol. 2012, 27, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Carew, R.M.; Wang, B.; Kantharidis, P. The role of EMT in renal fibrosis. Cell Tissue Res. 2012, 347, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Puigmulé, M.; López-Hellin, J.; Suñé, G.; Tornavaca, O.; Camaño, S.; Tejedor, A.; Meseguer, A. Differential proteomic analysis of cyclosporine A-induced toxicity in renal proximal tubule cells. Nephrol. Dial. Transplant. 2009, 24, 2672–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Li, C.; Shan, J.; Luo, L.; Feng, L.; Lu, J.; Li, S.; Long, D.; Li, Y. Mechanisms of renal cell apoptosis induced by cyclosporine A: A systematic review of in vitro studies. Am. J. Nephrol. 2011, 33, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.; Woolf, A.S. Uroplakins: New molecular players in the biology of urinary tract malformations. Kidney Int. 2007, 71, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Min, G.; Wang, H.; Sun, T.-T.; Kong, X.-P. Structural basis for tetraspanin functions as revealed by the cryo-EM structure of uroplakin complexes at 6-Å resolution. J. Cell Biol. 2006, 173, 975–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karashima, T. Interaction of periplakin and envoplakin with intermediate filaments. J. Cell Sci. 2002, 115, 5027–5037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhrberg, C.; Hajibagheri, M.A.; Simon, M.; Dooley, T.P.; Watt, F.M. Envoplakin, a novel precursor of the cornified envelope that has homology to desmoplakin. J. Cell Biol. 1996, 134, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Ruhrberg, C.; Hajibagheri, M.A.N.; Parry, D.A.D.; Watt, F.M. Periplakin, a Novel Component of Cornified Envelopes and Desmosomes That Belongs to the Plakin Family and Forms Complexes with Envoplakin. J. Cell Biol. 1997, 139, 1835–1849. [Google Scholar] [CrossRef] [Green Version]
- Salih, M.; Demmers, J.A.; Bezstarosti, K.; Leonhard, W.N.; Losekoot, M.; van Kooten, C.; Gansevoort, R.T.; Peters, D.J.M.; Zietse, R.; Hoorn, E.J. Proteomics of urinary vesicles links plakins and complement to polycystic kidney disease. J. Am. Soc. Nephrol. 2016, 27, 3079–3092. [Google Scholar] [CrossRef] [Green Version]
- Sigdel, T.K.; Sarwal, M.M. Assessment of circulating protein signatures for kidney transplantation in pediatric recipients. Front. Med. 2017, 4, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarró, E.; Jacobs-Cachá, C.; Itarte, E.; Meseguer, A. A pharmacologically-based array to identify targets of cyclosporine A-induced toxicity in cultured renal proximal tubule cells. Toxicol. Appl. Pharmacol. 2012, 258, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, F.; Mestre, E.; Essig, M.; Sauvage, F.L.; Marquet, P.; Gastinel, L.N. Quantitative proteomic analysis of cyclosporine-induced toxicity in a human kidney cell line and comparison with tacrolimus. J. Proteom. 2011, 75, 677–694. [Google Scholar] [CrossRef] [PubMed]
- Lynöe, N.; Sandlund, M.; Dahlqvist, G.; Jacobsson, L. Informed consent: Study of quality of information given to participants in a clinical trial. BMJ 1991, 303, 610–613. [Google Scholar] [CrossRef] [Green Version]
- Monguió-Tortajada, M.; Morón-Font, M.; Gámez-Valero, A.; Carreras-Planella, L.; Borràs, F.E.; Franquesa, M. Extracellular-Vesicle Isolation from Different Biological Fluids by Size-Exclusion Chromatography. Curr. Protoc. Stem Cell Biol. 2019, 49, e82. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef] [Green Version]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Pathan, M.; Keerthikumar, S.; Ang, C.-S.; Gangoda, L.; Quek, C.Y.J.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef]
- Pathan, M.; Keerthikumar, S.; Chisanga, D.; Alessandro, R.; Ang, C.-S.; Askenase, P.; Batagov, A.O.; Benito-Martin, A.; Camussi, G.; Clayton, A.; et al. A novel community driven software for functional enrichment analysis of extracellular vesicles data. J. Extracell. Vesicles 2017, 6, 1321455. [Google Scholar] [CrossRef] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdottir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group, Sample | Age | Gender | DM | HT | Crea. | Prot. | Months from RT | LD | Donor Age |
|---|---|---|---|---|---|---|---|---|---|
| N1 | 64 | F | − | − | 0.86 | 99 | 137.9 | − | 53 |
| N2 | 65 | M | − | + | 0.86 | 110 | 186.6 | − | 37 |
| N3 | 45 | F | − | + | 0.82 | 30 | 113.6 | − | 43 |
| N4 | 42 | M | − | + | 0.89 | 187 | 186.6 | − | 30 |
| N5 | 57 | M | − | + | 0.9 | 92 | 166.4 | − | 45 |
| N6 | 65 | F | − | − | 0.85 | 55 | 70.0 | + | 37 |
| N7 | 69 | F | − | + | 1.14 | 86 | 57.8 | + | 58 |
| C8 | 55 | M | + | + | 2.29 | 506 | 0.5 | + | 62 |
| C9 | 33 | M | − | − | 1.93 | 232 | 2.8 | + | 59 |
| C10 | 49 | F | − | + | 3.08 | 427 | 238.8 | − | 45 |
| C11 | 50 | F | − | − | 2.49 | 207 | 5.5 | − | 60 |
| C12 | 41 | F | − | + | 1.80 | 76 | 25.7 | − | 34 |
| I13 | 50 | F | − | − | 2.00 | 62 | 21.7 | + | 48 |
| I14 | 64 | M | − | + | 1.60 | 1600 | 84.6 | − | 71 |
| I15 | 68 | M | − | + | 2.49 | 94 | 25.8 | − | 67 |
| I16 | 68 | M | + | + | 2.62 | 800 | 15.1 | − | 38 |
| I17 | 53 | F | − | + | 2.30 | 806 | 252.3 | − | 35 |
| Sig. | ns | ns | ns | ns | *** | ns | ns | ns | ns |
| p-value | 0.064 a | 0.784 b | 0.452 b | 0.784 b | <0.001 a | 0.093 a | 0.113 a | 0.784 b | 0.387 a |
| Group, Sample | Induction Treatment | IS at Urine Collection | High CNI | Diagnosis |
|---|---|---|---|---|
| N1 | IL2RA | PR, CSA | − | NKF |
| N2 | rATG | PR, TAC, MMF | − | NKF |
| N3 | IL2RA | PR, TAC | − | NKF |
| N4 | IL2RA | PR, TAC | − | NKF |
| N5 | IL2RA | PR, TAC | − | NKF |
| N6 | IL2RA | PR, TAC, MMF | − | NKF |
| N7 | IL2RA | PR, TAC, MMF | − | NKF |
| C8 | IL2RA | PR, TAC, MMF | + | aCNIT |
| C9 | IL2RA | PR, TAC, MMF | + | aCNIT |
| C10 | IL2RA | PR, CSA, MMF | + | cCNIT |
| C11 | IL2RA | PR, TAC, MMF | + | aCNIT |
| C12 | IL2RA | PR, TAC, MMF | + | aCNIT |
| I13 | IL2RA | PR, TAC, MMF | − | IFTA (G2) |
| I14 | IL2RA | PR, TAC, MMF | − | IFTA (G1) |
| I15 | IL2RA | PR, TAC, MMF | − | IFTA (G2) |
| I16 | IL2RA | PR, TAC, MMF | − | IFTA (G2) |
| I17 | IL2RA | PR, CSA | − | IFTA (G2) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreras-Planella, L.; Juega, J.; Taco, O.; Cañas, L.; Franquesa, M.; Lauzurica, R.; Borràs, F.E. Proteomic Characterization of Urinary Extracellular Vesicles from Kidney-Transplanted Patients Treated with Calcineurin Inhibitors. Int. J. Mol. Sci. 2020, 21, 7569. https://doi.org/10.3390/ijms21207569
Carreras-Planella L, Juega J, Taco O, Cañas L, Franquesa M, Lauzurica R, Borràs FE. Proteomic Characterization of Urinary Extracellular Vesicles from Kidney-Transplanted Patients Treated with Calcineurin Inhibitors. International Journal of Molecular Sciences. 2020; 21(20):7569. https://doi.org/10.3390/ijms21207569
Chicago/Turabian StyleCarreras-Planella, Laura, Javier Juega, Omar Taco, Laura Cañas, Marcella Franquesa, Ricardo Lauzurica, and Francesc Enric Borràs. 2020. "Proteomic Characterization of Urinary Extracellular Vesicles from Kidney-Transplanted Patients Treated with Calcineurin Inhibitors" International Journal of Molecular Sciences 21, no. 20: 7569. https://doi.org/10.3390/ijms21207569
APA StyleCarreras-Planella, L., Juega, J., Taco, O., Cañas, L., Franquesa, M., Lauzurica, R., & Borràs, F. E. (2020). Proteomic Characterization of Urinary Extracellular Vesicles from Kidney-Transplanted Patients Treated with Calcineurin Inhibitors. International Journal of Molecular Sciences, 21(20), 7569. https://doi.org/10.3390/ijms21207569