Facilitation of Bone Healing Processes Based on the Developmental Function of Meox2 in Tooth Loss Lesion

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
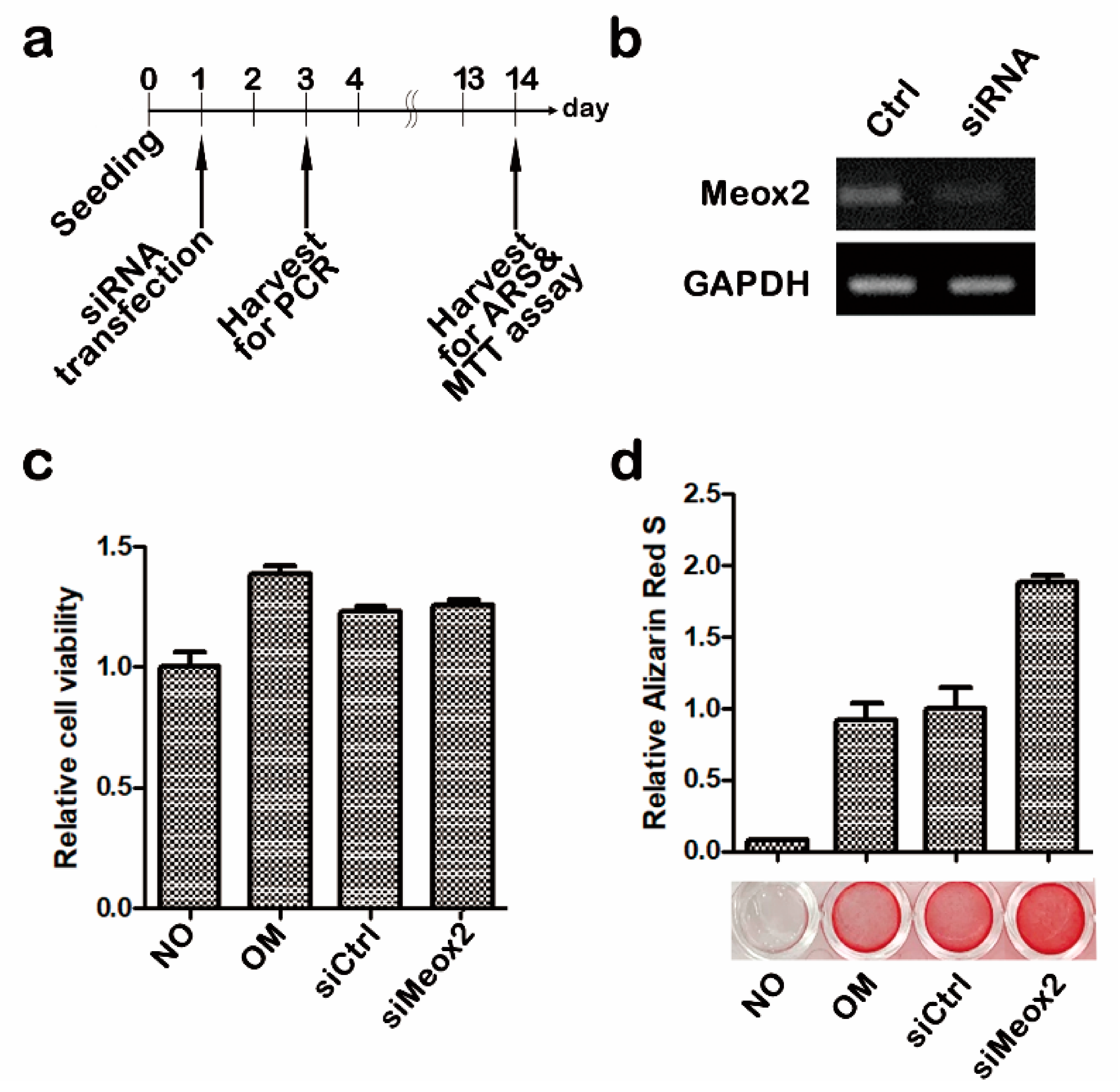
2.1. In Vitro Assays after Knocking Down of Meox2 on Hpdlfs
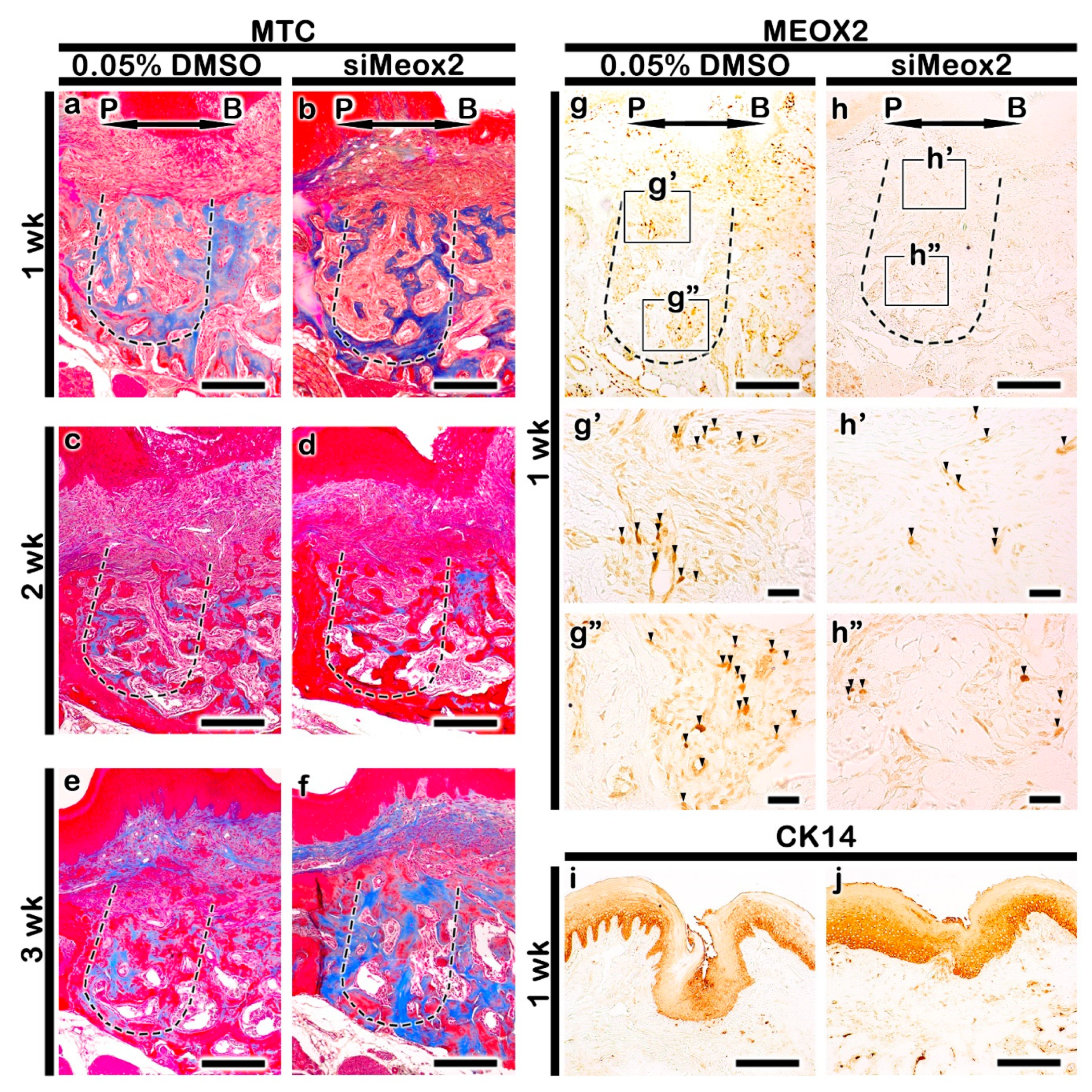
2.2. Altered Histogenesis and Protein Localizations after the Knockdown of Meox2
2.3. Localization Patterns of Osteoblast Related Factor
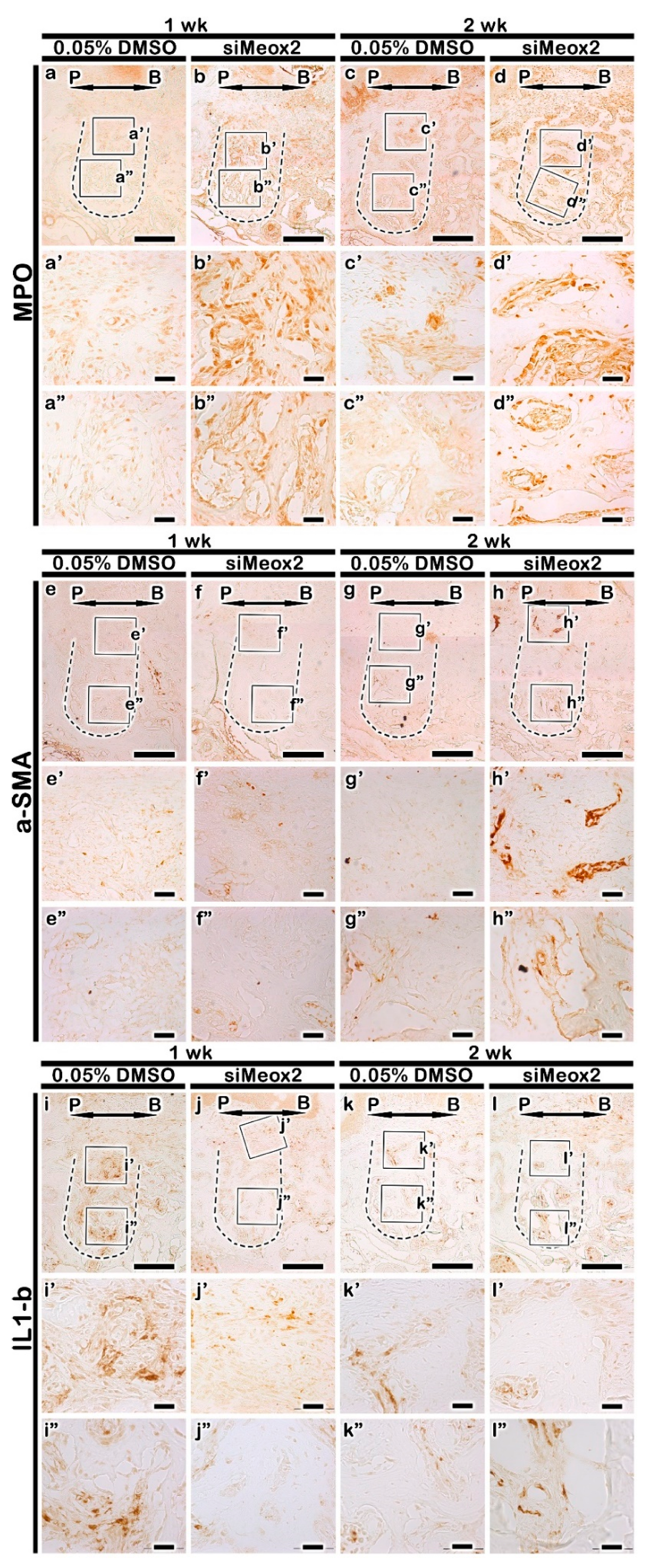
2.4. Immunolocalization Patterns of Inflammatory Molecules
2.5. Micro-CT Evaluations of Bone Formation after siRNA Delivery
3. Discussion
3.1. Periodontal Disease and Various Clinical Approaches
3.2. Application of Developmental Mechanisms on Functional Restoration
3.3. Facilitation of Bone Formation by Knocking Down of Meox2
3.4. Functional Evaluation of Meox2 for Modulating Inflammation
3.5. Tissue Regeneration Using Developmental Regulating Gene Application
4. Materials and Methods
4.1. In Vitro Cell Cultivation Experiment
4.1.1. Cell Culture and siRNA Transfection
4.1.2. Osteogenic Differentiation
4.1.3. Proliferation Assay
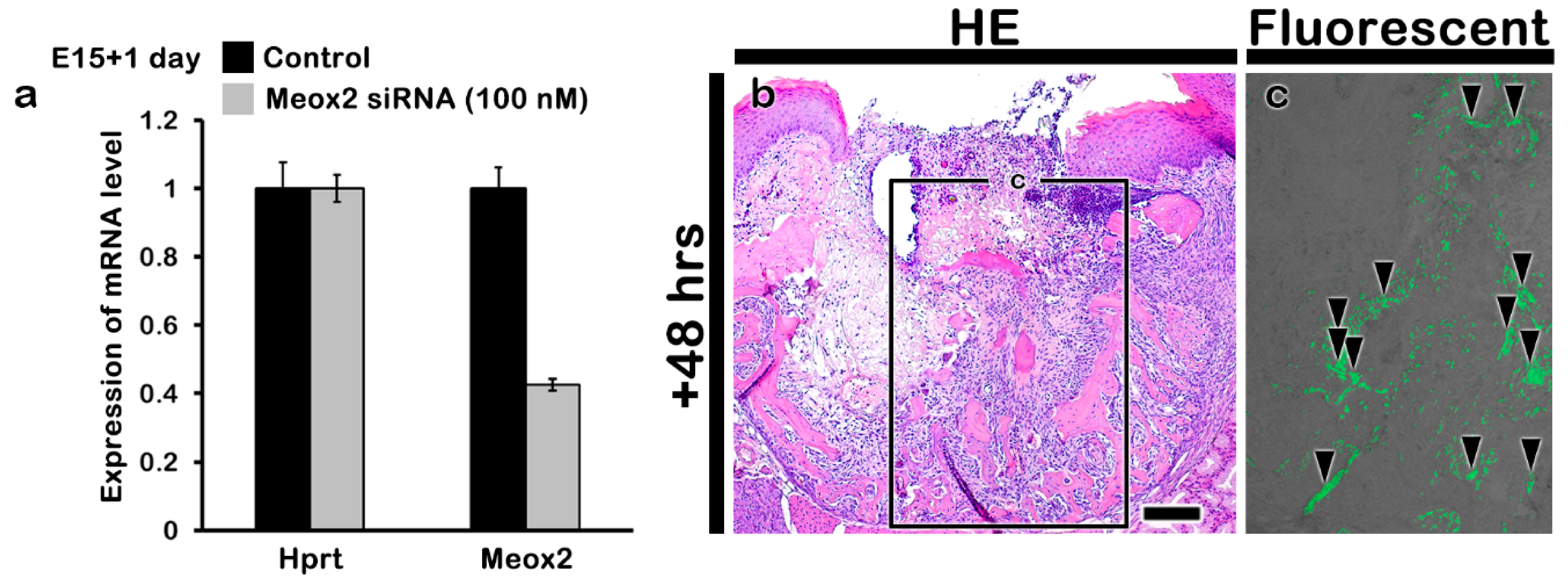
4.2. In Vivo Animal Experiment
4.2.1. Animals
4.2.2. Periodontitis Model System and Tooth Extraction
4.2.3. siRNA Delivery in the Extraction Socket
4.2.4. Histology and Immunostaining
4.2.5. Micro-CT Evaluation
4.3. In Vitro Mesenchymal Tissue Cultivation and RT-qPCR Evaluation
4.4. Photography and Image Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yu, T.; Klein, O.D. Molecular and cellular mechanisms of tooth development, homeostasis and repair. Development 2020, 147, dev184754. [Google Scholar] [CrossRef]
- Neupane, S.; Aryal, Y.P.; Kim, T.-Y.; Yeon, C.-Y.; An, C.-H.; Kim, J.-Y.; Yamamoto, H.; Lee, Y.; Sohn, W.-J.; Kim, J.-Y. Signaling Modulations of miR-206-3p in Tooth Morphogenesis. Int. J. Mol. Sci. 2020, 21, 5251. [Google Scholar] [CrossRef]
- Lin, Y.; Zheng, L.; Fan, L.; Kuang, W.; Guo, R.; Lin, J.; Wu, J.; Tan, J. The Epigenetic Regulation in Tooth Development and Regeneration. Curr. Stem Cell Res. Ther. 2017, 13. [Google Scholar] [CrossRef]
- Balic, A.; Thesleff, I. Tissue Interactions Regulating Tooth Development and Renewal. In Current Topics in Developmental Biology; Academic Press: New York, NY, USA, 2015; Volume 115, pp. 157–186. ISBN 9780124081413. [Google Scholar]
- Jussila, M.; Thesleff, I. Signaling networks regulating tooth organogenesis and regeneration, and the specification of dental mesenchymal and epithelial cell lineages. Cold Spring Harb. Perspect. Biol. 2012, 4, a008425. [Google Scholar] [CrossRef]
- Nakamura, T.; Jimenez-Rojo, L.; Koyama, E.; Pacifici, M.; de Vega, S.; Iwamoto, M.; Fukumoto, S.; Unda, F.; Yamada, Y. Epiprofin Regulates Enamel Formation and Tooth Morphogenesis by Controlling Epithelial-Mesenchymal Interactions During Tooth Development. J. Bone Miner. Res. 2017, 32, 601–610. [Google Scholar] [CrossRef] [Green Version]
- Puthiyaveetil, J.S.V.; Kota, K.; Chakkarayan, R.; Chakkarayan, J.; Thodiyil, A.K.P. Epithelial—Mesenchymal interactions in tooth development and the significant role of growth factors and genes with emphasis on mesenchyme—A review. J. Clin. Diagn. Res. 2016, 10, ZE05–ZE09. [Google Scholar]
- Santosh, A.B.R.; Jones, T.J. The epithelial-mesenchymal interactions: Insights into physiological and pathological aspects of oral tissues. Oncol. Rev. 2014, 8, 1–5. [Google Scholar]
- Tummers, M.; Thesleff, I. The importance of signal pathway modulation in all aspects of tooth development. J. Exp. Zool. B Mol. Dev. Evol. 2009, 312, 309–319. [Google Scholar] [CrossRef]
- Zhou, T.; Pan, J.; Wu, P.; Huang, R.; Du, W.; Zhou, Y.; Wan, M.; Fan, Y.; Xu, X.; Zhou, X.; et al. Dental Follicle Cells: Roles in Development and beyond. Stem Cells Int. 2019, 2019, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Cho, S.W.; Hwang, H.J.; Lee, M.J.; Lee, J.M.; Cai, J.; Choi, S.H.; Kim, C.K.; Jung, H.S. Evidence for expansion-based temporal BMP4/NOGGIN interactions in specifying periodontium morphogenesis. Cell Tissue Res. 2007, 330, 123–132. [Google Scholar] [CrossRef]
- Cho, M.I.; Garant, P.R. Development and general structure of the periodontium. Periodontology 2000 2000, 24, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ren, J.; Li, R.; Guan, C.; Feng, Z.; Bao, B.; Wang, W.; Zhou, C. Tooth Regeneration: Insights from Tooth Development and Spatial-Temporal Control of Bioactive Drug Release. Stem Cell Rev. Rep. 2020, 16, 41–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, T.; Bakker, A.D.; Everts, V.; Smit, T.H. The intricate anatomy of the periodontal ligament and its development: Lessons for periodontal regeneration. J. Periodontal Res. 2017, 52, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Parada, C.; Chai, Y. Cellular and molecular mechanisms of tooth root development. Development 2017, 144, 374–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Ruan, J.; Weir, M.D.; Ren, K.; Schneider, A.; Wang, P.; Oates, T.W.; Chang, X.; Xu, H.H.K. Periodontal Bone-Ligament-Cementum Regeneration via Scaffolds and Stem Cells. Cells 2019, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Bosshardt, D.D.; Stadlinger, B.; Terheyden, H. Cell-to-cell communication—Periodontal regeneration. Clin. Oral Implant. Res. 2015, 26, 229–239. [Google Scholar] [CrossRef]
- Gasner, N.S.; Schure, R.S. Periodontal Disease. In StatPearls; StatPearls Publishing: Petersburg, FL, USA, 2020. [Google Scholar]
- Dentino, A.; Lee, S.; Mailhot, J.; Hefti, A.F. Principles of periodontology. Periodontology 2000 2013, 61, 16–53. [Google Scholar] [CrossRef] [Green Version]
- Könönen, E.; Gursoy, M.; Gursoy, U. Periodontitis: A Multifaceted Disease of Tooth-Supporting Tissues. J. Clin. Med. 2019, 8, 1135. [Google Scholar] [CrossRef] [Green Version]
- Hoare, A.; Soto, C.; Rojas-Celis, V.; Bravo, D. Chronic Inflammation as a Link between Periodontitis and Carcinogenesis. Mediat. Inflamm. 2019, 2019, 1029857. [Google Scholar] [CrossRef] [Green Version]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef]
- Naruishi, K.; Nagata, T. Biological effects of interleukin-6 on Gingival Fibroblasts: Cytokine regulation in periodontitis. J. Cell. Physiol. 2018, 233, 6393–6400. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, A.; Gigante, I.; Colucci, S.; Grano, M. Periodontal Disease: Linking the Primary Inflammation to Bone Loss. Clin. Dev. Immunol. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochran, D.L. Inflammation and Bone Loss in Periodontal Disease. J. Periodontol. 2008, 79, 1569–1576. [Google Scholar] [CrossRef]
- Slots, J. Periodontitis: Facts, fallacies and the future. Periodontology 2000 2017, 75, 7–23. [Google Scholar] [CrossRef]
- Mikkola, M.L.; Millar, S.E. The mammary bud as a skin appendage: Unique and shared aspects of development. J. Mammary Gland Biol. Neoplasia 2006, 11, 187–203. [Google Scholar] [CrossRef]
- Jernvall, J.; Thesleff, I. Tooth shape formation and tooth renewal: Evolving with the same signals. Development 2012, 139, 3487–3497. [Google Scholar] [CrossRef] [Green Version]
- Neupane, S.; Adhikari, N.; Jung, J.-K.; An, C.-H.; Lee, S.; Jun, J.-H.; Kim, J.-Y.; Lee, Y.; Sohn, W.-J.; Kim, J.-Y. Regulation of mesenchymal signaling in palatal mucosa differentiation. Histochem. Cell Biol. 2018, 149, 143–152. [Google Scholar] [CrossRef]
- Jin, J.Z.; Ding, J. Analysis of Meox-2 mutant mice reveals a novel postfusion-based cleft palate. Dev. Dyn. 2006, 235, 539–546. [Google Scholar] [CrossRef]
- Li, Q.; Ding, J. Gene expression analysis reveals that formation of the mouse anterior secondary palate involves recruitment of cells from the posterior side. Int. J. Dev. Biol. 2007, 51, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Mankoo, B.S.; Skuntz, S.; Harrigan, I.; Grigorieva, E.; Candia, A.; Wright, C.V.E.; Arnheiter, H.; Pachnis, V. The concerted action of Meox homeobox genes is required upstream of genetic pathways essential for the formation, patterning and differentiation of somites. Development 2003, 130, 4655–4664. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Song, Y.; Zhao, X.; Zhang, X.; Fermin, C.; Chen, Y. Rescue of cleft palate in Msx1-deficient mice by transgenic Bmp4 reveals a network of BMP and Shh signaling in the regulation of mammalian palatogenesis. Development 2002, 129, 4135–4146. [Google Scholar]
- Adhikari, N.; Neupane, S.; Aryal, Y.P.; Choi, M.; Sohn, W.J.; Lee, Y.; Jung, J.K.; Ha, J.H.; Choi, S.Y.; Suh, J.Y.; et al. Effects of oleanolic acid acetate on bone formation in an experimental periodontitis model in mice. J. Periodontal Res. 2019, 54, 533–545. [Google Scholar] [CrossRef]
- Graziani, F.; Karapetsa, D.; Alonso, B.; Herrera, D. Nonsurgical and surgical treatment of periodontitis: How many options for one disease? Periodontology 2000 2017, 75, 152–188. [Google Scholar] [CrossRef]
- Deas, D.E.; Moritz, A.J.; Sagun, R.S.; Gruwell, S.F.; Powell, C.A. Scaling and root planing vs. conservative surgery in the treatment of chronic periodontitis. Periodontology 2000 2016, 71, 128–139. [Google Scholar] [CrossRef]
- Smiley, C.J.; Tracy, S.L.; Abt, E.; Michalowicz, B.S.; John, M.T.; Gunsolley, J.; Cobb, C.M.; Rossmann, J.; Harrel, S.K.; Forrest, J.L.; et al. Evidence-based clinical practice guideline on the nonsurgical treatment of chronic periodontitis by means of scaling and root planing with or without adjuncts. J. Am. Dent. Assoc. 2015, 146, 525–535. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Cheng, Z.; Wang, Y.; Huang, T.; Lai, K.; Du, X.; Jiang, Z.; Yang, G. Adenovirus-Mediated LAMA3 Transduction Enhances Hemidesmosome Formation and Periodontal Reattachment during Wound Healing. Mol. Ther.- Methods Clin. Dev. 2020, 18, 291–303. [Google Scholar] [CrossRef]
- Li, J.; Zhang, F.; Zhang, N.; Geng, X.; Meng, C.; Wang, X.; Yang, Y. Osteogenic capacity and cytotherapeutic potential of periodontal ligament cells for periodontal regeneration In Vitro and In Vivo. PeerJ 2019, 2019, e6589. [Google Scholar] [CrossRef] [Green Version]
- Hägi, T.T.; Klemensberger, S.; Bereiter, R.; Nietzsche, S.; Cosgarea, R.; Flury, S.; Lussi, A.; Sculean, A.; Eick, S. A biofilm pocket model to evaluate different non-surgical periodontal treatment modalities in terms of biofilm removal and reformation, surface alterations and attachment of periodontal ligament fibroblasts. PLoS ONE 2015, 10, e0131056. [Google Scholar] [CrossRef] [Green Version]
- Draenert, F.G.; Huetzen, D.; Neff, A.; Mueller, W.E.G. Vertical bone augmentation procedures: Basics and techniques in dental implantology. J. Biomed. Mater. Res.- Part A 2014, 102, 1605–1613. [Google Scholar] [CrossRef]
- Salvi, G.E.; Mischler, D.C.; Schmidlin, K.; Matuliene, G.; Pjetursson, B.E.; Brägger, U.; Lang, N.P. Risk factors associated with the longevity of multi-rooted teeth. Long-term outcomes after active and supportive periodontal therapy. J. Clin. Periodontol. 2014, 41, 701–707. [Google Scholar] [CrossRef]
- Iguchi, S.; Suzuki, D.; Kawano, E.; Mashimo, T.; Kajiya, M.; Toriumi, T.; Kawai, T.; Kurihara, H.; Isokawa, K.; Sato, S.; et al. Effect of local bone marrow stromal cell administration on ligature-induced periodontitis in mice. J. Oral Sci. 2017, 59, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.J.; Hu, B.B.; Shi, X.L.; Ren, M.M.; Yu, W.B.; Cen, S.D.; Hu, R.D.; Deng, H. Baicalein enhances the osteogenic differentiation of human periodontal ligament cells by activating the Wnt/β-catenin signaling pathway. Arch. Oral Biol. 2017, 78, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Min, K.K.; Neupane, S.; Adhikari, N.; Sohn, W.J.; An, S.Y.; Kim, J.Y.; An, C.H.; Lee, Y.; Kim, Y.G.; Park, J.W.; et al. Effects of resveratrol on bone-healing capacity in the mouse tooth extraction socket. J. Periodontal Res. 2020, 55, 247–257. [Google Scholar] [CrossRef]
- Bhattarai, G.; Poudel, S.B.; Kook, S.H.; Lee, J.C. Resveratrol prevents alveolar bone loss in an experimental rat model of periodontitis. Acta Biomater. 2016, 29, 398–408. [Google Scholar] [CrossRef]
- Tang, H.; Mattheos, N.; Yao, Y.; Jia, Y.; Ma, L.; Gong, P. In Vivo osteoprotegerin gene therapy preventing bone loss induced by periodontitis. J. Periodontal Res. 2015, 50, 434–443. [Google Scholar] [CrossRef]
- Wu, Y.H.; Kuraji, R.; Taya, Y.; Ito, H.; Numabe, Y. Effects of theaflavins on tissue inflammation and bone resorption on experimental periodontitis in rats. J. Periodontal Res. 2018, 53, 1009–1019. [Google Scholar] [CrossRef]
- Iyyanar, P.P.R.; Thangaraj, M.P.; Frank Eames, B.; Nazaralia, A.J. HtrA1 is a novel transcriptional target of Runx2 that promotes osteogenic differentiation. Cell. Physiol. Biochem. 2019, 53, 832–850. [Google Scholar] [CrossRef]
- Valcourt, U.; Thuault, S.; Pardali, K.; Heldin, C.-H.; Moustakas, A. Functional role of Meox2 during the epithelial cytostatic response to TGF-β. Mol. Oncol. 2007, 1, 55–71. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Hajishengallis, G. Optimization of the ligature-induced periodontitis model in mice. J. Immunol. Methods 2013, 394, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhou, Z.; Mao, Z.; Shen, M.; Chen, N.; Miao, D. Role of p53 deficiency in socket healing after tooth extractions. J. Mol. Histol. 2020, 51, 55–65. [Google Scholar] [CrossRef]
- Nakashima, K.; de Crombrugghe, B. Transcriptional mechanisms in osteoblast differentiation and bone formation. Trends Genet. 2003, 19, 458–466. [Google Scholar] [CrossRef]
- Simon, D.; Derer, A.; Andes, F.T.; Lezuo, P.; Bozec, A.; Schett, G.; Herrmann, M.; Harre, U. Galectin-3 as a novel regulator of osteoblast-osteoclast interaction and bone homeostasis. Bone 2017, 105, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, E.F. Cellular mechanisms of bone remodeling. Rev. Endocr. Metab. Disord. 2010, 11, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahama, K.I. Cellular communications in bone homeostasis and repair. Cell. Mol. Life Sci. 2010, 67, 4001–4009. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast–osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef]
- Goodman, S.B.; Pajarinen, J.; Yao, Z.; Lin, T. Inflammation and Bone Repair: From Particle Disease to Tissue Regeneration. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef]
- Graves, D.T.; Fine, D.; Teng, Y.-T.A.; Van Dyke, T.E.; Hajishengallis, G. The use of rodent models to investigate host-bacteria interactions related to periodontal diseases. J. Clin. Periodontol. 2008, 35, 89–105. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Wang, Q.; Chen, Q. The cytokine network involved in the host immune response to periodontitis. Int. J. Oral Sci. 2019, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of Bone Resorption in Periodontitis. J. Immunol. Res. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Douville, J.M.; Cheung, D.Y.C.; Herbert, K.L.; Moffatt, T.; Wigle, J.T. Mechanisms of MEOX1 and MEOX2 Regulation of the Cyclin Dependent Kinase Inhibitors p21CIP1/WAF1 and p16INK4a in Vascular Endothelial Cells. PLoS ONE 2011, 6, e29099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Cao, H.; Yang, Y.; Zhou, Y.; Gu, Y.; Zhao, X.; Zhang, Y.; Zhao, Z.; Zhang, L.; Yin, J. Effects of vascular endothelial cells on osteogenic differentiation of noncontact co-cultured periodontal ligament stem cells under hypoxia. J. Periodontal Res. 2013, 48, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, R.T.; Yang, S.; Rutherford, R.B.; Krebsbach, P.H.; Zhao, M.; Wang, D. Gene Therapy Approaches for Bone Regeneration. Cells Tissues Organs 2004, 176, 95–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewkes, B.C.; Barlow, L.A.; Delay, E.R. Effect of radiation on sucrose detection thresholds of mice. Chem. Senses 2018, 43, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, S.Y.; Lee, Y.J.; Neupane, S.; Jun, J.H.; Kim, J.Y.; Lee, Y.; Choi, K.S.; An, C.H.; Suh, J.Y.; Shin, H.I.; et al. Effects of vascular formation during alveolar bone process morphogenesis in mice. Histochem. Cell Biol. 2017, 148, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, N.; Neupane, S.; Gwon, G.J.; Kim, J.Y.; An, C.H.; Lee, S.; Sohn, W.J.; Lee, Y.; Kim, J.Y. Grhl3 modulates epithelial structure formation of the circumvallate papilla during mouse development. Histochem. Cell Biol. 2017, 147, 5–16. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-Y.; Park, J.-K.; Prasad Aryal, Y.; Lee, E.-S.; Neupane, S.; Sung, S.; Pokharel, E.; Yeon, C.-Y.; Kim, J.-Y.; Jung, J.-K.; et al. Facilitation of Bone Healing Processes Based on the Developmental Function of Meox2 in Tooth Loss Lesion. Int. J. Mol. Sci. 2020, 21, 8701. https://doi.org/10.3390/ijms21228701
Kim T-Y, Park J-K, Prasad Aryal Y, Lee E-S, Neupane S, Sung S, Pokharel E, Yeon C-Y, Kim J-Y, Jung J-K, et al. Facilitation of Bone Healing Processes Based on the Developmental Function of Meox2 in Tooth Loss Lesion. International Journal of Molecular Sciences. 2020; 21(22):8701. https://doi.org/10.3390/ijms21228701
Chicago/Turabian StyleKim, Tae-Young, Jae-Kyung Park, Yam Prasad Aryal, Eui-Seon Lee, Sanjiv Neupane, Shijin Sung, Elina Pokharel, Chang-Yeol Yeon, Ji-Youn Kim, Jae-Kwang Jung, and et al. 2020. "Facilitation of Bone Healing Processes Based on the Developmental Function of Meox2 in Tooth Loss Lesion" International Journal of Molecular Sciences 21, no. 22: 8701. https://doi.org/10.3390/ijms21228701
APA StyleKim, T. -Y., Park, J. -K., Prasad Aryal, Y., Lee, E. -S., Neupane, S., Sung, S., Pokharel, E., Yeon, C. -Y., Kim, J. -Y., Jung, J. -K., Yamamoto, H., An, C. -H., Lee, Y., Sohn, W. -J., Jang, I. -H., An, S. -Y., & Kim, J. -Y. (2020). Facilitation of Bone Healing Processes Based on the Developmental Function of Meox2 in Tooth Loss Lesion. International Journal of Molecular Sciences, 21(22), 8701. https://doi.org/10.3390/ijms21228701