Impact of Dim Light at Night on Urinary 6-Sulphatoxymelatonin Concentrations and Sleep in Healthy Humans



Abstract
:1. Introduction
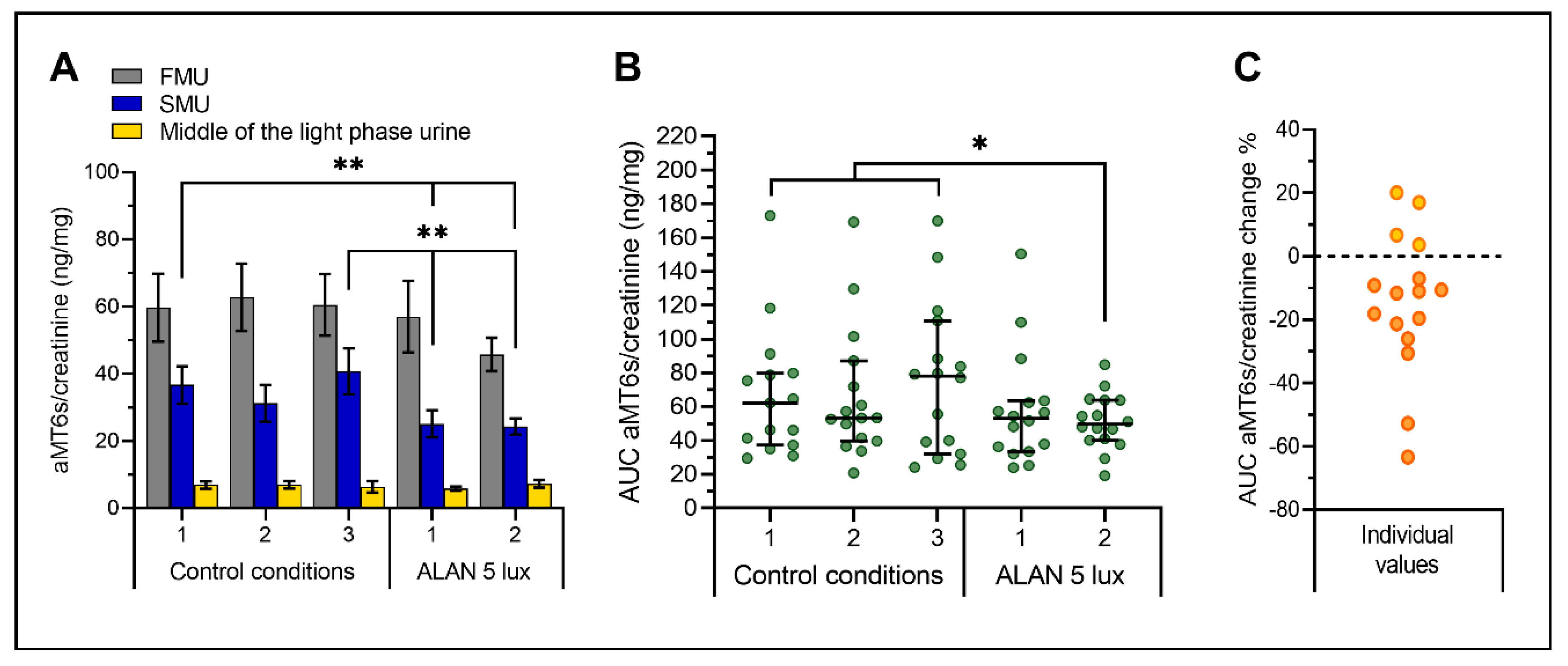
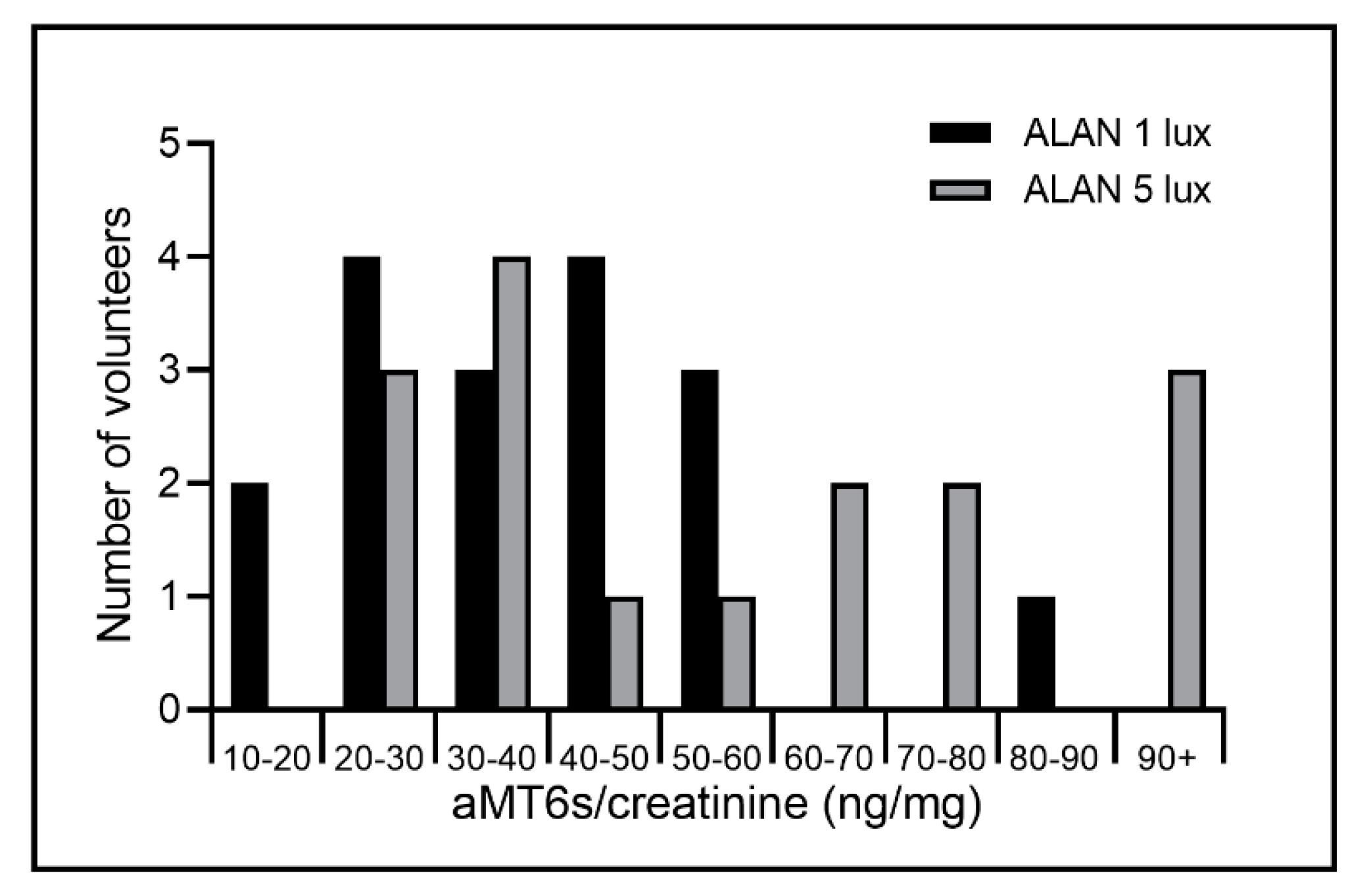
2. Results
2.1. ALAN to 1 Lux Exposure
2.2. ALAN up to 5 Lux Exposure
3. Discussion
4. Materials and Methods
4.1. Volunteers
4.2. Ethical Considerations
4.3. Study Design
4.3.1. Experiment 1—ALAN to 1 Lux Exposure
4.3.2. Experiment 2—ALAN up to 5 Lux Exposure
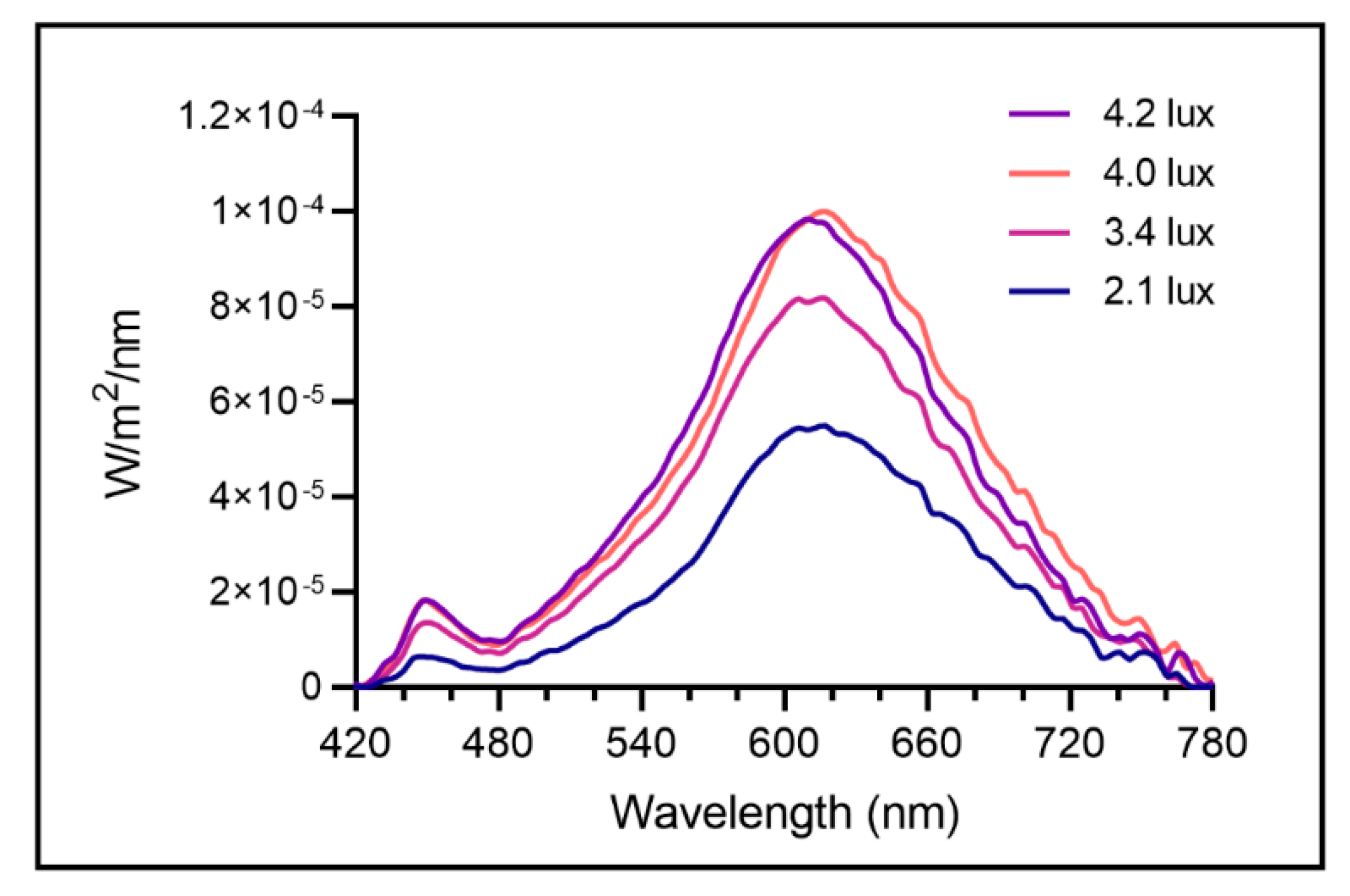
4.4. Lighting Conditions
4.5. Sleep Assessment
4.6. Melatonin Measurement
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FMU | First-morning urine |
| SMU | Second-morning urine |
| ALAN | Artificial light at night |
| SCN | Suprachiasmatic nucleus |
| ipRGCs | Intrinsically photosensitive retinal ganglion cells |
| VLPO | Ventrolateral preoptic area |
| M1 | Subtype of ipRGC |
| AUC | Area under the curve |
| CI | Confidence interval |
| AA-NAT | Arylalkylamine-N-acetyltransferase |
Appendix A


References
- Cho, Y.M.; Ryu, S.H.; Lee, B.R.; Kim, K.H.; Lee, E.; Choi, J. Effects of artificial light at night on human health: A literature review of observational and experimental studies applied to exposure assessment. Chronobiol. Int. 2015, 32, 1294–1310. [Google Scholar] [CrossRef]
- Hölker, F.; Moss, T.; Griefahn, B.; Kloas, W.; Voigt, C.C.; Henckel, D.; Hänel, A.; Kappeler, P.M.; Völker, S.; Schwope, A.; et al. The dark side of light: A transdisciplinary research agenda for light pollution policy. Ecol. Soc. 2010, 15. [Google Scholar] [CrossRef]
- Gaston, K.J.; Visser, M.E.; Hölker, F. The biological impacts of artificial light at night: The research challenge. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C.C.M.; Elvidge, C.D.; Baugh, K.; Portnov, B.A.; Rybnikova, N.A.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitschan, M.; Aguirre, G.K.; Brainard, D.H.; Sweeney, A.M. Variation of outdoor illumination as a function of solar elevation and light pollution. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Stevens, R.G.; Davis, S. The melatonin hypothesis: Electric power and breast cancer. Environ. Health Perspect. 1996, 104, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Arendt, J. Melatonin and the pineal gland: Influence on mammalian seasonal and circadian physiology. Rev. Reprod. 1998, 3, 13–22. [Google Scholar] [CrossRef]
- Arendt, J. Melatonin: Countering Chaotic Time Cues. Front. Endocrinol. (Lausanne) 2019, 10, 391. [Google Scholar] [CrossRef] [Green Version]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Von Gall, C.; Stehle, J.H.; Weaver, D.R. Mammalian melatonin receptors: Molecular biology and signal transduction. Cell Tissue Res. 2002, 309, 151–162. [Google Scholar] [CrossRef]
- Benloucif, S.; Burgess, H.J.; Klerman, E.B.; Lewy, A.J.; Middleton, B.; Murphy, P.J.; Parry, B.L.; Revell, V.L. Measuring melatonin in humans. J. Clin. Sleep Med. 2008, 4, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.; Cook, M.R.; Kavet, R.; Sastre, A.; Smith, D.K. Prediction of nocturnal plasma melatonin from morning urinary measures. J. Pineal Res. 1998, 24, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Bojkowski, C.J.; Arendt, J.; Shih, M.C.; Markey, S.P. Melatonin secretion in humans assessed by measuring its metabolite, 6-sulfatoxymelatonin. Clin. Chem. 1987, 33, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Grubisic, M.; Haim, A.; Bhusal, P.; Dominoni, D.M.; Gabriel, K.M.A.; Jechow, A.; Kupprat, F.; Lerner, A.; Marchant, P.; Riley, W.; et al. Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability 2019, 11, 6400. [Google Scholar] [CrossRef] [Green Version]
- Wever, R.A. Light Effects on Human Circadian Rhythms: A Review of Recent Andechs Experiments. J. Biol. Rhythms 1989, 4, 49–73. [Google Scholar] [CrossRef] [Green Version]
- Zeitzer, J.M.; Dijk, D.-J.; Kronauer, R.E.; Brown, E.N.; Czeisler, C.A. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef]
- Phillips, A.J.K.; Vidafar, P.; Burns, A.C.; McGlashan, E.M.; Anderson, C.; Rajaratnam, S.M.W.; Lockley, S.W.; Cain, S.W. High sensitivity and interindividual variability in the response of the human circadian system to evening light. Proc. Natl. Acad. Sci. USA 2019, 116, 12019–12024. [Google Scholar] [CrossRef] [Green Version]
- Obayashi, K.; Saeki, K.; Iwamoto, J.; Okamoto, N.; Tomioka, K.; Nezu, S.; Ikada, Y.; Kurumatani, N. Exposure to Light at Night, Nocturnal Urinary Melatonin Excretion, and Obesity/Dyslipidemia in the Elderly: A Cross-Sectional Analysis of the HEIJO-KYO Study. J. Clin. Endocrinol. Metab. 2013, 98, 337–344. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Kurumatani, N. Association between light exposure at night and insomnia in the general elderly population: The HEIJO-KYO cohort. Chronobiol. Int. 2014, 31, 976–982. [Google Scholar] [CrossRef]
- Esaki, Y.; Kitajima, T.; Obayashi, K.; Saeki, K.; Fujita, K.; Iwata, N. Light exposure at night and sleep quality in bipolar disorder: The APPLE cohort study. J. Affect. Disord. 2019, 257, 314–320. [Google Scholar] [CrossRef]
- Foster, R.G. Sleep, circadian rhythms and health. Interface Focus 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, R.G.; Brainard, G.C.; Blask, D.E.; Lockley, S.W.; Motta, M.E. Breast cancer and circadian disruption from electric lighting in the modern world. CA. Cancer J. Clin. 2014, 64, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Blask, D.E.; Dauchy, R.T.; Brainard, G.C.; Hanifin, J.P. Circadian stage-dependent inhibition of human breast cancer metabolism and growth by the nocturnal melatonin signal: Consequences of its disruption by light at night in rats and women. Integr. Cancer Ther. 2009, 8, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Zeman, M.; Herichova, I. Melatonin and clock genes expression in the cardiovascular system. Front. Biosci. Schol. 2013, 5 S, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Stein, R.M.; Kang, H.J.; McCorvy, J.D.; Glatfelter, G.C.; Jones, A.J.; Che, T.; Slocum, S.; Huang, X.P.; Savych, O.; Moroz, Y.S.; et al. Virtual discovery of melatonin receptor ligands to modulate circadian rhythms. Nature 2020, 579, 609–614. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Do Amaral, F.G. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [Green Version]
- Vartanian, G.V.; Li, B.Y.; Chervenak, A.P.; Walch, O.J.; Pack, W.; Ala-Laurila, P.; Wong, K.Y. Melatonin Suppression by Light in Humans Is More Sensitive Than Previously Reported. J. Biol. Rhythms 2015, 30, 351–354. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, D.C.; Fogerson, P.M.; Lazzerini Ospri, L.; Thomsen, M.B.; Layne, R.M.; Severin, D.; Zhan, J.; Singer, J.H.; Kirkwood, A.; Zhao, H.; et al. Light Affects Mood and Learning through Distinct Retina-Brain Pathways. Cell 2018, 175, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Gooley, J.J.; Lu, J.; Fischer, D.; Saper, C.B. A broad role for melanopsin in nonvisual photoreception. J. Neurosci. 2003, 23, 7093–7106. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Sonoda, T.; Schmidt, T.M. M1 Intrinsically Photosensitive Retinal Ganglion Cells Integrate Rod and Melanopsin Inputs to Signal in Low Light. Cell Rep. 2019, 29, 3349–3355. [Google Scholar] [CrossRef] [Green Version]
- Lax, P.; Ortuño-Lizarán, I.; Maneu, V.; Vidal-Sanz, M.; Cuenca, N. Photosensitive melanopsin-containing retinal ganglion cells in health and disease: Implications for circadian rhythms. Int. J. Mol. Sci. 2019, 20, 3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, C.H.; Lee, H.J.; Yoon, H.K.; Kang, S.G.; Bok, K.N.; Jung, K.Y.; Kim, L.; Lee, E. Il Exposure to dim artificial light at night increases REM sleep and awakenings in humans. Chronobiol. Int. 2016, 33, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Paksarian, D.; Rudolph, K.E.; Stapp, E.K.; Dunster, G.P.; He, J.; Mennitt, D.; Hattar, S.; Casey, J.A.; James, P.; Merikangas, K.R. Association of Outdoor Artificial Light at Night with Mental Disorders and Sleep Patterns among US Adolescents. JAMA Psychiatry 2020, 20892, 1–10. [Google Scholar] [CrossRef]
- Duffy, J.F.; Cain, S.W.; Chang, A.M.; Phillips, A.J.K.; Münch, M.Y.; Gronfier, C.; Wyatt, J.K.; Dijk, D.J.; Wright, K.P.; Czeisler, C.A. Sex difference in the near-24-hour intrinsic period of the human circadian timing system. Proc. Natl. Acad. Sci. USA 2011, 108, 15602–15608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hébert, M.; Martin, S.K.; Lee, C.; Eastman, C.I. The effects of prior light history on the suppression of melatonin by light in humans. J. Pineal Res. 2002, 33, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Nathan, P.J.; Wyndham, E.L.; Burrows, G.D.; Norman, T.R. The effect of gender on the melatonin suppression by light: A dose response relationship. J. Neural Transm. 2000, 107, 271–279. [Google Scholar] [CrossRef]
- Smolensky, M.H.; Sackett-Lundeen, L.L.; Portaluppi, F. Nocturnal light pollution and underexposure to daytime sunlight: Complementary mechanisms of circadian disruption and related diseases. Chronobiol. Int. 2015, 32, 1029–1048. [Google Scholar] [CrossRef]
- Chaves, I.; van der Eerden, B.; Boers, R.; Boers, J.; Streng, A.A.; Ridwan, Y.; Schreuders-Koedam, M.; Vermeulen, M.; van der Pluijm, I.; Essers, J.; et al. Gestational jet lag predisposes to later-life skeletal and cardiac disease. Chronobiol. Int. 2019, 36, 657–671. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Light and circadian signaling pathway in pregnancy: Programming of adult health and disease. Int. J. Mol. Sci. 2020, 21, 2232. [Google Scholar] [CrossRef] [Green Version]
- Galdames, H.A.; Torres-Farfan, C.; Spichiger, C.; Mendez, N.; Abarzua-Catalan, L.; Alonso-Vazquez, P.; Richter, H.G. Impact of gestational chronodisruption on fetal cardiac genomics. J. Mol. Cell. Cardiol. 2014, 66, 1–11. [Google Scholar] [CrossRef]
- Parveen, A.; Akash, M.S.H.; Rehman, K.; Kyunn, W.W. Dual role of p21 in the progression of cancer and its treatment. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Dominoni, D.M.; Nelson, R.J. Artificial light at night as an environmental pollutant: An integrative approach across taxa, biological functions, and scientific disciplines. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2018, 329, 387–393. [Google Scholar] [CrossRef]
- Folkard, S.; Monk, T.H.; Lobuan, M.C. Towards a Predictive Test of Adjustment to Shift Work. Ergonomics 1979, 22, 79–91. [Google Scholar] [CrossRef]
- Stebelová, K.; Molčan, Ľ.; Okuliarová, M.; Hanuliak, P.; Hartman, P.; Hraška, J.; Zeman, M. The influence of indoor lighting with low blue light dose on urine 6-sulphatoxymelatonin concentrations and sleep efficiency of healthy volunteers. Biol. Rhythm Res. 2015, 46, 137–145. [Google Scholar] [CrossRef]
- Kushida, C.A.; Chang, A.; Gadkary, C.; Guilleminault, C.; Carrillo, O.; Dement, W.C. Comparison of actigraphic, polysomnographic, and subjective assessment of sleep parameters in sleep-disordered patients. Sleep Med. 2001, 2, 389–396. [Google Scholar] [CrossRef]
- Littner, M.; Kushida, C.A.; Anderson, W.M.D.; Bailey, D.; Berry, R.B.; Davila, D.G.; Hirshkowitz, M.; Kapen, S.; Kramer, M.; Loube, D.; et al. Practice parameters for the role of actigraphy in the study of sleep and circadian rhythms: An update for 2002. Sleep 2003, 26, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Aldhous, M.E.; Arendt, J. Radioimmunoassay for 6-Sulphatoxymelatonin in Urine Using an Iodinated Tracer. Ann. Clin. Biochem. 1988, 25, 298–303. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Conditions | Samples | Mean aMT6s ng/mg Creatinine n = 17 Each | SEM | Minimum Value ng/mg | Maximum Value ng/mg |
|---|---|---|---|---|---|
| Control (3 nights) | First morning urine 1 | 39.75 | ±2.69 | 13.30 | 97.16 |
| Middle of the light phase urine 2 | 5.69 | ±0.73 | 0.98 | 29.37 | |
| ALAN 1 lux (4 nights) | First morning urine 1 | 42.22 | ±2.50 | 10.69 | 107.69 |
| Middle of the light phase urine 2 | 5.52 | ±0.72 | 0.74 | 43.93 |
| Control Nights Mean ± SEM | ALAN 1 Lux Mean ± SEM | p1 | |
|---|---|---|---|
| Time in bed | 7:43 ± 0:14 | 8:07 ± 0:11 | 0.080 |
| Assumed sleep | 7:23 ± 0:14 | 7:45 ± 0:10 | 0.086 |
| Actual sleep time | 6:21 ± 0:12 | 6:39 ± 0:09 | 0.069 |
| Actual sleep (%) | 86.09 ± 1.14 | 85.92 ± 1.00 | 0.737 |
| Actual wake time | 1:02 ± 0:05 | 1:06 ± 0:05 | 0.269 |
| Actual wake (%) | 13.91 ± 1.14 | 14.08 ± 1.00 | 0.735 |
| Sleep efficiency | 82.38 ± 1.12 | 82.11 ± 1.18 | 0.752 |
| Sleep latency | 0:15 ± 0:02 | 0:16 ± 0:02 | 0.655 |
| Sleep bouts | 27.25 ± 1.75 | 28.89 ± 1.42 | 0.159 |
| Wake bouts | 27.27 ± 1.73 | 28.91 ± 1.41 | 0.156 |
| Mean sleep bout time | 0:15 ± 0:01 | 0:14 ± 0:00 | 0.312 |
| Mean wake bout time | 0:02 ± 0:00 | 0:02 ± 0:00 | 0.822 |
| Immobile mins | 375 ± 12 | 391 ± 10 | 0.117 |
| Immobile time (%) | 84.71 ± 1.30 | 84.06 ± 1.31 | 0.129 |
| Moving mins | 68.09 ± 6.47 | 74.45 ± 6.23 | 0.080 |
| Moving time (%) | 15.29 ± 1.30 | 15.94 ± 1.31 | 0.129 |
| No of immobile phases | 42.26 ± 2.71 | 45.32 ± 2.48 | 0.077 |
| Mean length immobility | 9.57 ± 0.7 | 9.29 ± 0.65 | 0.324 |
| One Minute immobility | 7.50 ± 0.91 | 8.84 ± 1.03 | 0.003 ** |
| One Min immobility (%) | 16.84 ± 1.33 | 18.71 ± 1.44 | 0.073 |
| Total activity score | 7480 ± 838 | 8295 ± 950 | 0.233 |
| Mean activity score | 16.77 ± 1.79 | 17.71 ± 2.00 | 0.355 |
| Mean score in active periods | 112 ± 9 | 112 ± 9 | 0.974 |
| Fragmentation index | 32.13 ± 2.49 | 34.66 ± 2.68 | 0.042 * |
| Control Conditions Mean ± SEM | ALAN 5 Lux Mean ± SEM | p1 | |
|---|---|---|---|
| Time in bed | 7:49 ± 0:09 | 8:08 ± 0:08 | 0.083 |
| Assumed sleep | 7:42 ± 0:09 | 8:01 ± 0:08 | 0.088 |
| Actual sleep time | 6:44 ± 0:09 | 7:00 ± 0:09 | 0.172 |
| Actual sleep (%) | 87.10 ± 1.10 | 86.83 ± 1.47 | 0.771 |
| Actual wake time | 0:57 ± 0:05 | 1:02 ± 0:07 | 0.341 |
| Actual wake (%) | 11.94 ± 1.10 | 12.33 ± 1.45 | 0.671 |
| Sleep efficiency | 85.87 ± 1.16 | 85.43 ± 1.43 | 0.654 |
| Sleep latency | 0:05 ± 0:01 | 0:05 ± 0:01 | 0.496 |
| Sleep bouts | 26.84 ± 1.83 | 28.07 ± 2.60 | 0.405 |
| Wake bouts | 26.87 ± 1.81 | 27.93 ± 2.66 | 0.497 |
| Mean sleep bout time | 0:16 ± 0:01 | 0:17 ± 0:02 | 0.586 |
| Mean wake bout time | 0:02 ± 0:00 | 0:02 ± 0:00 | 0.525 |
| Immobile mins | 410 ± 9 | 427 ± 10 | 0.133 |
| Immobile time (%) | 88.41 ± 0.83 | 88.23 ± 1.21 | 0.812 |
| Moving mins | 51.60 ± 3.86 | 54.40 ± 6.03 | 0.469 |
| Moving time (%) | 10.64 ± 0.85 | 10.80 ± 1.22 | 0.836 |
| No of immobile phases | 38.03 ± 2.55 | 39.33 ± 3.56 | 0.584 |
| Mean length immobility | 11.36 ± 0.88 | 11.93 ± 1.21 | 0.499 |
| One Minute immobility | 4.33 ± 0.63 | 5.77 ± 1.04 | 0.074 |
| One Min immobility (%) | 10.09 ± 0.99 | 12.63 ± 1.42 | 0.037 * |
| Total activity score | 8035 ± 1444 | 7657 ± 875 | 0.792 |
| Mean activity score | 16.72 ± 3.05 | 15.33 ± 1.76 | 0.645 |
| Mean score in active periods | 159 ± 32 | 145 ± 13 | 0.659 |
| Fragmentation index | 21.23 ± 1.77 | 24.00 ± 2.52 | 0.123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stebelova, K.; Roska, J.; Zeman, M. Impact of Dim Light at Night on Urinary 6-Sulphatoxymelatonin Concentrations and Sleep in Healthy Humans. Int. J. Mol. Sci. 2020, 21, 7736. https://doi.org/10.3390/ijms21207736
Stebelova K, Roska J, Zeman M. Impact of Dim Light at Night on Urinary 6-Sulphatoxymelatonin Concentrations and Sleep in Healthy Humans. International Journal of Molecular Sciences. 2020; 21(20):7736. https://doi.org/10.3390/ijms21207736
Chicago/Turabian StyleStebelova, Katarina, Jan Roska, and Michal Zeman. 2020. "Impact of Dim Light at Night on Urinary 6-Sulphatoxymelatonin Concentrations and Sleep in Healthy Humans" International Journal of Molecular Sciences 21, no. 20: 7736. https://doi.org/10.3390/ijms21207736
APA StyleStebelova, K., Roska, J., & Zeman, M. (2020). Impact of Dim Light at Night on Urinary 6-Sulphatoxymelatonin Concentrations and Sleep in Healthy Humans. International Journal of Molecular Sciences, 21(20), 7736. https://doi.org/10.3390/ijms21207736