Irisin Gene Delivery Ameliorates Burn-Induced Sensory and Motor Neuropathy

 , , and
, , and 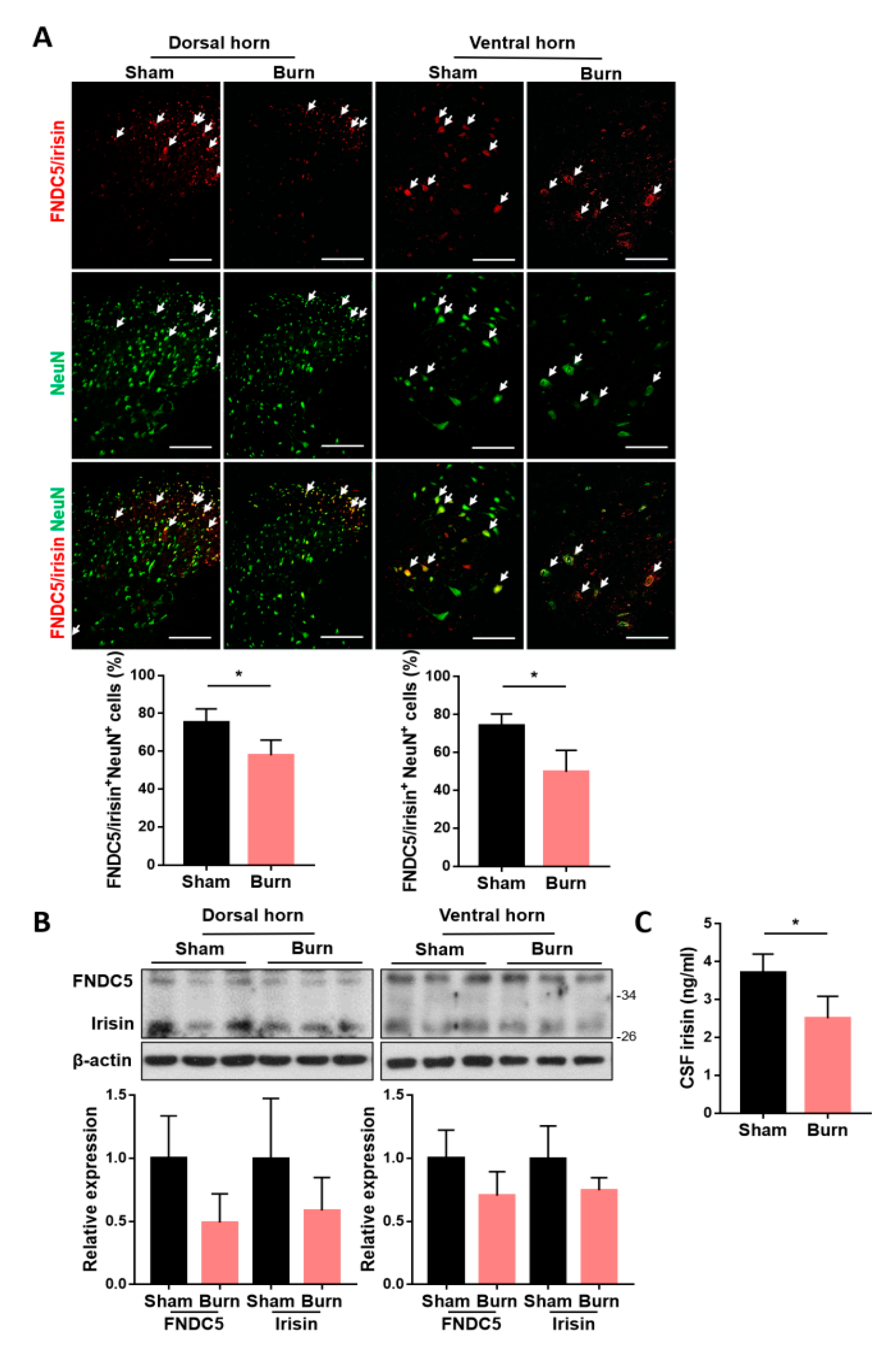{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expressions of Spinal FNDC5/Irisin and CSF Irisin Were Decreased after Burn Injury
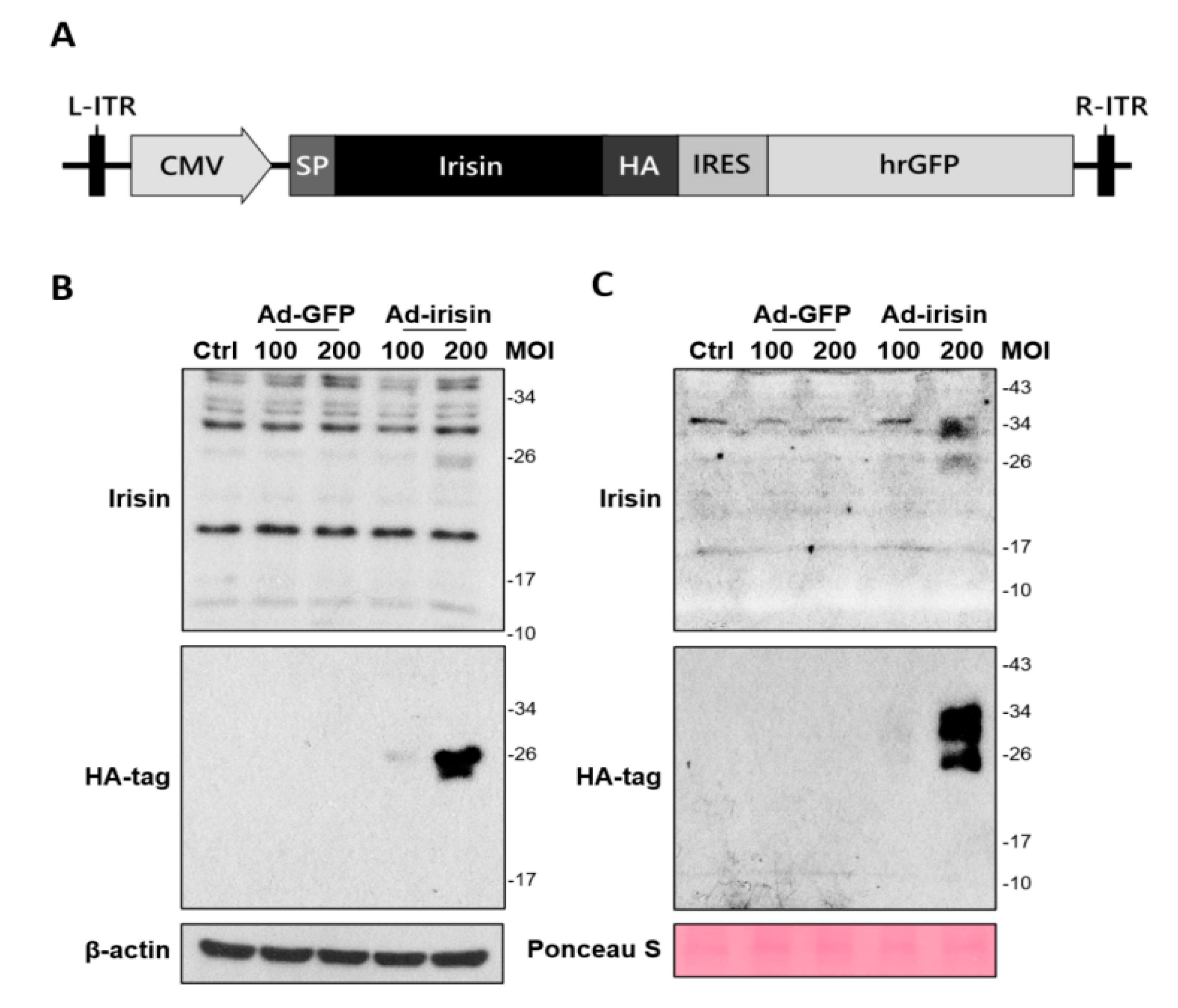
2.2. Adenoviral Vector Design for Irisin Gene Delivery and Validation of Recombinant Irisin Secretion
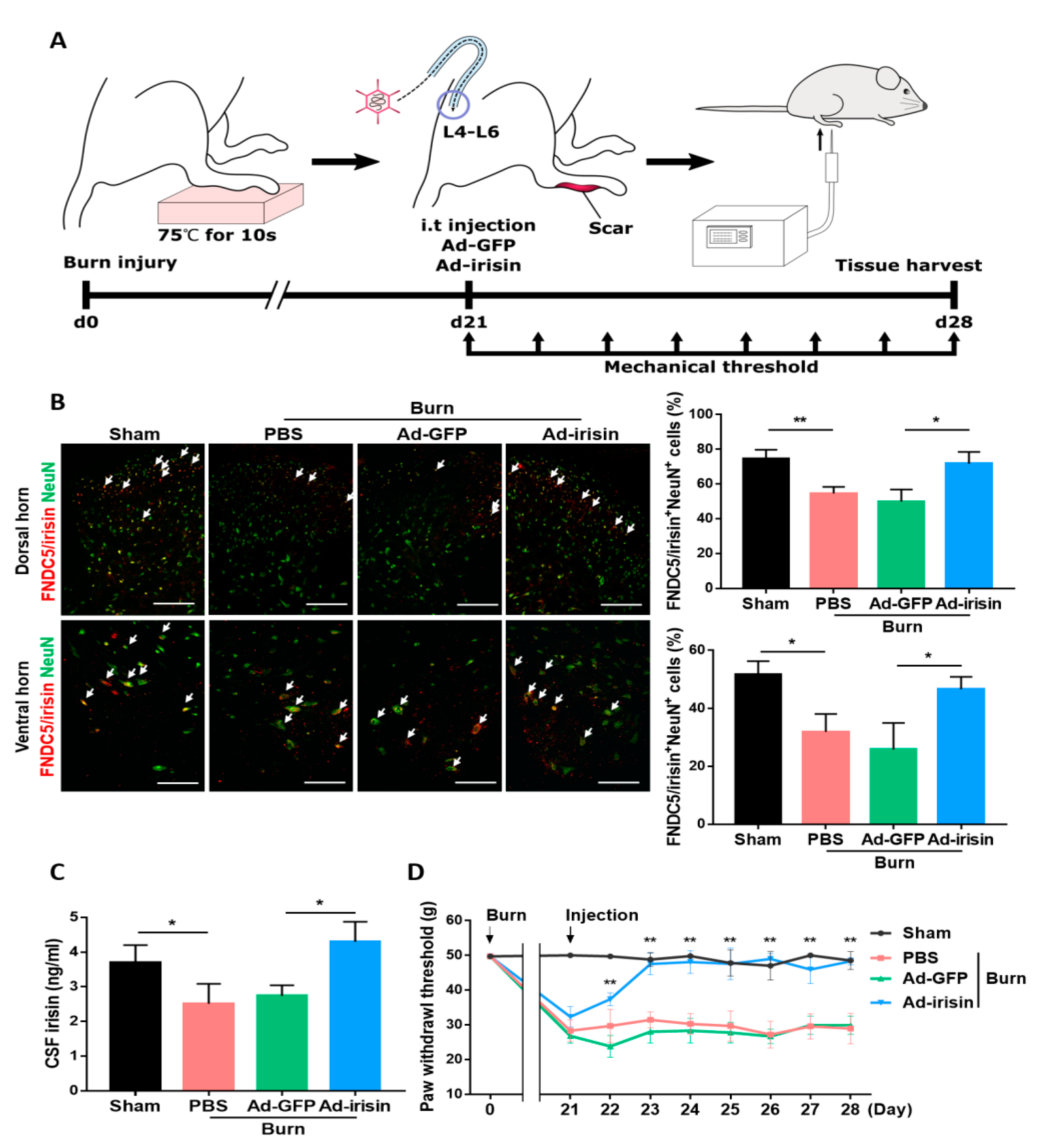
2.3. Intrathecal Irisin Gene Delivery Alleviated Postburn Allodynia
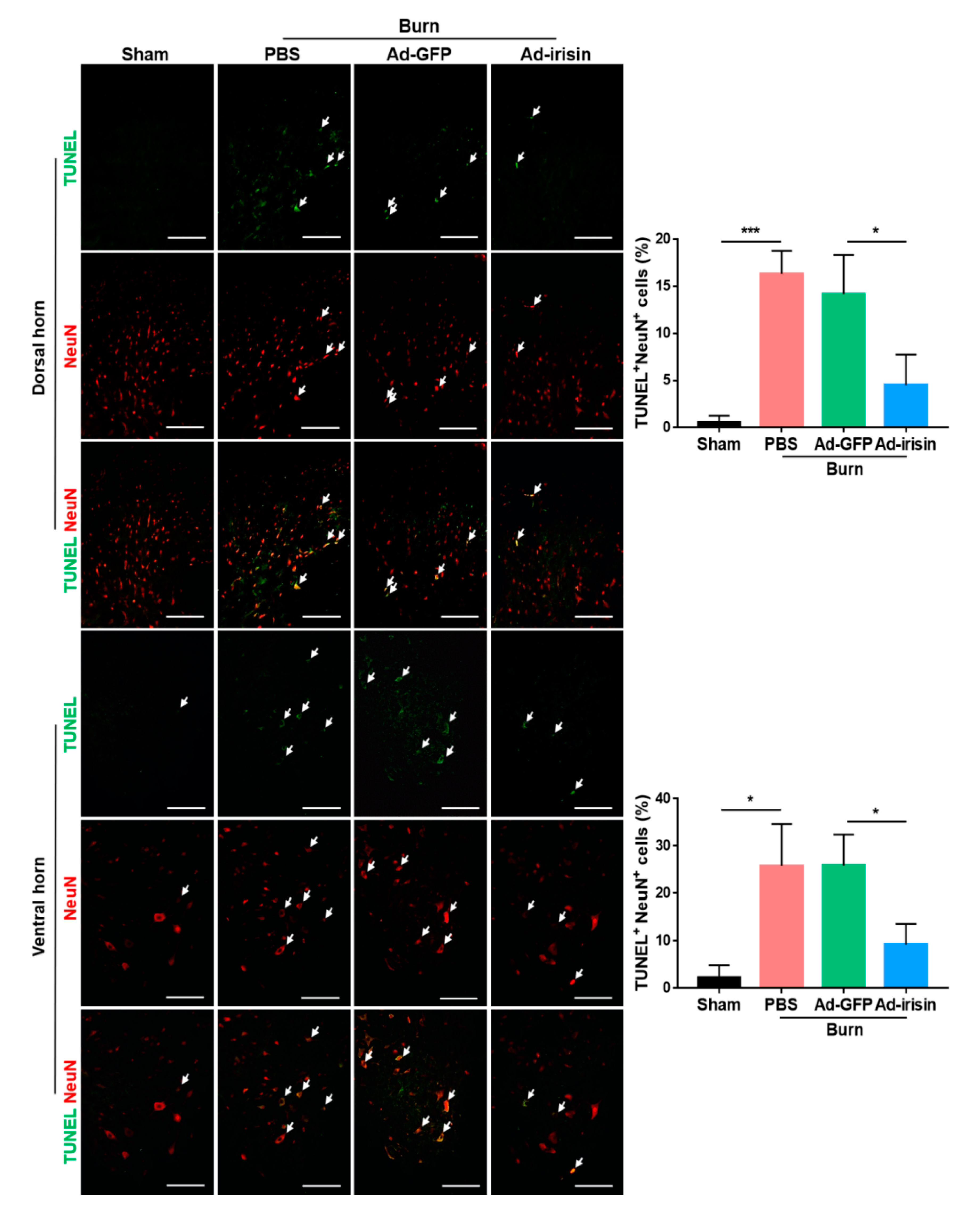
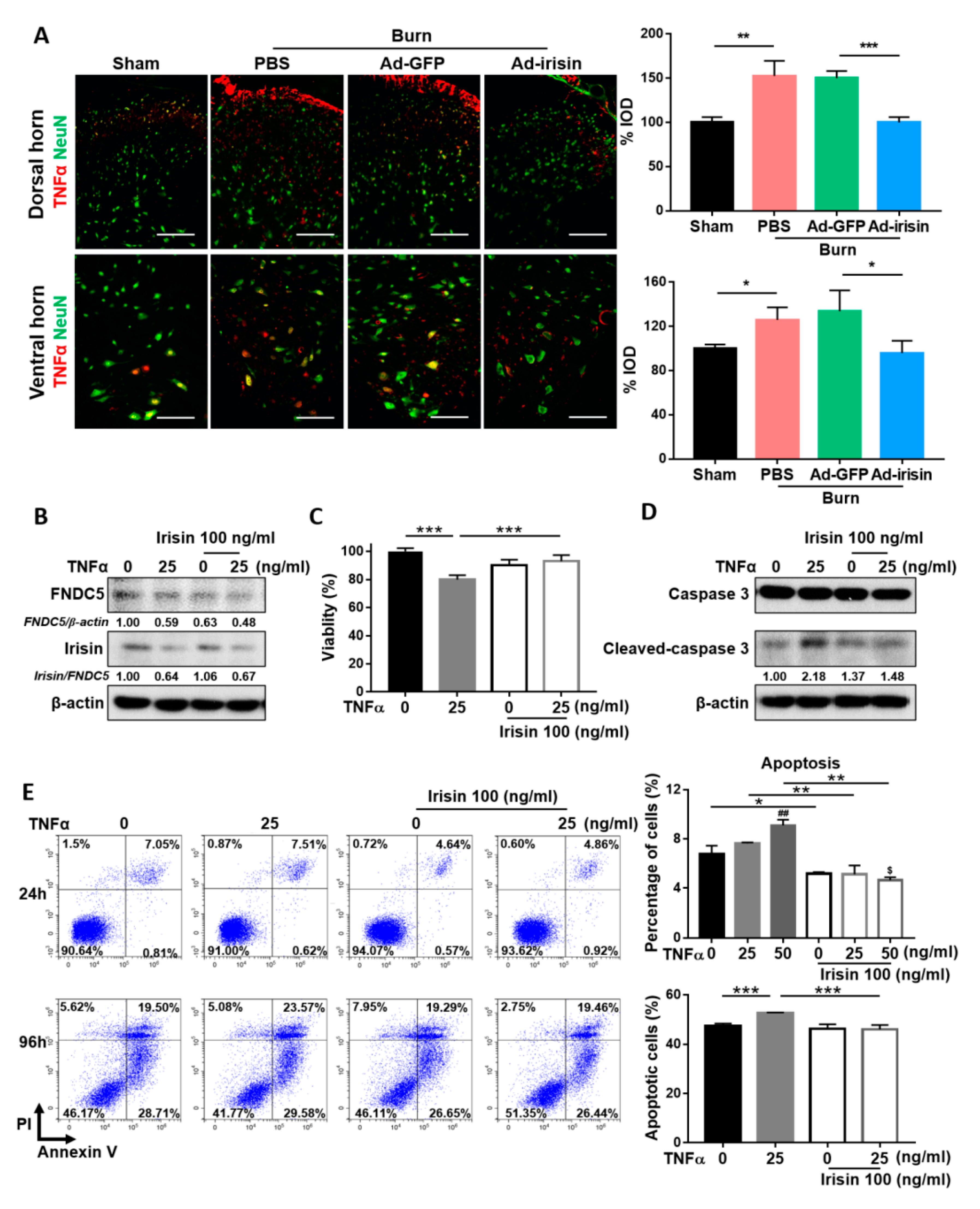
2.4. Irisin Gene Delivery Alleviated Neuronal Apoptosis in the Spinal Cord Postburn
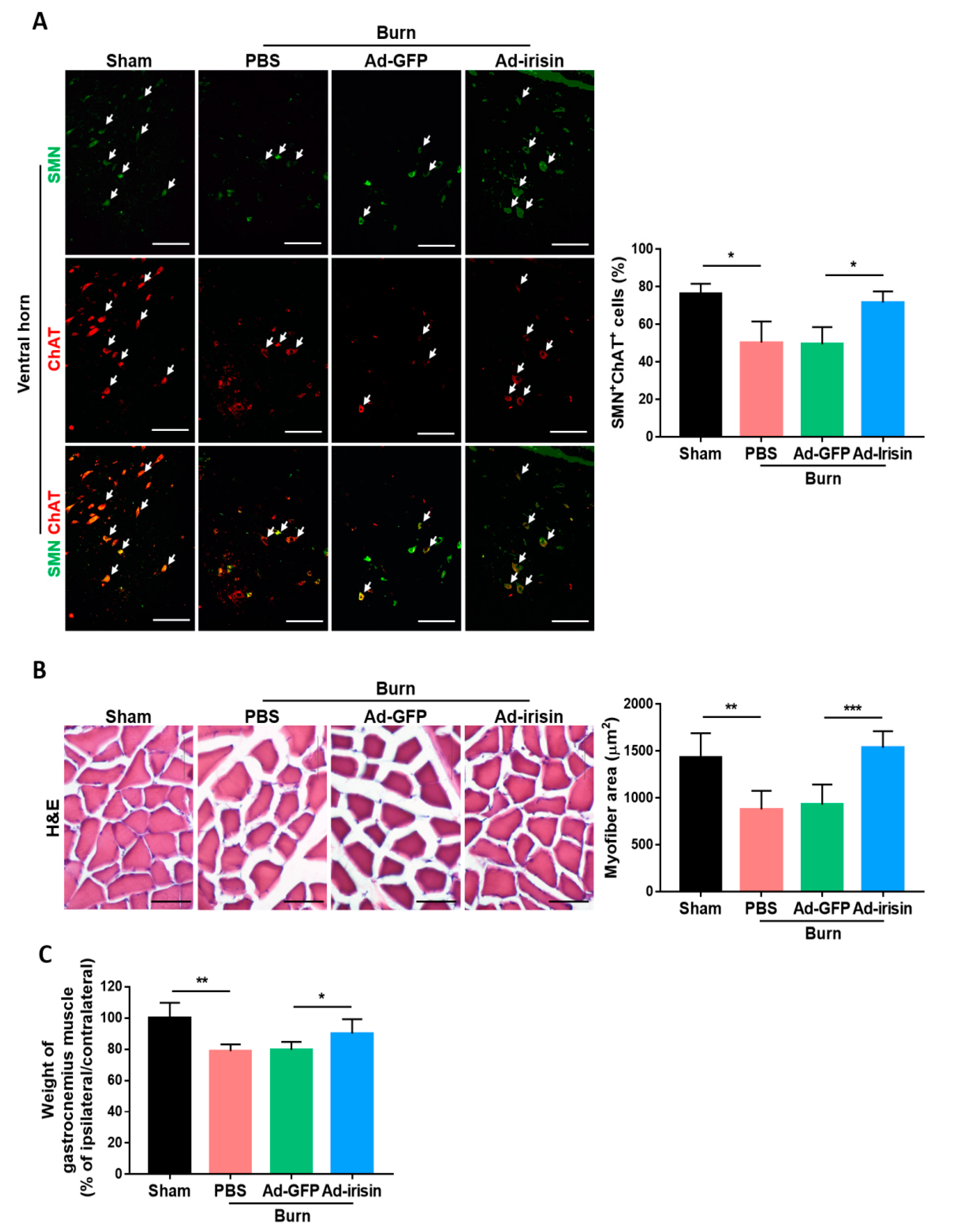
2.5. Irisin Improved the Survival of Motor Neuron Cells and Muscle Size Following Burn Injury
2.6. Irisin Attenuated TNFα-Induced Neuronal Damage
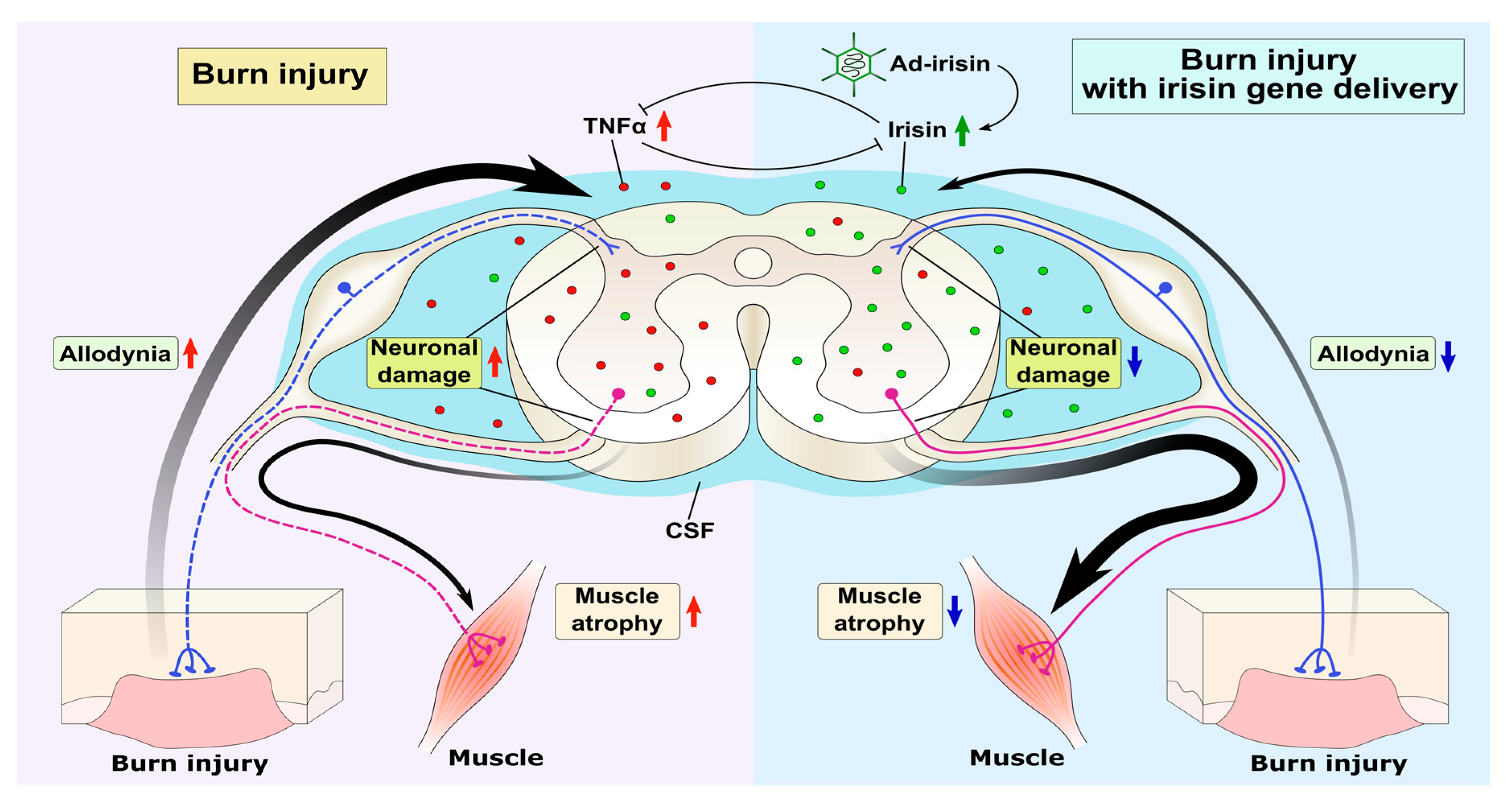
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Adenovirus
4.4. Recombinant Proteins
4.5. Burn Injury Model and Mechanical Allodynia Test
4.6. Intrathecal Injections
4.7. Immunofluorescence and H&E Staining
4.8. Immunoblotting Analysis and Irisin ELISA
4.9. Flow Cytometry Analysis
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FDNC5 | Fibronectin Type III Domain Containing 5 |
| TNFα | Tumor necrosis factor alpha |
| CSF | Cerebrospinal fluid |
| NeuN | Neuronal nuclei |
| SMN | Survival motor neuron |
| ChAT | Choline acetyltransferase |
| IL-6 | Interleukin 6 |
| IL-10 | Interleukin 10 |
| IL-1β | Interleukin 1 beta |
| NF-κB | Nuclear Factor Kappa B |
| Iba1 | Ionized calcium-binding adaptor molecule 1 |
| MPO-1 | Myeloperoxidase-1 |
| MCP1 | Monocyte chemoattractant protein-1 |
| CXCL2 | C-X-C Motif Chemokine Ligand 2 |
| GFAP | Glial fibrillary acidic protein |
| BDNF | Brain-derived neurotrophic factor |
| GDNF | Glial cell-derived neurotrophic factor |
| COX-2 | Cyclooxygenase-2 |
| PGE2 | Prostaglandin E2 |
| PGI2 | Prostaglandin I2 |
| GABA | γ-Aminobutyric acid |
| MOI | Multiplicity of infection |
| HA | Hemagglutinin |
| GFP | Green fluorescent protein |
References
- Marquez, S.; Turley, J.J.E.; Peters, W.J. Neuropathy in burn patients. Brain 1993, 116, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Vetrichevvel, T.P.; Randall, S.M.; Fear, M.W.; Wood, F.M.; Boyd, J.H.; Duke, J.M. Burn injury and long-term nervous system morbidity: A population-based cohort study. BMJ Open 2016, 6, e012668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamam, Y.; Tamam, C.; Tamam, B.; Ustundag, M.; Orak, M.; Tasdemir, N. Peripheral neuropathy after burn injury. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 17. [Google Scholar]
- Klein, M.B.; Hollingworth, W.; Rivara, F.P.; Kramer, C.B.; Askay, S.W.; Heimbach, D.M.; Gibran, N.S. Hospital Costs Associated With Pediatric Burn Injury. J. Burn. Care Res. 2008, 29, 632–637. [Google Scholar] [CrossRef]
- Malenfant, A.; Forget, R.; Amsel, R.; Papillon, J.; Frigon, J.-Y.; Choinière, M. Tactile, thermal and pain sensibility in burned patients with and without chronic pain and paresthesia problems. Pain 1998, 77, 241–251. [Google Scholar] [CrossRef]
- Bijlard, E. A Systematic Review on the Prevalence, Etiology, and Pathophysiology of Intrinsic Pain in Dermal Scar Tissue. Pain Physician 2017, 1, 1–13. [Google Scholar] [CrossRef]
- Strong, A.L.; Agarwal, S.; Cederna, P.S.; Levi, B. Peripheral Neuropathy and Nerve Compression Syndromes in Burns. Clin. Plast. Surg. 2017, 44, 793–803. [Google Scholar] [CrossRef]
- Alfonso-Sanchez, J.L.; Pereperez, S.B.; Bastida, J.; Martínez, M. Cost-Utility Analysis Applied to the Treatment of Burn Patients in a Specialized Center. Arch. Surg. 2007, 142, 50. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Jia, H.; Dai, H.; Li, W.; Zhang, L. Protective effects of pentoxifylline on the brain following remote burn injury. Burns 2010, 36, 1300–1308. [Google Scholar] [CrossRef]
- Morgan, M.; Deuis, J.R.; Froesig-Joergensen, M.; Lewis, R.J.; Cabot, P.J.; Gray, P.; Vetter, I. Burn Pain: A Systematic and Critical Review of Epidemiology, Pathophysiology, and Treatment. Pain Med. 2017, 19, 708–734. [Google Scholar] [CrossRef]
- Evers, L.H.; Bhavsar, D.; Mailänder, P. The biology of burn injury. Exp. Dermatol. 2010, 19, 777–783. [Google Scholar] [CrossRef]
- Lin, C.-H.; Wu, S.-H.; Lee, S.-S.; Lin, Y.-N.; Kuo, Y.-R.; Chai, C.-Y.; Huang, S.-H. Autologous Adipose-Derived Stem Cells Reduce Burn-Induced Neuropathic Pain in a Rat Model. Int. J. Mol. Sci. 2017, 19, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, J.C.; Harris, N.L.; El Shami, A.; Sheridan, R.L.; Schulz, J.T.; Bilodeau, M.-L.; Ryan, C.M. A Descriptive Review of Neuropathic-Like Pain After Burn Injury. J. Burn. Care Res. 2006, 27, 524–528. [Google Scholar] [CrossRef]
- Cavalli, E.; Mammana, S.; Nicoletti, F.; Bramanti, P.; Mazzon, E. The neuropathic pain: An overview of the current treatment and future therapeutic approaches. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419838383. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Calvert, C.T.; Cairns, B.A.; Hultman, C.S. Lower Extremity Nerve Decompression in Burn Patients. Ann. Plast. Surg. 2013, 70, 563–567. [Google Scholar] [CrossRef]
- Higashimori, H.; Whetzel, T.P.; Mahmood, T.; Carlsen, R.C. Peripheral axon caliber and conduction velocity are decreased after burn injury in mice. Muscle Nerve 2005, 31, 610–620. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; Huang, S.-H.; Cheng, K.-I.; Chai, C.-Y.; Yeh, J.-L.; Wu, T.-C.; Hsu, Y.-C.; Kwan, A.-L. Third-Degree Hindpaw Burn Injury Induced Apoptosis of Lumbar Spinal Cord Ventral Horn Motor Neurons and Sciatic Nerve and Muscle Atrophy in Rats. BioMed Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Ma, L.; Zhou, Y.; Khan, M.A.S.; Yasuhara, S.; Martyn, J.A.J. Burn-Induced Microglia Activation is Associated With Motor Neuron Degeneration and Muscle Wasting in Mice. Shock 2019, 51, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Bostroem, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostroem, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nat. Cell Biol. 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Arhire, L.I.; Mihalache, L.; Covasa, M. Irisin: A Hope in Understanding and Managing Obesity and Metabolic Syndrome. Front. Endocrinol. 2019, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Perakakis, N.; Triantafyllou, G.A.; Fernández-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and role of irisin in glucose homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Gizaw, M.; Anandakumar, P.; Debela, T. A Review on the Role of Irisin in Insulin Resistance and Type 2 Diabetes Mellitus. J. Pharmacopunct. 2017, 20, 235–242. [Google Scholar]
- Islam, M.R.; Young, M.F.; Wrann, C.D. The Role of FNDC5/Irisin in the Nervous System and as a Mediator for Beneficial Effects of Exercise on the Brain. In Hormones, Metabolism and the Benefits of Exercise; Spiegelman, B., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 93–102. [Google Scholar]
- Mazur-Bialy, A.I.; Pochec, E.; Zarawski, M. Anti-Inflammatory Properties of Irisin, Mediator of Physical Activity, Are Connected with TLR4/MyD88 Signaling Pathway Activation. Int. J. Mol. Sci. 2017, 18, 701. [Google Scholar] [CrossRef]
- Li, D.-J.; Li, Y.-H.; Yuan, H.-B.; Qu, L.-F.; Wang, P. The novel exercise-induced hormone irisin protects against neuronal injury via activation of the Akt and ERK1/2 signaling pathways and contributes to the neuroprotection of physical exercise in cerebral ischemia. In Metabolism: Clinical and experimental. Metabolism 2017, 68, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, H.; Wang, H.; Wang, J.-H.; Song, F.; Sun, Y. Irisin Exerts Neuroprotective Effects on Cultured Neurons by Regulating Astrocytes. Mediat. Inflamm. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Guo, P.; Li, X.; Ke, J.; Wang, Y.; Wu, H. Neuroprotective effects of irisin against cerebral ischemia/reperfusion injury via Notch signaling pathway. Biomed. Pharmacother. 2019, 120, 109452. [Google Scholar] [CrossRef]
- Ellis, A.; Bennett, D.L.H. Neuroinflammation and the generation of neuropathic pain. Br. J. Anaesth. 2013, 111, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Skaper, S.D.; Facci, L.; Zusso, M.; Giusti, P. Neuroinflammation, Mast Cells, and Glia: Dangerous Liaisons. Neuroscientist 2017, 23, 478–498. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Li, Y.-T.; Li, Z.-Y.; Hung, C.-H. Exercise Training Attenuates Neuropathic Pain and Cytokine Expression After Chronic Constriction Injury of Rat Sciatic Nerve. Anesth. Analg. 2012, 114, 1330–1337. [Google Scholar] [CrossRef]
- Dobson, J.L.; McMillan, J.; Li, L. Benefits of exercise intervention in reducing neuropathic pain. Front. Cell. Neurosci. 2014, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.M. Exercise-Induced Modulation of Neuroinflammation in Models of Alzheimer’s Disease. Brain Plast. 2018, 4, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Ignácio, Z.M.; Da Silva, R.S.; Plissari, M.E.; Quevedo, J.; Réus, G.Z. Physical Exercise and Neuroinflammation in Major Depressive Disorder. Mol. Neurobiol. 2019, 56, 8323–8335. [Google Scholar] [CrossRef] [PubMed]
- Dameni, S.; Janzadeh, A.; Yousefifard, M.; Nasirinezhad, F. The effect of intrathecal injection of irisin on pain threshold and expression rate of GABAB receptors in peripheral neuropathic pain model. J. Chem. Neuroanat. 2018, 91, 17–26. [Google Scholar] [CrossRef]
- Farzad, B.; Rajabi, H.; Gharakhanlou, R.; Allison, D.J.; Hayat, P.; Jameie, S.B. Swimming Training Attenuates Allodynia and Hyperalgesia Induced by Peripheral Nerve Injury in an Adult Male Rat Neuropathic Model: Effects on Irisin and GAD65. Pain Med. 2018, 19, 2236–2245. [Google Scholar] [CrossRef] [Green Version]
- Saraç, F.; Sarsu, S.B.; Yeniocak, S.; Şahin, K.; Yucetas, E.; Yildirim, D.; Koldas, M.; Uzun, O. The Diagnostic Value of Irisin in Pediatric Patients with Acute Abdominal Pain. Emerg. Med. Int. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Orellana, C.; Calvet, J.; Navarro, N.; Galisteo, C.; Gratacós, J.; Larrosa, M. AB0962 Irisin levels are associated with exercise, pain and function in patients with knee osteoarthritis. Osteoarthritis 2018, 77, 1605. [Google Scholar] [CrossRef]
- Huang, S.-H.; Wu, S.-H.; Lee, S.-S.; Chang, K.-P.; Chai, C.-Y.; Yeh, J.-L.; Lin, S.-D.; Kwan, A.-L.; Wang, H.-M.D.; Lai, C.-S. Fat Grafting in Burn Scar Alleviates Neuropathic Pain via Anti-Inflammation Effect in Scar and Spinal Cord. PLoS ONE 2015, 10, e0137563. [Google Scholar] [CrossRef] [Green Version]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Simon, C.M.; Janas, A.M.; Lotti, F.; Tapia, J.C.; Pellizzoni, L.; Mentis, G.Z. A Stem Cell Model of the Motor Circuit Uncouples Motor Neuron Death from Hyperexcitability Induced by SMN Deficiency. Cell Rep. 2016, 16, 1416–1430. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Simard, L.R. Survival motor neuron (SMN) protein: Role in neurite outgrowth and neuromuscular maturation during neuronal differentiation and development. Hum. Mol. Genet. 2002, 11, 1605–1614. [Google Scholar] [CrossRef] [Green Version]
- Chaytow, H.; Huang, Y.-T.; Gillingwater, T.H.; Faller, K.M.E. The role of survival motor neuron protein (SMN) in protein homeostasis. Cell. Mol. Life Sci. 2018, 75, 3877–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laird, A.S.; Mackovski, N.; Rinkwitz, S.; Becker, T.S.; Giacomotto, J. Tissue-specific models of spinal muscular atrophy confirm a critical role of SMN in motor neurons from embryonic to adult stages. Hum. Mol. Genet. 2016, 25, 1728–1738. [Google Scholar] [CrossRef] [PubMed]
- Soler-Botija, C.; Cuscó, I.; Lopez, E.; Clua, A.; Saladich, I.G.; Baiget, M.; Ferrer, I.; Tizzano, E.F. Choline acetyltransferase expression does not identify early pathogenic events in fetal SMA spinal cord. Neuromuscul. Disord. 2005, 15, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.; Cahill, C.M. TNF-alpha and neuropathic pain—A review. J. Neuroinflamm. 2010, 7, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merienne, N.; Le Douce, J.; Faivre, E.; Déglon, N.; Bonvento, G. Efficient gene delivery and selective transduction of astrocytes in the mammalian brain using viral vectors. Front. Cell. Neurosci. 2013, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Liu, D. N-Glycosylation is required for FDNC5 stabilization and irisin secretion. Biochem. J. 2017, 474, 3167–3177. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Tan, A.; Saab, C.; Waxman, S.G. Unilateral Focal Burn Injury Is Followed by Long-Lasting Bilateral Allodynia and Neuronal Hyperexcitability in Spinal Cord Dorsal Horn. J. Pain 2010, 11, 119–130. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H. Peripheral nerve injury induced changes in the spinal cord and strategies to counteract/enhance the changes to promote nerve regeneration. Neural Regen. Res. 2019, 15, 189–198. [Google Scholar] [CrossRef]
- Kowalske, K.; Holavanahalli, R.; Helm, P. Neuropathy After Burn Injury. J. Burn Care Rehabil. 2001, 22, 353–357. [Google Scholar] [CrossRef]
- Ji, R.-R.; Xu, Z.-Z.; Gao, Y.-J. Emerging targets in neuroinflammation-driven chronic pain. Nat. Rev. Drug Discov. 2014, 13, 533–548. [Google Scholar] [CrossRef] [Green Version]
- Huo, W.; Zhang, Y.; Liu, Y.; Lei, Y.; Sun, R.; Zhang, W.; Huang, Y.; Mao, Y.; Wang, C.; Ma, Z.; et al. Dehydrocorydaline attenuates bone cancer pain by shifting microglial M1/M2 polarization toward the M2 phenotype. Mol. Pain 2018, 14, 1744806918781733. [Google Scholar] [CrossRef]
- Ye, W.; Wang, J.; Lin, D.; Ding, Z. The immunomodulatory role of irisin on osteogenesis via AMPK-mediated macrophage polarization. Int. J. Biol. Macromol. 2020, 146, 25–35. [Google Scholar] [CrossRef]
- Wrann, C.D.; White, J.P.; Salogiannnis, J.; Laznik-Bogoslavski, D.; Wu, J.; Ma, D.; Lin, J.D.; Greenberg, M.E.; Spiegelman, B.M. Exercise induces hippocampal BDNF through a PGC-1alpha/FNDC5 pathway. Cell Metab. 2013, 18, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.T.-C.; Guo, D.; Campanelli, D.; Frattini, F.; Mayer, F.; Zhou, L.; Kuner, R.; Heppenstall, P.A.; Knipper, M.; Hu, J. Presynaptic GABAergic inhibition regulated by BDNF contributes to neuropathic pain induction. Nat. Commun. 2014, 5, 5331. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.Y.; Song, J. The Role of Irisin in Alzheimer’s Disease. J. Clin. Med. 2018, 7, 407. [Google Scholar] [CrossRef] [Green Version]
- Wahab, F.; Drummer, C.; Mätz-Rensing, K.; Fuchs, E.; Behr, R. Irisin is expressed by undifferentiated spermatogonia and modulates gene expression in organotypic primate testis cultures. Mol. Cell. Endocrinol. 2020, 504, 110670. [Google Scholar] [CrossRef]
- Sikandar, S.; Minett, M.S.; Millet, Q.; Santana-Varela, S.; Lau, J.; Wood, J.N.; Zhao, J. Brain-derived neurotrophic factor derived from sensory neurons plays a critical role in chronic pain. Brain 2018, 141, 1028–1039. [Google Scholar] [CrossRef]
- Boucher, T.J.; Okuse, K.; Bennett, D.L.H.; Munson, J.B.; Wood, J.N.; McMahon, S.B. Potent Analgesic Effects of GDNF in Neuropathic Pain States. Science 2000, 290, 124–127. [Google Scholar] [CrossRef]
- Takasu, K.; Sakai, A.; Hanawa, H.; Shimada, T.; Suzuki, H. Overexpression of GDNF in the Uninjured DRG Exerts Analgesic Effects on Neuropathic Pain Following Segmental Spinal Nerve Ligation in Mice. J. Pain 2011, 12, 1130–1139. [Google Scholar] [CrossRef]
- Yan, Q.; Elliott, J.; Snider, W.D. Brain-derived neurotrophic factor rescues spinal motor neurons from axotomy-induced cell death. Nat. Cell Biol. 1992, 360, 753–755. [Google Scholar] [CrossRef]
- Keeler, B.E.; Liu, G.; Siegfried, R.N.; Zhukareva, V.; Murray, M.; Houlé, J.D. Acute and prolonged hindlimb exercise elicits different gene expression in motoneurons than sensory neurons after spinal cord injury. Brain Res. 2012, 1438, 8–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobianchi, S.; Casals-Diaz, L.; Jaramillo, J.; Navarro, X. Differential effects of activity dependent treatments on axonal regeneration and neuropathic pain after peripheral nerve injury. Exp. Neurol. 2013, 240, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Detloff, M.R.; Smith, E.J.; Molina, D.Q.; Ganzer, P.D.; Houlé, J.D. Acute exercise prevents the development of neuropathic pain and the sprouting of non-peptidergic (GDNF- and artemin-responsive) c-fibers after spinal cord injury. Exp. Neurol. 2014, 255, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Dun, S.L.; Lyu, R.-M.; Chen, Y.-H.; Chang, J.-K.; Luo, J.J.; Dun, N.J. Irisin-immunoreactivity in neural and non-neural cells of the rodent. Neuroscience 2013, 240, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.H.; Yaksh, T.L. The effect of spinal GABA receptor agonists on tactile allodynia in a surgically-induced neuropathic pain model in the rat. Pain 1997, 70, 15–22. [Google Scholar] [CrossRef]
- Braz, J.M.; Sharif-Naeini, R.; Vogt, D.; Kriegstein, A.; Alvarez-Buylla, A.; Rubenstein, J.L.; Basbaum, A.I. Forebrain GABAergic Neuron Precursors Integrate into Adult Spinal Cord and Reduce Injury-Induced Neuropathic Pain. Neuron 2012, 74, 663–675. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Lei, Y.; Tian, Y.; Xu, S.; Shen, X.; Wu, H.; Bao, S.; Wang, F. The etiological contribution of GABAergic plasticity to the pathogenesis of neuropathic pain. Mol. Pain 2019, 15, 1744806919847366. [Google Scholar] [CrossRef]
- Roberts, L.D.; Ashmore, T.; McNally, B.D.; Murfitt, S.A.; Fernandez, B.O.; Feelisch, M.; Lindsay, R.; Siervo, M.; Williams, E.A.; Murray, A.J.; et al. Inorganic nitrate mimics exercise-stimulated muscular fiber-Type switching and myokine and g-Aminobutyric acid release. Diabetes 2017, 71, 674–688. [Google Scholar] [CrossRef] [Green Version]
- Pour, M.B.; Bayat, M.; Golab, F.; Eftekharzadeh, M.; Katebi, M.; Soleimani, M.; Karimzadeh, F. The effect of exercise on GABA signaling pathway in the model of chemically induced seizures. Life Sci. 2019, 232, 116667. [Google Scholar] [CrossRef]
- Coxon, J.P.; Cash, R.F.H.; Hendrikse, J.J.; Rogasch, N.C.; Stavrinos, E.; Suo, C.; Yücel, M. GABA concentration in sensorimotor cortex following high-intensity exercise and relationship to lactate levels. J. Physiol. 2017, 596, 691–702. [Google Scholar] [CrossRef]
- Maddock, R.J.; Casazza, G.A.; Fernandez, D.H.; Maddock, M.I. Acute Modulation of Cortical Glutamate and GABA Content by Physical Activity. J. Neurosci. 2016, 36, 2449–2457. [Google Scholar] [CrossRef]
- Tai, M. Gene transfer of glial cell line-derived neurotrophic factor promotes functional recovery following spinal cord contusion. Exp. Neurol. 2003, 183, 508–515. [Google Scholar] [CrossRef]
- Hu, T.-H.; Huang, C.-C.; Liu, L.-F.; Lin, P.-R.; Liu, S.-Y.; Chang, H.-W.; Changchien, C.-S.; Lee, C.-M.; Chuang, J.-H.; Tai, M.H. Expression of hepatoma-derived growth factor in hepatocellular carcinoma. Cancer 2003, 98, 1444–1456. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-H.; Yang, S.-M.; Lo, J.-J.; Wu, S.-H.; Tai, M.-H. Irisin Gene Delivery Ameliorates Burn-Induced Sensory and Motor Neuropathy. Int. J. Mol. Sci. 2020, 21, 7798. https://doi.org/10.3390/ijms21207798
Huang S-H, Yang S-M, Lo J-J, Wu S-H, Tai M-H. Irisin Gene Delivery Ameliorates Burn-Induced Sensory and Motor Neuropathy. International Journal of Molecular Sciences. 2020; 21(20):7798. https://doi.org/10.3390/ijms21207798
Chicago/Turabian StyleHuang, Shu-Hung, Shih-Ming Yang, Jing-Jou Lo, Sheng-Hua Wu, and Ming-Hong Tai. 2020. "Irisin Gene Delivery Ameliorates Burn-Induced Sensory and Motor Neuropathy" International Journal of Molecular Sciences 21, no. 20: 7798. https://doi.org/10.3390/ijms21207798
APA StyleHuang, S. -H., Yang, S. -M., Lo, J. -J., Wu, S. -H., & Tai, M. -H. (2020). Irisin Gene Delivery Ameliorates Burn-Induced Sensory and Motor Neuropathy. International Journal of Molecular Sciences, 21(20), 7798. https://doi.org/10.3390/ijms21207798