Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit during Gestation

 , and
, and 
Abstract
:1. Introduction
2. Results
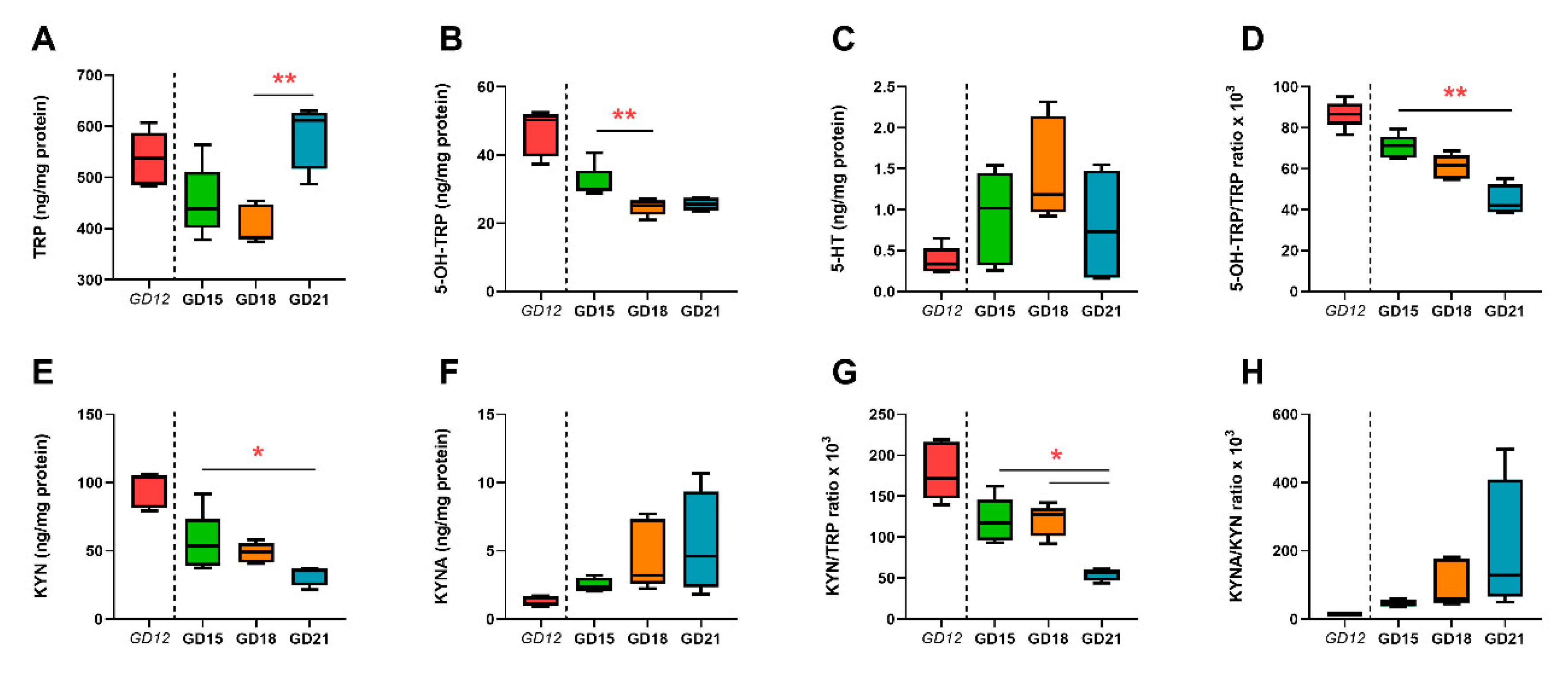
2.1. Basal Levels of TRP, KYN, KYNA, 5-OH-TRP, and 5-HT in the Maternal Plasma and Placenta
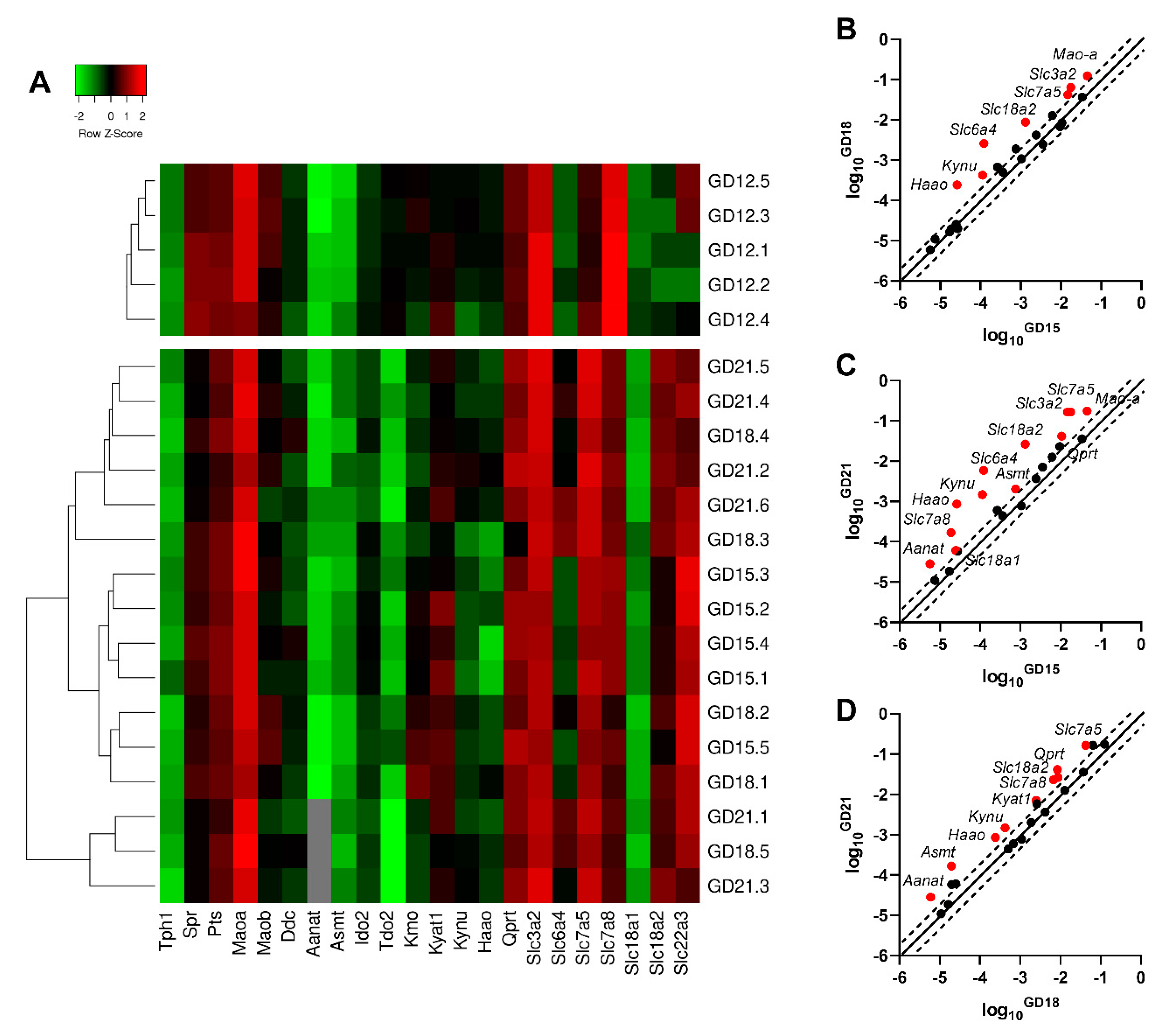
2.2. Gene Expression of Enzymes and Transporters Involved in TRP Metabolic Pathways in the Rat Placenta
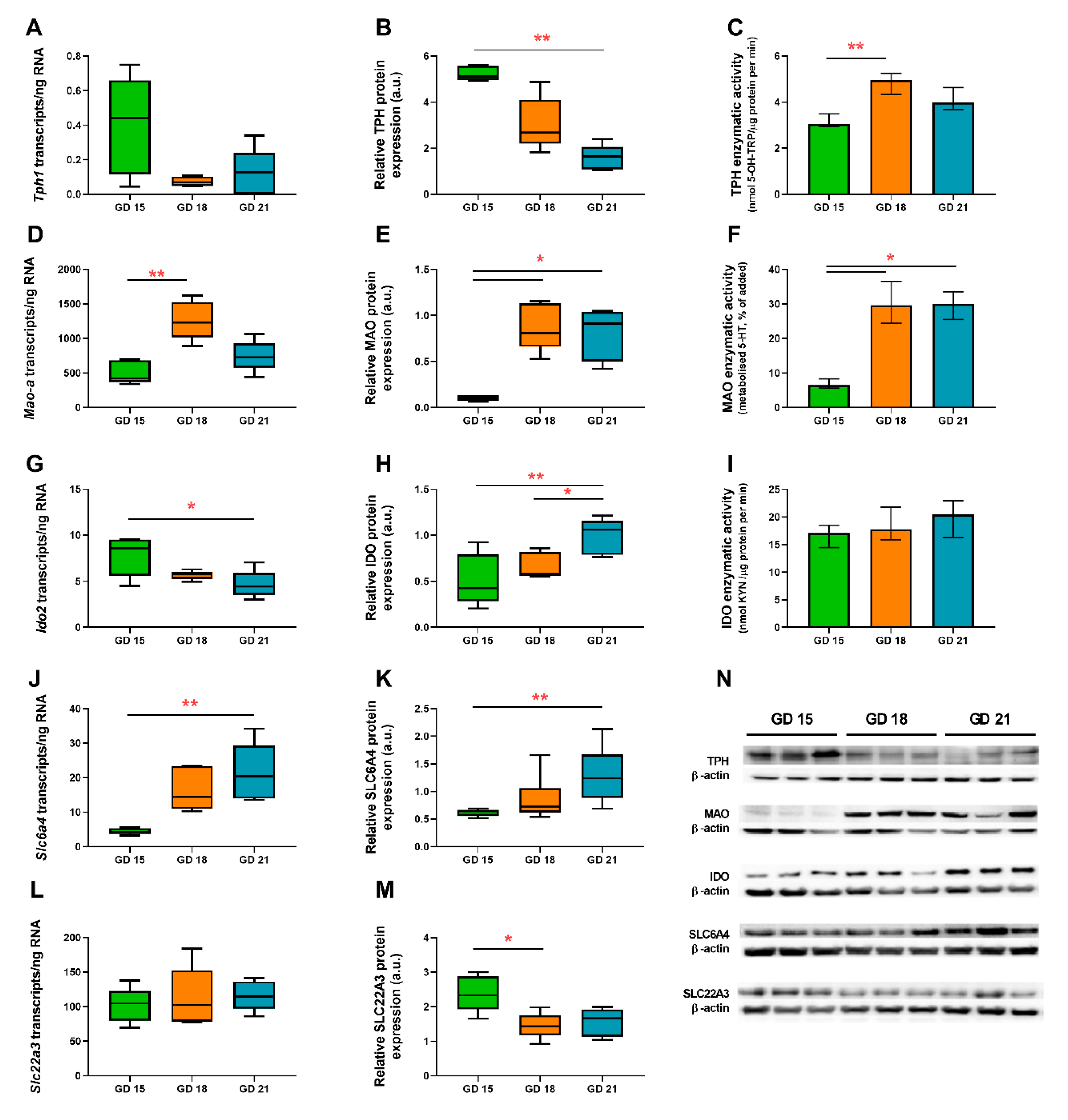
2.3. Absolute Quantification of Tph1, Mao-a, Slc6a4, Slc22a3, and Ido2 Gene Expression in the Rat Placenta during Gestation
2.4. Protein Analysis of TPH, MAO, SLC6A4, SLC22A3, and IDO in the Rat Placenta during Gestation
2.5. In Vitro Functional Assays of TPH, MAO, and IDO in the Rat Placenta
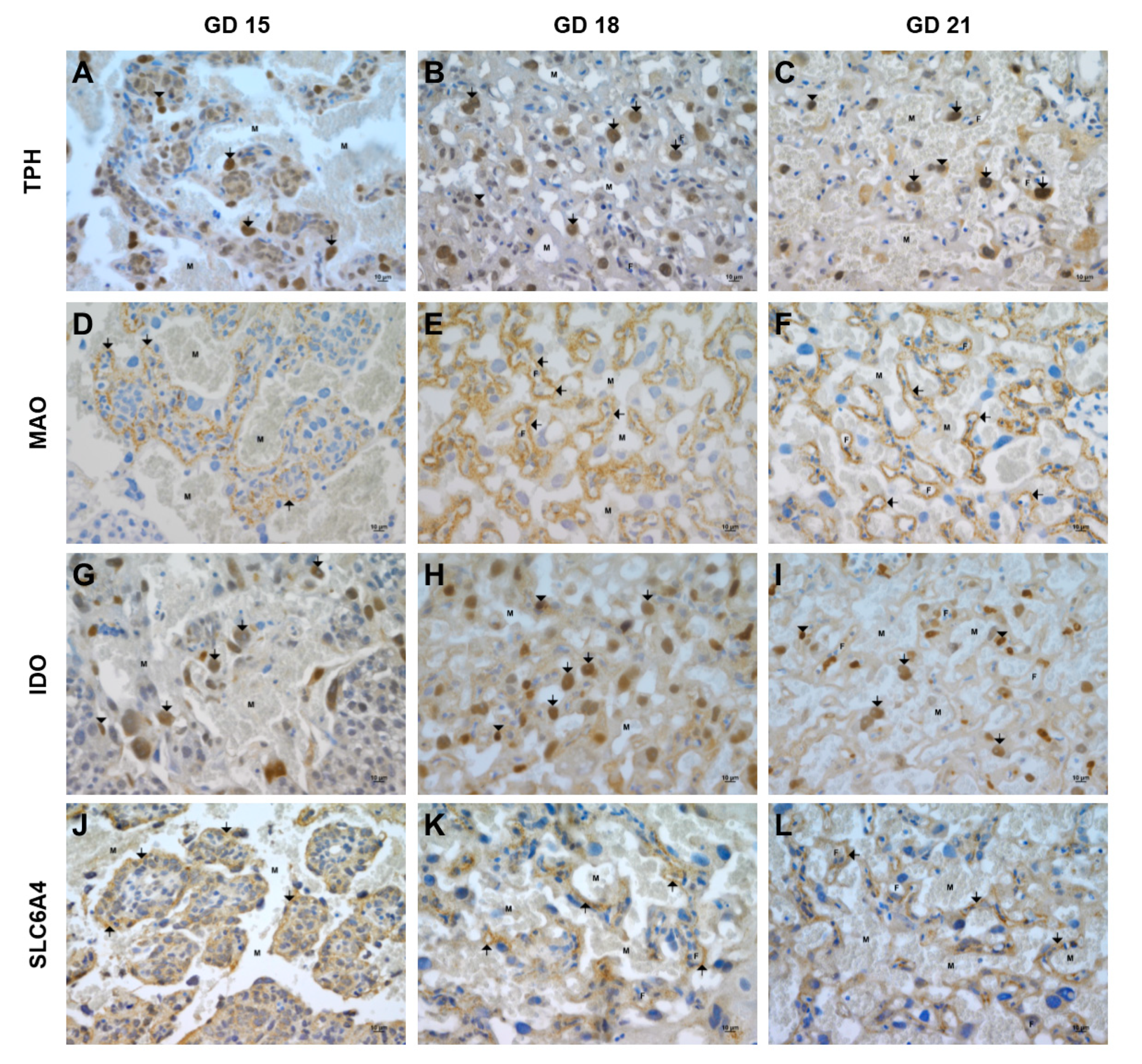
2.6. Immunohistochemical Localization of TPH, MAO, IDO, and SLC6A4 in the Rat Placenta at Different Stages of Gestation
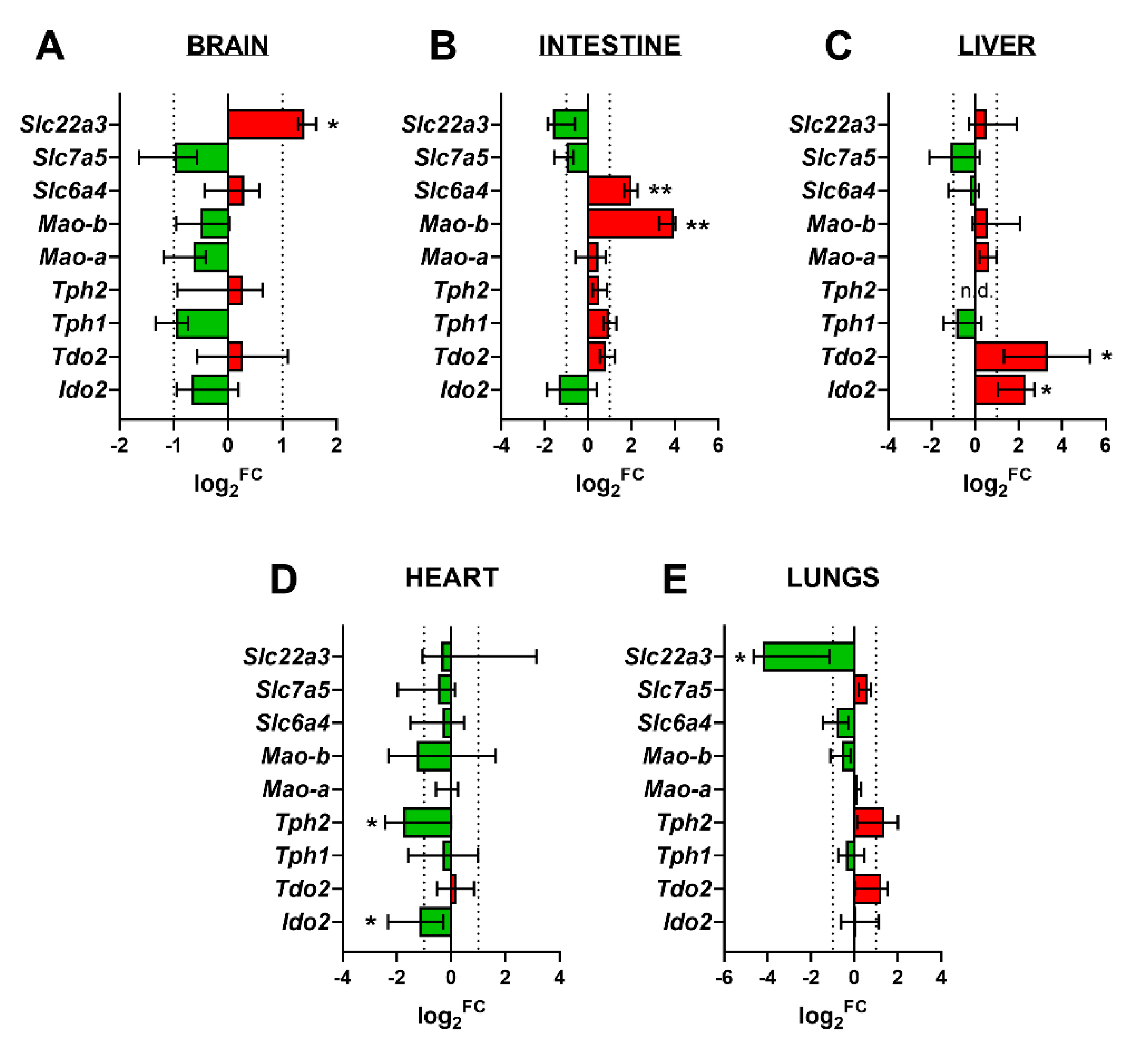
2.7. Gene Expression of Key Enzymes/Transporters of TRP Metabolic Pathways in Fetal Organs
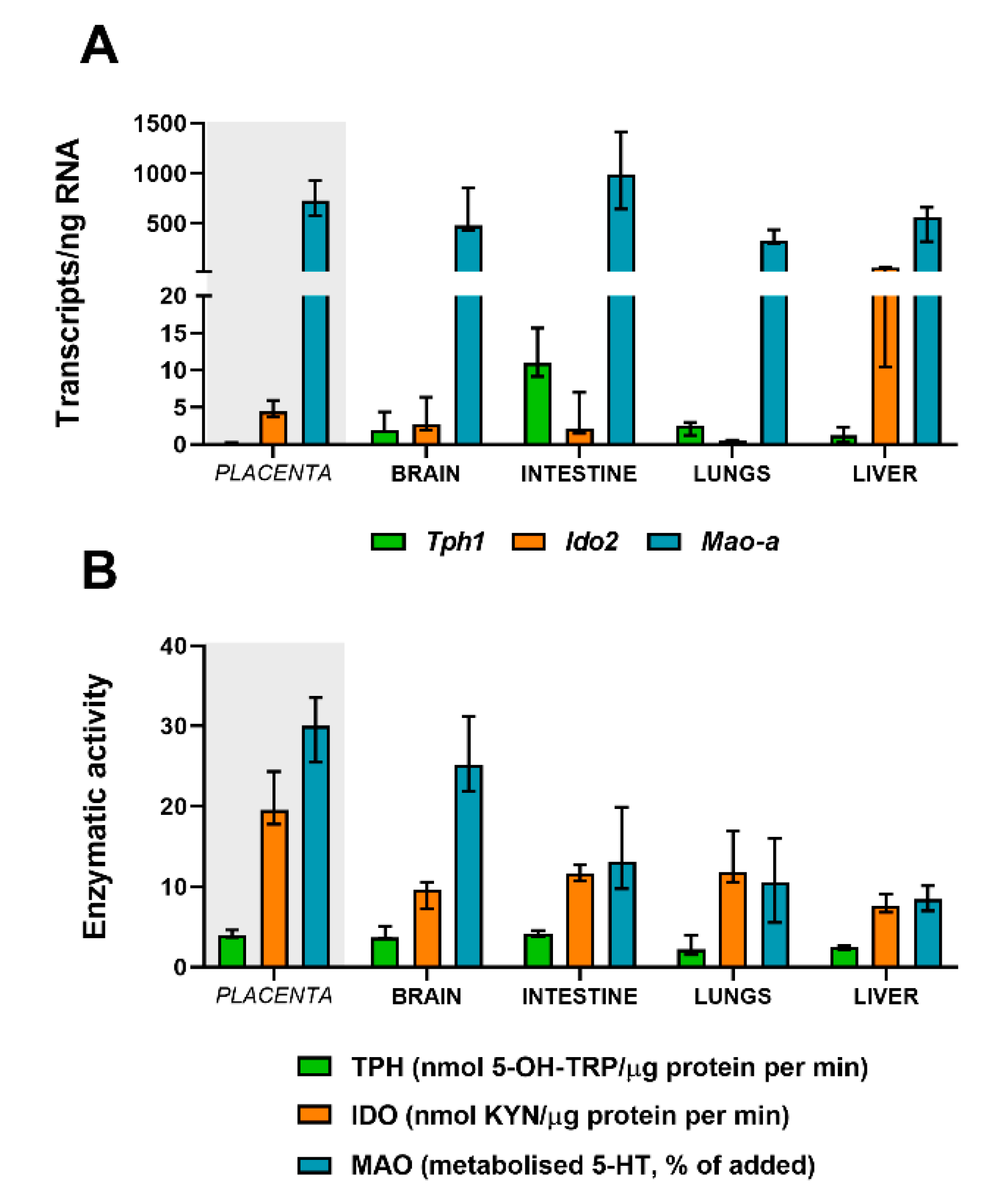
2.8. Absolute Quantification of Tph, Mao, and Ido in Fetal Organs at Term
2.9. Functional Activity of TPH, MAO, and IDO in Fetal Organs at Term
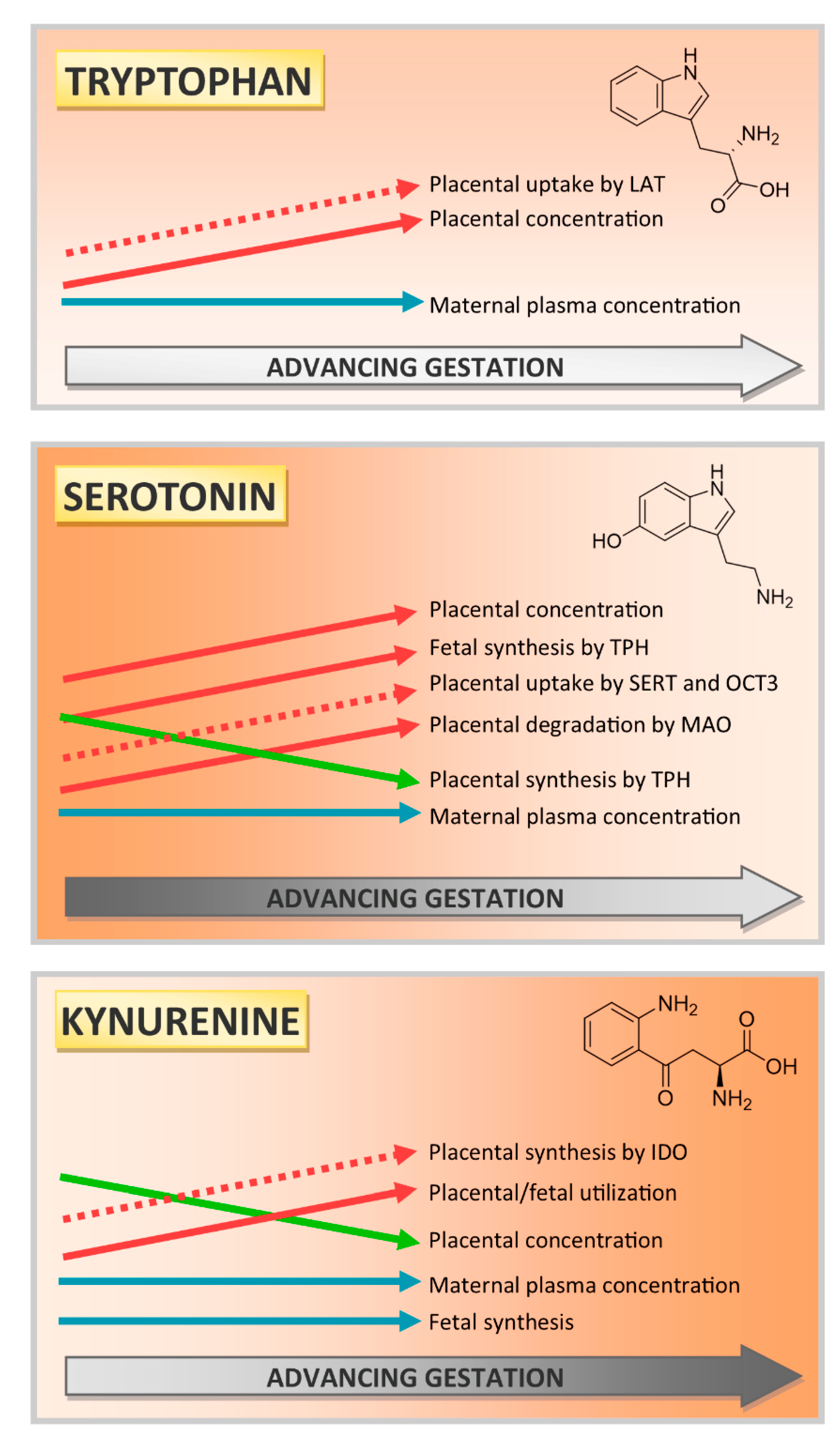
3. Discussion and Conclusions
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Rat Placenta and Fetal Organ Sample Collection
4.3. RNA Isolation, Reverse Transcription, and Quantitative PCR Analysis
4.4. Droplet Digital PCR Assay
4.5. Preparation of Tissue Homogenates
4.6. Western Blot Analysis
4.7. 5-HT Metabolism by MAO in the Placenta and Fetal Organs
4.8. TRP Metabolism by IDO in the Placenta and Fetal Organs
4.9. TRP Metabolism by TPH in the Placenta and Fetal Organs
4.10. Extraction of TRP and Its Metabolites from Biological Samples
4.11. HPLC Analysis of TRP, 5-HT, 5-OH-TRP, KYN, and KYNA
4.12. Immunohistochemical Characterization of IDO, TPH, MAO, and SERT in the Rat Placenta
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-HT | 5-hydroxytryptamine (serotonin) |
| 5-OH-TRP | 5-hydroxy-l-tryptophan |
| AANAT | aralkylamine N-acetyltransferase |
| ASMT | acetylserotonin O-methyltransferase |
| BH4 | tetrahydrobiopterin |
| DDC | dopa decarboxylase |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| GD | gestational day |
| HAAO | 3-hydroxyanthranilate 3,4-dioxygenase |
| IDO | indoleamine 2,3-dioxygenase |
| KMO | kynurenine 3-monooxygenase |
| KYAT1 | kynurenine aminotransferase 1 |
| KYN | kynurenine |
| KYNA | kynurenic acid |
| KYNU | kynureninase |
| LAT | L-amino acid transporter |
| MAO | monoamine oxidase |
| NMDA | N-methyl-D-aspartate |
| OCT3 | organic cation transporter 3 |
| PTS | 6-pyruvoyltetrahydropterin synthase |
| QPRT | quinolinate phosphoribosyltransferase |
| QUIN | quinolinic acid |
| SERT | serotonin transporter |
| SPR | sepiapterin reductase |
| TDO | tryptophan 2,3-dioxygenase |
| TPH | tryptophan hydroxylase |
| TRP | tryptophan |
| YWHAZ | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta |
References
- Sedlmayr, P.; Blaschitz, A.; Stocker, R. The Role of Placental Tryptophan Catabolism. Front. Immunol. 2014, 5, 230. [Google Scholar] [CrossRef] [Green Version]
- Yudilevich, D.L.; Sweiry, J.H. Transport of amino acids in the placenta. Biochim. Biophys. Acta (BBA)—Rev. Biomembr. 1985, 822, 169–201. [Google Scholar] [CrossRef]
- Badawy, A.A.-B. Tryptophan metabolism, disposition and utilization in pregnancy. Biosci. Rep. 2015, 35, e00261. [Google Scholar] [CrossRef] [PubMed]
- Teshigawara, T.; Mouri, A.; Kubo, H.; Nakamura, Y.; Shiino, T.; Okada, T.; Morikawa, M.; Nabeshima, T.; Ozaki, N.; Yamamoto, Y.; et al. Changes in tryptophan metabolism during pregnancy and postpartum periods: Potential involvement in postpartum depressive symptoms. J. Affect. Disord. 2019, 255, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Zardoya-Laguardia, P.; Blaschitz, A.; Hirschmugl, B.; Lang, I.; A Herzog, S.; Nikitina, L.; Gauster, M.; Hausler, M.; Cervar-Zivkovic, M.; Karpf, E.; et al. Endothelial indoleamine 2,3-dioxygenase-1 regulates the placental vascular tone and is deficient in intrauterine growth restriction and pre-eclampsia. Sci. Rep. 2018, 8, 5488. [Google Scholar] [CrossRef] [PubMed]
- Alwasel, S.H.; Bagby, S.P.; Boyd, R.; Boyd, R.; Burdge, G.; Carter, A.M.; Cetin, I.; Cole, Z.; Cooper, C.; Critchley, H.; et al. The Placenta and Human Developmental Programming; Cambridge University Press (CUP): Cambridge, UK, 2009. [Google Scholar]
- Staud, F.; Karahoda, R. Trophoblast: The central unit of fetal growth, protection and programming. Int. J. Biochem. Cell Biol. 2018, 105, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Billett, E.E. Monoamine Oxidase (MAO) in Human Peripheral Tissues. Neurotoxicology 2004, 25, 139–148. [Google Scholar] [CrossRef]
- Laurent, L.; DeRoy, K.; St-Pierre, J.; Côté, F.; Sanderson, J.T.; Vaillancourt, C. Human placenta expresses both peripheral and neuronal isoform of tryptophan hydroxylase. Biochimie 2017, 140, 159–165. [Google Scholar] [CrossRef]
- Viau, M.; Lafond, J.; Vaillancourt, C. Expression of placental serotonin transporter and 5-HT 2A receptor in normal and gestational diabetes mellitus pregnancies. Reprod. Biomed. Online 2009, 19, 207–215. [Google Scholar] [CrossRef]
- Mitchell, J.A. Serotonin-induced disruption of implantation in the rat: I. Serum progesterone, implantation site blood flow, and intrauterine pO2. Biol. Reprod. 1983, 28, 830–835. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.A. Serotonin-induced disruption of implantation in the rat: II. Suppression of decidualization. Biol. Reprod. 1983, 29, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnin, A.; Levitt, P. Fetal, maternal, and placental sources of serotonin and new implications for developmental programming of the brain. Neuroscience 2011, 197, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavarone, M.S.; Shuey, D.L.; Tamir, H.; Sadler, T.W.; Lauder, J.M. Serotonin and cardiac morphogenesis in the mouse embryo. Teratology 1993, 47, 573–584. [Google Scholar] [CrossRef]
- Côté, F.; Fligny, C.; Bayard, E.; Launay, J.-M.; Gershon, M.D.; Mallet, J.; Vodjdani, G. Maternal serotonin is crucial for murine embryonic development. Proc. Natl. Acad. Sci. USA 2006, 104, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nat. Cell Biol. 2011, 472, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J.; Quaratella, S.B.; Setaro, A.C.; Siegman, E.C.; Subha, Z.T.; Tal, R.; Milano, K.M.; Steck, T.L. Pathway of Maternal Serotonin to the Human Embryo and Fetus. Endocrinology 2018, 159, 1609–1629. [Google Scholar] [CrossRef]
- Arevalo, R.; Afonso, D.; Castro, R.; Rodriguez, M. Fetal brain serotonin synthesis and catabolism is under control by mother intake of tryptophan. Life Sci. 1991, 49, 53–66. [Google Scholar] [CrossRef]
- Karahoda, R.; Horackova, H.; Kastner, P.; Matthios, A.; Cerveny, L.; Kucera, R.; Kacerovsky, M.; Duintjer Tebbens, J.; Bonnin, A.; Abad, C.; et al. Serotonin homeostasis in the materno-foetal interface at term: Role of transporters (SERT/SLC6A4 and OCT3/SLC22A3) and monoamine oxidase A (MAO-A) in uptake and degradation of serotonin by human and rat term placenta. Acta Physiol. 2020, 229, e13478. [Google Scholar] [CrossRef]
- Ranzil, S.; Walker, D.W.; Borg, A.J.; Wallace, E.M.; Ebeling, P.R.; Murthi, P. The relationship between the placental serotonin pathway and fetal growth restriction. Biochimie 2018, 161, 80–87. [Google Scholar] [CrossRef]
- Robson, J.M.; Sullivan, F.M. Mechanism of lethal action of 5-hydroxytryptamine on the foetus. J. Endocrinol. 1963, 25, 553–554. [Google Scholar] [CrossRef]
- Munn, D.H.; Zhou, M.; Attwood, J.T.; Bondarev, I.; Conway, S.J.; Marshall, B.; Brown, C.; Mellor, A.L. Prevention of Allogeneic Fetal Rejection by Tryptophan Catabolism. Science 1998, 281, 1191–1193. [Google Scholar] [CrossRef] [PubMed]
- Uyttenhove, C.; Pilotte, L.; Théate, I.; Stroobant, V.; Colau, D.; Parmentier, N.; Boon, T.; Eynde, B.J.V.D. Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase. Nat. Med. 2003, 9, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, F.; Kuroiwa, T.; Takikawa, O.; Kido, R. Human indolylamine 2,3-dioxygenase. Its tissue distribution, and characterization of the placental enzyme. Biochem. J. 1985, 230, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Manuelpillai, U.; Wallace, E.M.; Nicholls, T.; Guillemin, G.; Phillips, D.J.; Walker, D. Increased mRNA Expression of Kynurenine Pathway Enzymes in Human Placentae Exposed to Bacterial Endotoxin. Adv. Exp. Med. Biol. 2003, 527, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Ligam, P.; Manuelpillai, U.; Wallace, E.; Walker, D. Localisation of Indoleamine 2,3-dioxygenase and Kynurenine Hydroxylase in the Human Placenta and Decidua: Implications for Role of the Kynurenine Pathway in Pregnancy. Placenta 2005, 26, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.C.; Vezzani, A.; French, E.D.; Schwarcz, R. Kynurenic acid blocks neurotoxicity and seizures induced in rats by the related brain metabolite quinolinic acid. Neurosci. Lett. 1984, 48, 273–278. [Google Scholar] [CrossRef]
- Pocivavsek, A.; Thomas, M.A.R.; Elmer, G.I.; Bruno, J.P.; Schwarcz, R. Continuous kynurenine administration during the prenatal period, but not during adolescence, causes learning and memory deficits in adult rats. Psychopharmacology 2014, 231, 2799–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pershing, M.L.; Bortz, D.M.; Pocivavsek, A.; Fredericks, P.J.; Jørgensen, C.V.; Vunck, S.A.; Leuner, B.; Schwarcz, R.; Bruno, J. Elevated levels of kynurenic acid during gestation produce neurochemical, morphological, and cognitive deficits in adulthood: Implications for schizophrenia. Neuropharmacol. 2015, 90, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Forrest, C.; Khalil, O.; Pisar, M.; McNair, K.; Kornisiuk, E.; Snitcofsky, M.; Gonzalez, N.; Jerusalinsky, D.; Darlington, L.; Stone, T.W. Changes in synaptic transmission and protein expression in the brains of adult offspring after prenatal inhibition of the kynurenine pathway. Neuroscience 2013, 254, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Alexander, K.S.; Pocivavsek, A.; Wu, H.-Q.; Pershing, M.L.; Schwarcz, R.; Bruno, J. Early Developmental Elevations of Brain Kynurenic Acid Impair Cognitive Flexibility in Adults: Reversal with Galantamine. Neuroscience 2013, 238, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Goeden, N.; Notarangelo, F.M.; Pocivavsek, A.; Beggiato, S.; Bonnin, A.; Schwarcz, R. Prenatal Dynamics of Kynurenine Pathway Metabolism in Mice: Focus on Kynurenic Acid. Dev. Neurosci. 2017, 39, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Karahoda, R.; Abad, C.; Horackova, H.; Kastner, P.; Zaugg, J.; Cerveny, L.; Kucera, R.; Albrecht, C.; Staud, F. Dynamics of Tryptophan Metabolic Pathways in Human Placenta and Placental-Derived Cells: Effect of Gestation Age and Trophoblast Differentiation. Front. Cell Dev. Biol. 2020, 8, 937. [Google Scholar] [CrossRef]
- Mateos, S.S.; Sánchez, C.L.; Paredes, S.D.; Barriga, C.; Rodríguez, A.B. Circadian Levels of Serotonin in Plasma and Brain after Oral Administration of Tryptophan in Rats. Basic Clin. Pharmacol. Toxicol. 2009, 104, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Kubesova, A.; Tejkalova, H.; Syslova, K.; Kacer, P.; Vondrousova, J.; Tyls, F.; Fujakova, M.; Palenicek, T.; Horacek, J. Biochemical, Histopathological and Morphological Profiling of a Rat Model of Early Immune Stimulation: Relation to Psychopathology. PLoS ONE 2015, 10, e0115439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, J.M.; Senior, J.B. THE 5-HYDROXYTRYPTAMINE CONTENT OF THE PLACENTA AND FOETUS DURING PREGNANCY IN MICE. Br. J. Pharmacol. Chemother. 1964, 22, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Sano, M.; Ferchaud-Roucher, V.; Kaeffer, B.; Poupeau, G.; Castellano, B.; Darmaun, D. Maternal and fetal tryptophan metabolism in gestating rats: Effects of intrauterine growth restriction. Amino Acids 2015, 48, 281–290. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2019, 72, 253–319. [Google Scholar] [CrossRef]
- Ahmadimoghaddam, D.; Hofman, J.; Zemankova, L.; Nachtigal, P.; Dolezelova, E.; Cerveny, L.; Ceckova, M.; Micuda, S.; Staud, F. Synchronized activity of organic cation transporter 3 (Oct3/Slc22a3) and multidrug and toxin extrusion 1 (Mate1/Slc47a1) transporter in transplacental passage of MPP+ in rat. Toxicol. Sci. 2012, 128, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Ahmadimoghaddam, D.; Staud, F. Transfer of metformin across the rat placenta is mediated by organic cation transporter 3 (OCT3/SLC22A3) and multidrug and toxin extrusion 1 (MATE1/SLC47A1) protein. Reprod. Toxicol. 2013, 39, 17–22. [Google Scholar] [CrossRef]
- Wu, H.-H.; Choi, S.; Levitt, P. Differential patterning of genes involved in serotonin metabolism and transport in extra-embryonic tissues of the mouse. Placenta 2016, 42, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Kono, H.; Lin, Y.C.; Yamaguchi, M.; Zuspan, F.P.; O’Shaughnessy, R.W.; Lee, A.C.; Furuhashi, N.; Yokaichiya, T.; Takayama, K.; Yajima, A. Monoamine Oxidase Activity in Rat Organs during Pregnancy. Tohoku J. Exp. Med. 1994, 172, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Klein, D.C.; Robinson, J.C. Monoamine oxidase in rat placenta, human placenta, and cultured choriocarcinoma. Reproduction 1976, 46, 477–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadimoghaddam, D.; Zemankova, L.; Nachtigal, P.; Dolezelova, E.; Neumanova, Z.; Cerveny, L.; Ceckova, M.; Kacerovsky, M.; Micuda, S.; Staud, F. Organic cation transporter 3 (OCT3/SLC22A3) and multidrug and toxin extrusion 1 (MATE1/SLC47A1) transporter in the placenta and fetal tissues: Expression profile and fetus protective role at different stages of gestation. Biol. Reprod. 2013, 88, 55. [Google Scholar] [CrossRef] [PubMed]
- Verhaagh, S.; Barlow, D.P.; Zwart, R. The extraneuronal monoamine transporter Slc22a3/Orct3 co-localizes with the Maoa metabolizing enzyme in mouse placenta. Mech. Dev. 2001, 100, 127–130. [Google Scholar] [CrossRef]
- Behura, S.K.; Dhakal, P.; Kelleher, A.M.; Balboula, A.; Patterson, A.; Spencer, T.E. The brain-placental axis: Therapeutic and pharmacological relevancy to pregnancy. Pharmacol. Res. 2019, 149, 104468. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Author response for “The placenta-brain-axis”. J. Neurosci. Res 2020. [Google Scholar] [CrossRef]
- Linask, K.K. The Heart-Placenta Axis in the First Month of Pregnancy: Induction and Prevention of Cardiovascular Birth Defects. J. Pregnancy 2013, 2013, 320413. [Google Scholar] [CrossRef] [Green Version]
- Maslen, C. Recent Advances in Placenta–Heart Interactions. Front. Physiol. 2018, 9, 735. [Google Scholar] [CrossRef] [Green Version]
- Lewinsohn, R.; Glover, V.; Sandler, M. Beta-phenylethylamine and benzylamine as substrates for human monoamine oxidase A: A source of some anomalies? Biochem. Pharm. 1980, 29, 777–781. [Google Scholar] [CrossRef]
- Kono, H.; Lin, Y.C.; Gu, Y.; Yamaguchi, M.; Zuspan, F.P.; Furuhashi, N.; Takayama, K.; Yajima, A. Gossypol effects on monoamine oxidase(MAO) activity in several organs of term rats. Tohoku J. Exp. Med. 1991, 163, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Blaschitz, A.; Gauster, M.; Fuchs, D.; Lang, I.; Maschke, P.; Ulrich, D.; Karpf, E.; Takikawa, O.; Schimek, M.G.; Dohr, G.; et al. Vascular Endothelial Expression of Indoleamine 2,3-Dioxygenase 1 Forms a Positive Gradient towards the Feto-Maternal Interface. PLoS ONE 2011, 6, e21774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notarangelo, F.M.; Beggiato, S.; Schwarcz, R. Assessment of Prenatal Kynurenine Metabolism Using Tissue Slices: Focus on the Neosynthesis of Kynurenic Acid in Mice. Dev. Neurosci. 2019, 41, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Milart, P.; Urbanska, E.M.; Turski, W.A.; Paszkowski, T.; Sikorski, R. Kynurenine Aminotransferase I Activity in Human Placenta. Placenta 2001, 22, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Guillemin, G.J.; Mansour, H.; Chan-Ling, T.; Grant, R.S. Changes in kynurenine pathway metabolism in the brain, liver and kidney of aged female Wistar rats. FEBS J. 2011, 278, 4425–4434. [Google Scholar] [CrossRef]
- Roper, M.D.; Franz, J.M. Glucocorticoid control of the development of tryptophan oxygenase in the young rat. J. Biol. Chem. 1977, 252, 4354–4360. [Google Scholar] [PubMed]
- Notarangelo, F.M.; Schwarcz, R. Restraint Stress during Pregnancy Rapidly Raises Kynurenic Acid Levels in Mouse Placenta and Fetal Brain. Dev. Neurosci. 2016, 38, 458–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorn, A.M. Liver Development; StemBook: Cambridge, MA, USA, 2008. [Google Scholar]
- Houwing, D.J.; Buwalda, B.; Van Der Zee, E.A.; De Boer, S.F.; Olivier, J. The Serotonin Transporter and Early Life Stress: Translational Perspectives. Front. Cell. Neurosci. 2017, 11, 117. [Google Scholar] [CrossRef] [Green Version]
- Sjaarda, C.P.; Hecht, P.; McNaughton, A.J.M.; Zhou, A.; Hudson, M.L.; Will, M.J.; Smith, G.; Ayub, M.; Liang, P.; Chen, N.; et al. Interplay between maternal Slc6a4 mutation and prenatal stress: A possible mechanism for autistic behavior development. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sakata, T.; Anzai, N.; Kimura, T.; Miura, D.; Fukutomi, T.; Takeda, M.; Sakurai, H.; Endou, H. Functional Analysis of Human Organic Cation Transporter OCT3 (SLC22A3) Polymorphisms. J. Pharmacol. Sci. 2010, 113, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.S.; Tsai, H.-W.; Ko, H.-C.; Chang, F.-M.; Yeh, T.-L. Association of tryptophan hydroxylase gene polymorphism with depression, anxiety and comorbid depression and anxiety in a population-based sample of postpartum Taiwanese women. Genes, Brain Behav. 2004, 3, 328–336. [Google Scholar] [CrossRef]
- Duan, K.-M.; Wang, S.-Y.; Yin, J.-Y.; Li, X.; Ma, J.-H.; Huang, Z.-D.; Zhou, Y.-Y.; Yu, H.-Y.; Yang, M.; Zhou, H.-H.; et al. The IDO genetic polymorphisms and postpartum depressive symptoms: An association study in Chinese parturients who underwent cesarean section. Arch. Women’s Ment. Heal. 2018, 22, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Bortolato, M.; Shih, J.C. Behavioral outcomes of monoamine oxidase deficiency: Preclinical and clinical evidence. Int. Rev. Neurobiol. 2011, 100, 13–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achtyes, E.; Keaton, S.A.; Smart, L.; Burmeister, A.R.; Heilman, P.L.; Krzyzanowski, S.; Nagalla, M.; Guillemin, G.J.; Galvis, M.L.E.; Lim, C.K.; et al. Inflammation and kynurenine pathway dysregulation in post-partum women with severe and suicidal depression. Brain, Behav. Immun. 2020, 83, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Keaton, S.A.; Heilman, P.; Bryleva, E.Y.; Madaj, Z.; Krzyzanowski, S.; Grit, J.; Miller, E.S.; Jälmby, M.; Kalapotharakos, G.; Racicot, K.; et al. Altered Tryptophan Catabolism in Placentas From Women With Pre-eclampsia. Int. J. Tryptophan Res. 2019, 12, 1178646919840321. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Wallace, E.M.; Walker, D.W. Altered placental tryptophan metabolic pathway in human fetal growth restriction. Placenta 2017, 52, 62–70. [Google Scholar] [CrossRef]
- Lane, R.H. Fetal Programming, Epigenetics, and Adult Onset Disease. Clin. Perinatol. 2014, 41, 815–831. [Google Scholar] [CrossRef]
- Mastorci, F.; Agrimi, J. Fetal Programming of Adult Disease in a Translational Point of View; Springer Science and Business Media LLC: Cham, Switzerland, 2019; pp. 19–34. [Google Scholar]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nat. Cell Biol. 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Carrasco, G.; Cruz, M.A.; Dominguez, A.; Gallardo, V.; Miguel, P.; González, C. The expression and activity of monoamine oxidase A, but not of the serotonin transporter, is decreased in human placenta from pre-eclamptic pregnancies. Life Sci. 2000, 67, 2961–2969. [Google Scholar] [CrossRef]
- Takikawa, O.; Kuroiwa, T.; Yamazaki, F.; Kido, R. Mechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activity. J. Biol. Chem. 1988, 263, 2041–2048. [Google Scholar]
- Goeden, N.; Velasquez, J.; Arnold, K.A.; Chan, Y.; Lund, B.T.; Anderson, G.M.; Bonnin, A. Maternal Inflammation Disrupts Fetal Neurodevelopment via Increased Placental Output of Serotonin to the Fetal Brain. J. Neurosci. 2016, 36, 6041–6049. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | GD 12 | GD 15 | GD 18 | GD 21 |
|---|---|---|---|---|
| Number of fetuses (n) | NA | 11.33 ± 4.61 | 10.50 ± 2.89 | 8.25 ± 5.50 |
| Rat weight (g) | 333.00 ± 39.15 | 413.00 ± 54.27 | 358.00 ± 27.06 | 405.00 ± 5.00 |
| Placental weight (g) | 1.34 ± 0.25 1 | 1.70 ± 0.36 | 3.58 ± 0.72 | 5.76 ± 1.96 |
| Fetus weight (g) | 1.75 ± 0.61 | - | - | |
| TRP plasma (µg/mL) | 16.83 ± 2.75 | 19.58 ± 3.05 | 19.60 ± 1.64 | 13.16 ± 5.34 |
| 5-OH-TRP plasma (ng/mL) | 78.81 ± 12.60 | 68.82 ± 7.94 | 76.04 ± 12.98 | 100.82 ± 37.76 |
| 5-HT plasma (ng/mL) | 51.78 ± 41.10 | 154.98 ± 171.90 | 292.83 ± 314.57 | 215.59 ± 295.30 |
| KYN plasma (ng/mL) | 456.88 ± 213.10 | 407.82 ± 50.35 | 504.21 ± 98.76 | 429.93 ± 130.39 |
| KYNA plasma (ng/mL) | 155.60 ± 11.60 | 159.77 ± 53.80 | 147.20 ± 44.23 | 148.41 ± 26.38 |
| 5-OH-TRP/TRP ratio × 103 | 4.73 ± 0.75 | 3.62 ± 0.86 | 3.94 ± 0.93 | 9.25 ± 5.61 |
| KYN/TRP ratio × 103 | 26.55 ± 9.69 | 21.03 ± 2.76 | 26.12 ± 6.91 | 36.33 ± 15.75 |
| KYNA/KYN ratio × 103 | 395.90 ± 151.48 | 410.26 ± 202.86 | 308.64 ± 138.30 | 359.73 ± 78.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abad, C.; Karahoda, R.; Kastner, P.; Portillo, R.; Horackova, H.; Kucera, R.; Nachtigal, P.; Staud, F. Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit during Gestation. Int. J. Mol. Sci. 2020, 21, 7578. https://doi.org/10.3390/ijms21207578
Abad C, Karahoda R, Kastner P, Portillo R, Horackova H, Kucera R, Nachtigal P, Staud F. Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit during Gestation. International Journal of Molecular Sciences. 2020; 21(20):7578. https://doi.org/10.3390/ijms21207578
Chicago/Turabian StyleAbad, Cilia, Rona Karahoda, Petr Kastner, Ramon Portillo, Hana Horackova, Radim Kucera, Petr Nachtigal, and Frantisek Staud. 2020. "Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit during Gestation" International Journal of Molecular Sciences 21, no. 20: 7578. https://doi.org/10.3390/ijms21207578
APA StyleAbad, C., Karahoda, R., Kastner, P., Portillo, R., Horackova, H., Kucera, R., Nachtigal, P., & Staud, F. (2020). Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit during Gestation. International Journal of Molecular Sciences, 21(20), 7578. https://doi.org/10.3390/ijms21207578