The Role of Cholesterol on Triterpenoid Saponin-Induced Endolysosomal Escape of a Saporin-Based Immunotoxin

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
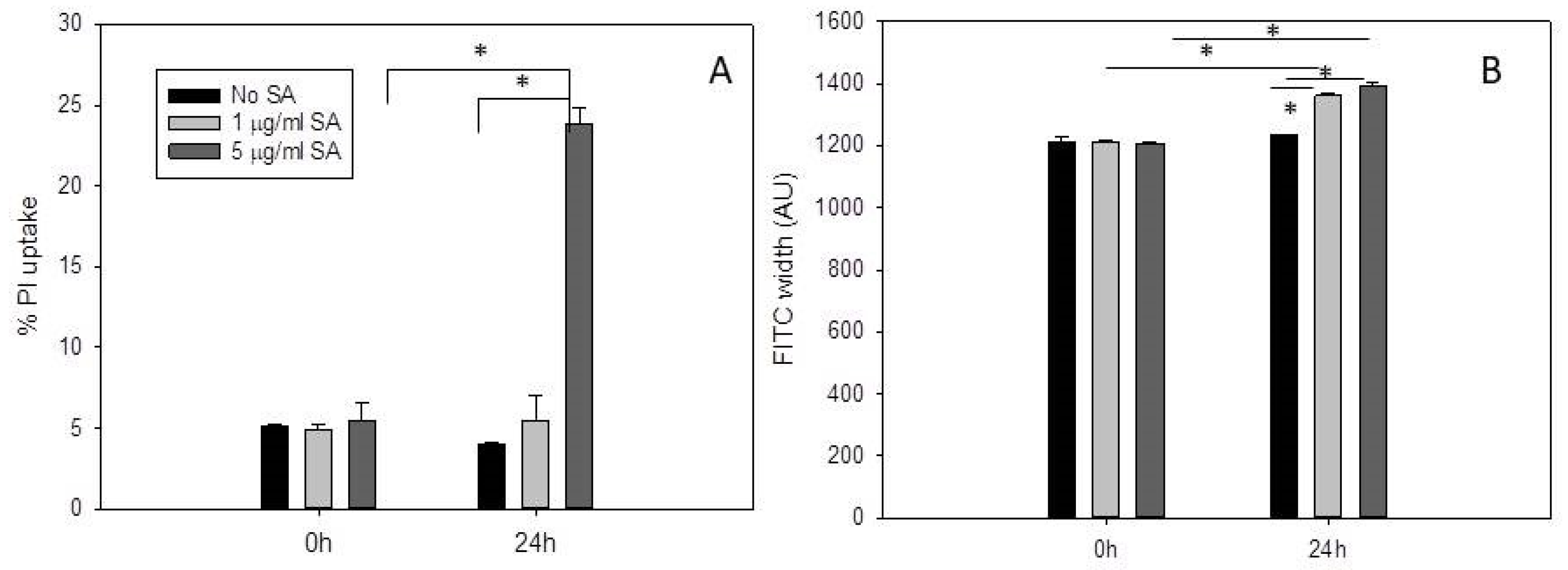
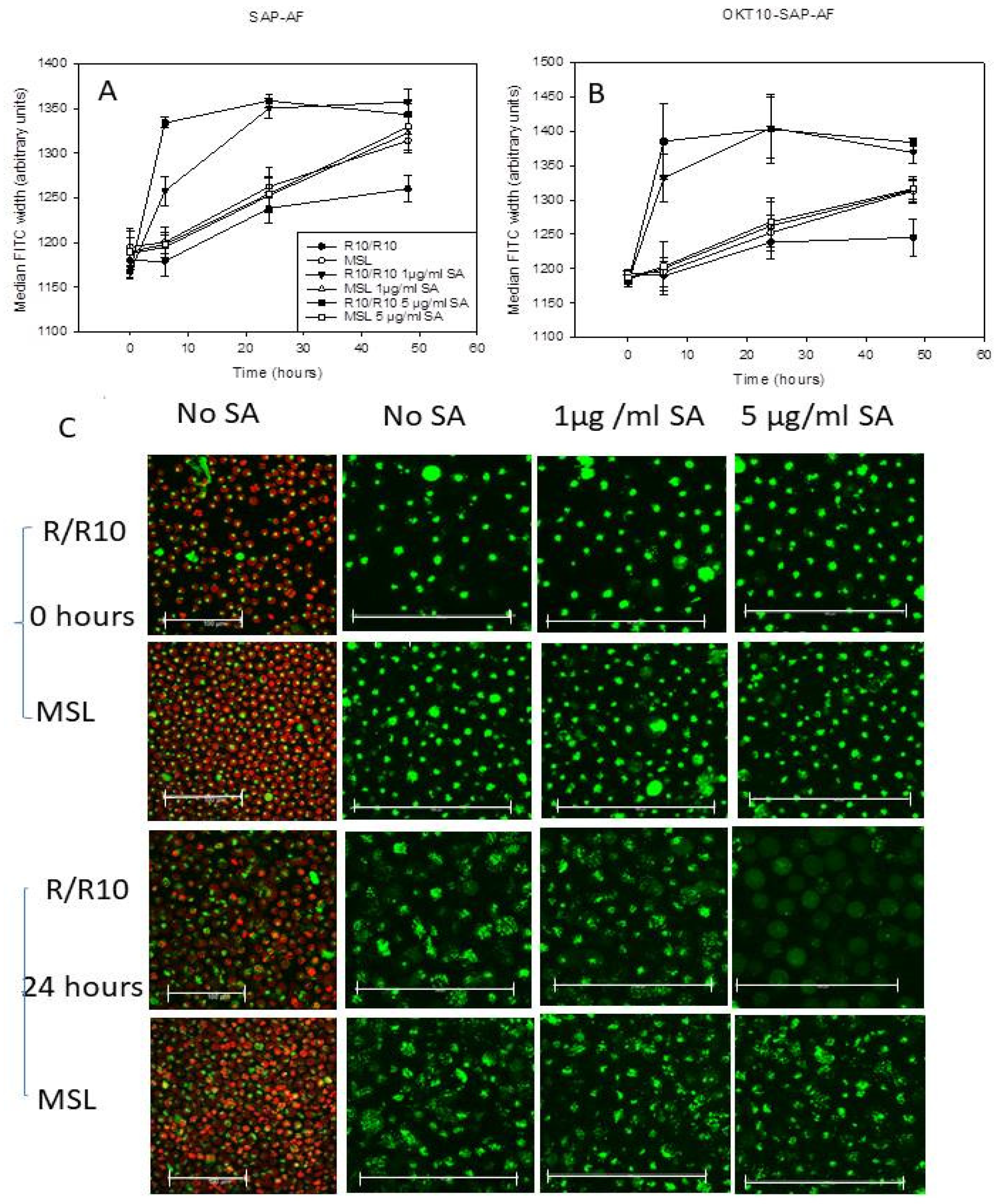
2.1. Propidium Iodide Uptake Increases in Daudi Cells Incubated with 5 µg/mL SA for 24 H after Pre-Exposure to OKT10-SAP-AF
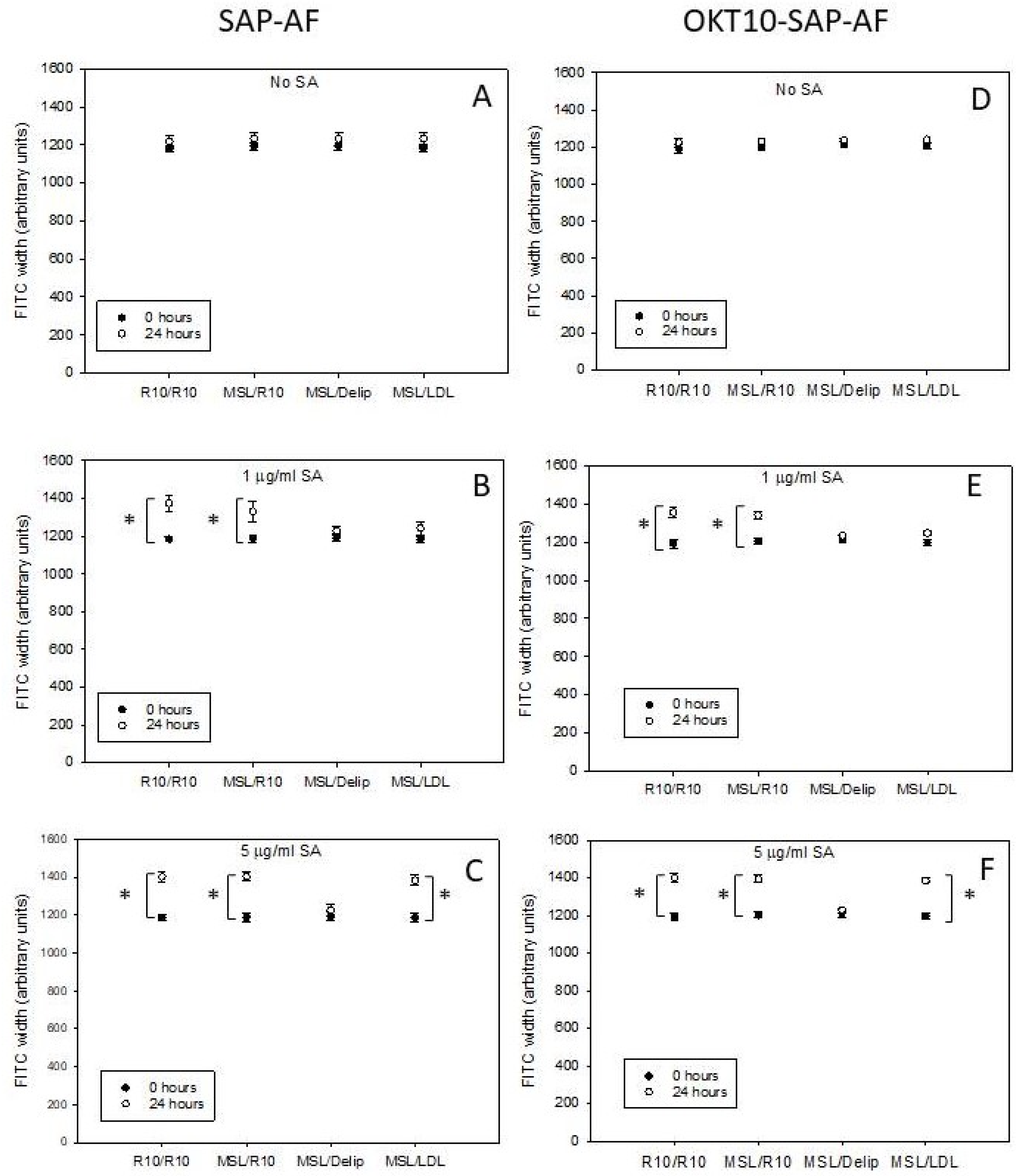
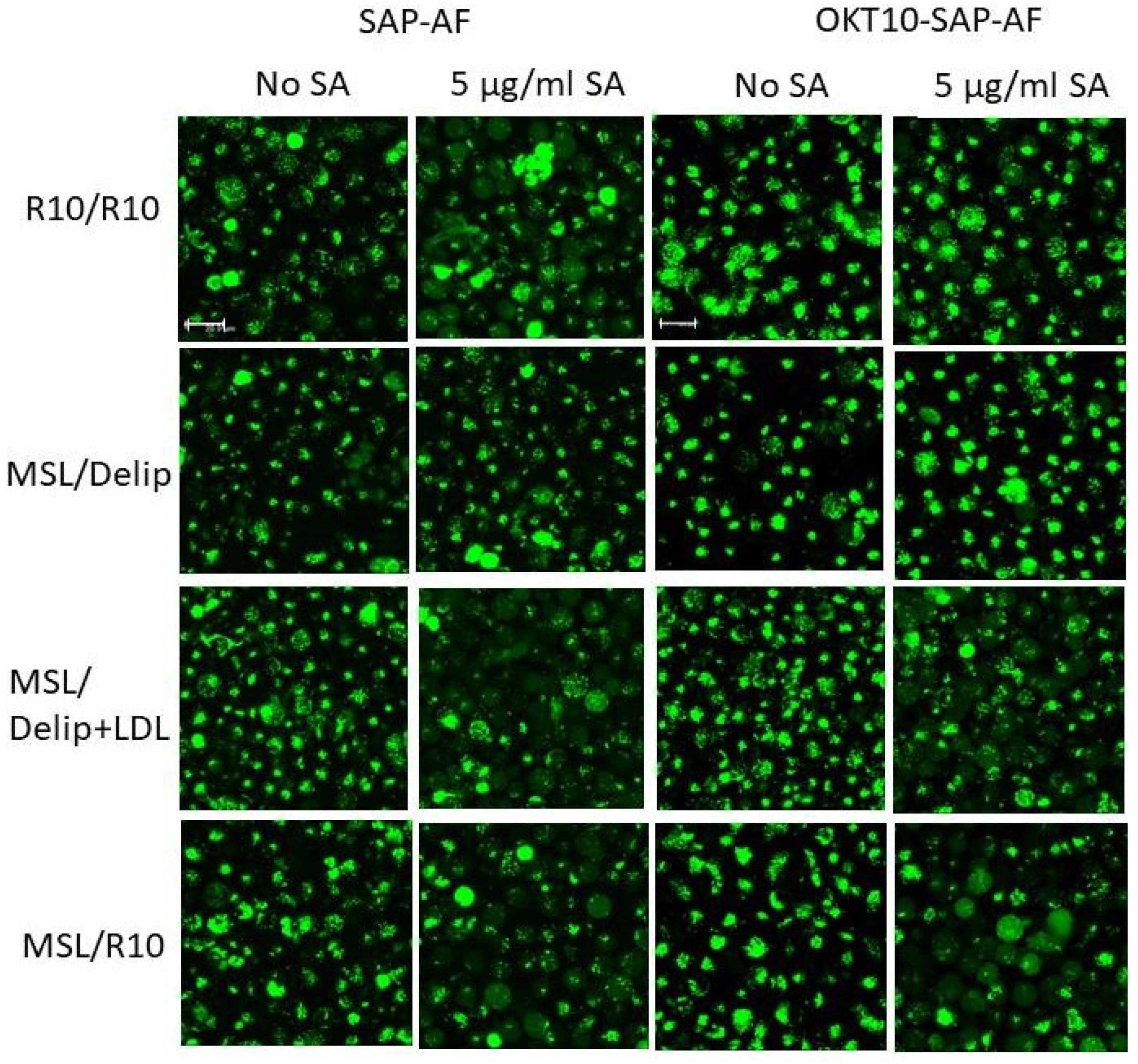
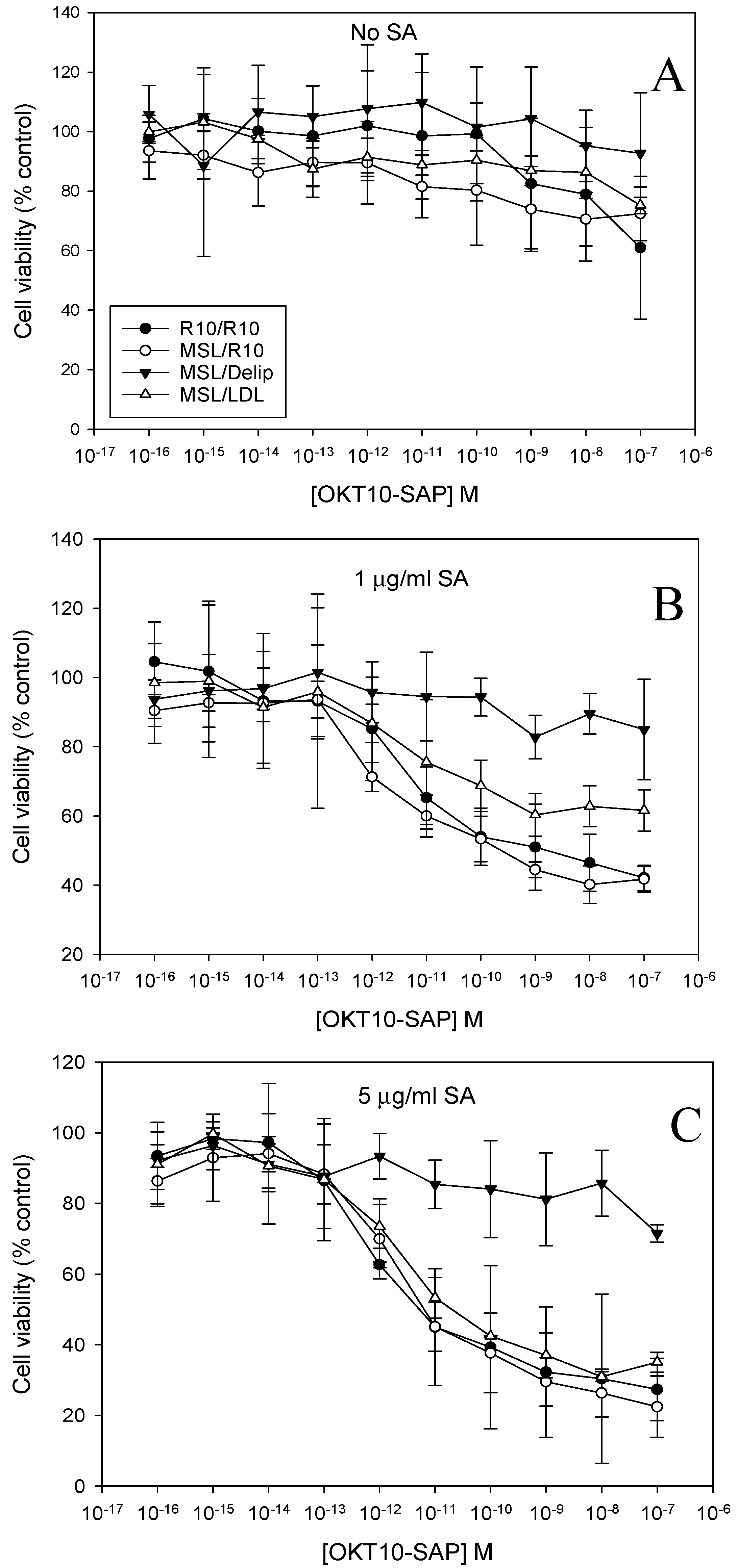
2.2. Lipid Depletion Reduces Endolysosomal Escape of OKT10-SAP-AF and SAP-AF Induced by the Presence of 1 and 5 µg/mL SA
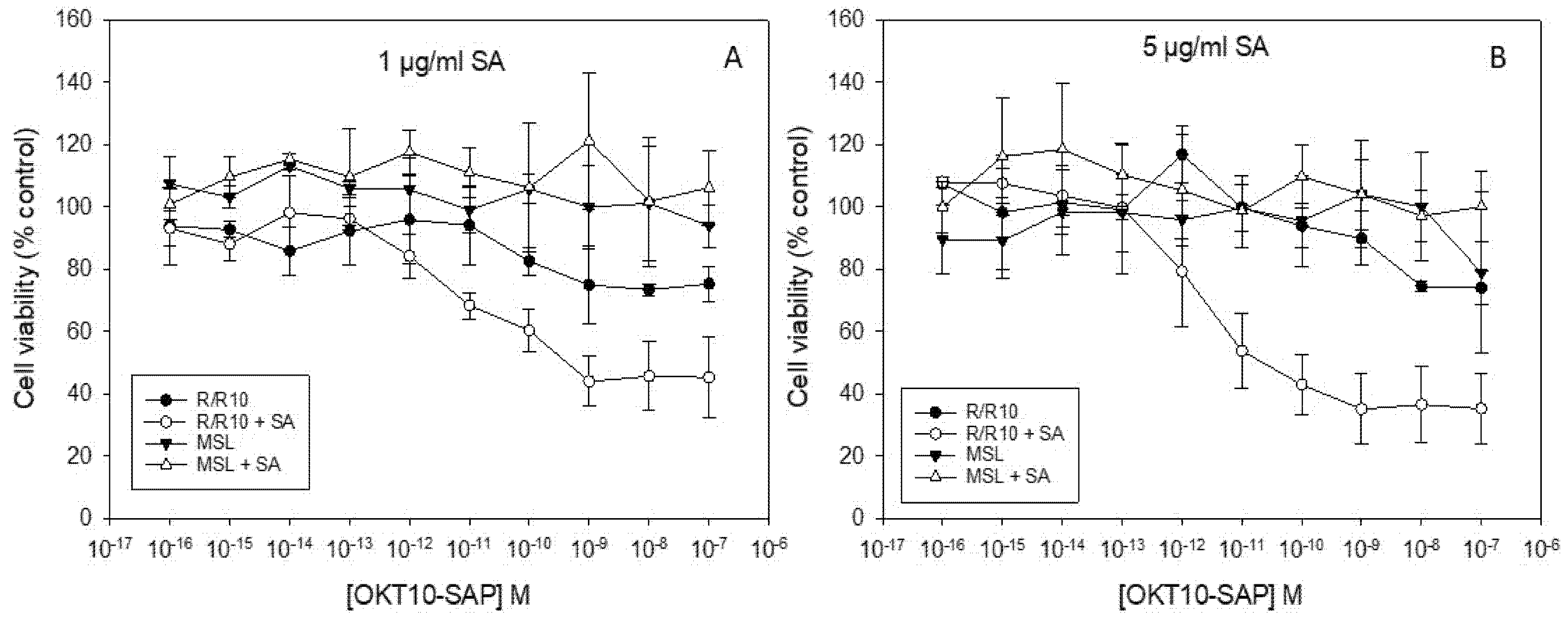
2.3. Lipid Depletion Abrogates SA Augmentation of OKT10-SAP and Saporin Using 1 and 5 µg/mL SA
2.4. LDL Repletion of Lipid Depleted Daudi Cells Restores OKT10-SAP-AF and AF-SAP Endolysosomal Escape Induced in the Presence of 5 µg/mL SA But Not 1 µg/mL SA
2.5. LDL Repletion of Lipid Depleted Daudi Cells Restores SA Augmentation of OKT10-SAP
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Saponinum Album
4.3. IT
4.4. Fluorescent Labelling of Saporin and OKT10-Saporin
4.5. Delipidation of Foetal Calf Serum
4.6. Cholesteterol and Lipid Depletion of Daudi Cells
4.7. Cholesterol Repletion
4.8. NR12S Assay
4.9. XTT Cytotoxicity Assay
4.10. Flow Cytometry
4.11. Confocal Microscopy
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SA | Saponinum album |
| IT | Immunotoxin |
| EGF | Epidermal growth factor |
| AF | Alexa Fluor |
| SAP | Saporin |
| FCS | Foetal calf serum |
| PM | Plasma membrane |
| LDL | Low density lipoprotein |
| ECV | Endosomal carrier vesicle |
| RIP | Ribosomal inactivating protein |
| FITC | Fluorescein isothiocyanate |
| SM | Sphingomyelin |
| PC | Phosphatidylcholine |
| PE | Phosphatidylethanolamine |
| PI | Propidium iodide |
| LBPA | Lyso-bis-phosphatidic acid |
| MβCD | Methyl-beta-cyclodextrin |
| U18666 | 3 beta-(2-diethylaminoethoxy)-androstenone HCl |
References
- Gilabert-Oriol, R.; Weng, A.; Mallinckrodt, B.; Melzig, M.F.; Fuchs, H.; Thakur, M. Immunotoxins constructed with ribosome-inactivating proteins and their enhancers: A lethal cocktail with tumor specific efficacy. Curr. Pharm. Des. 2014, 20, 6584–6643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirker, R.; FitzGerald, D.J.; Hamilton, T.C.; Ozols, R.F.; Laird, W.; Frankel, A.E.; Willingham, M.C.; Pastan, I. Characterization of immunotoxins active against ovarian cancer cell lines. J. Clin. Investig. 1985, 76, 1261–1267. [Google Scholar] [CrossRef]
- Ravel, S.; Colombatti, M.; Casellas, P. Internalization and intracellular fate of anti-CD5 monoclonal antibody and anti-CD5 ricin A-chain immunotoxin in human leukemic T cells. Blood 1992, 79, 1511–1517. [Google Scholar] [CrossRef] [Green Version]
- Weng, A.; Thakur, M.; von Mallinckrodt, B.; Beceren-Braun, F.; Gilabert-Oriol, R.; Wiesner, B.; Eichhorst, J.; Bottger, S.; Melzig, M.F.; Fuchs, H. Saponins modulate the intracellular trafficking of protein toxins. J. Control. Release Off. J. Control. Release Soc. 2012, 164, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Weng, A.; Pieper, A.; Mergel, K.; von Mallinckrodt, B.; Gilabert-Oriol, R.; Gorick, C.; Wiesner, B.; Eichhorst, J.; Melzig, M.F.; et al. Macromolecular interactions of triterpenoids and targeted toxins: Role of saponins charge. Int. J. Biol. Macromol. 2013, 61, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Stirpe, F. Ribosome-inactivating proteins. Toxicon Off. J. Int. Soc. Toxinol. 2004, 44, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Dennemarker, J.; Reinheckel, T. Specific functions of lysosomal proteases in endocytic and autophagic pathways. Biochim. Biophys. Acta 2012, 1824, 34–43. [Google Scholar] [CrossRef]
- Heisler, I.; Sutherland, M.; Bachran, C.; Hebestreit, P.; Schnitger, A.; Melzig, M.F.; Fuchs, H. Combined application of saponin and chimeric toxins drastically enhances the targeted cytotoxicity on tumor cells. J. Control. Release Off. J. Control. Release Soc. 2005, 106, 123–137. [Google Scholar] [CrossRef]
- Bachran, D.; Schneider, S.; Bachran, C.; Urban, R.; Weng, A.; Melzig, M.F.; Hoffmann, C.; Kaufmann, A.M.; Fuchs, H. Epidermal growth factor receptor expression affects the efficacy of the combined application of saponin and a targeted toxin on human cervical carcinoma cells. Int. J. Cancer 2010, 127, 1453–1461. [Google Scholar] [CrossRef]
- Holmes, S.E.; Bachran, C.; Fuchs, H.; Weng, A.; Melzig, M.F.; Flavell, S.U.; Flavell, D.J. Triterpenoid saponin augmention of saporin-based immunotoxin cytotoxicity for human leukaemia and lymphoma cells is partially immunospecific and target molecule dependent. Immunopharmacol. Immunotoxicol. 2014, 37, 42–55. [Google Scholar] [CrossRef]
- Keukens, E.A.; de Vrije, T.; van den Boom, C.; de Waard, P.; Plasman, H.H.; Thiel, F.; Chupin, V.; Jongen, W.M.; de Kruijff, B. Molecular basis of glycoalkaloid induced membrane disruption. Biochim. Biophys. Acta 1995, 1240, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Wang, R. Hemolytic mechanism of dioscin proposed by molecular dynamics simulations. J. Mol. Model. 2009, 16, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Armah, C.N.; Mackie, A.R.; Roy, C.; Price, K.; Osbourn, A.E.; Bowyer, P.; Ladha, S. The membrane-permeabilizing effect of avenacin A-1 involves the reorganization of bilayer cholesterol. Biophys. J. 1999, 76, 281–290. [Google Scholar] [CrossRef]
- Smith, W.S.; Baker, E.J.; Holmes, S.E.; Koster, G.; Hunt, A.N.; Johnston, D.A.; Flavell, S.U.; Flavell, D.J. Membrane Cholesterol is essential for Triterpenoid Saponin Augmentation of a Saporin-based Immunotoxin Directed Against CD19 on Human Lymphoma Cells. Biochim. Biophys. Acta Biomembr. 2017, 1859, 993–1007. [Google Scholar] [CrossRef]
- Wensley, J.H.; Johnston, D.A.; Smith, W.S.; Holmes, S.E.; Flavell, S.U.; Flavell, D.J. A Flow Cytometric Method to Quantify the Endosomal Escape of a protein Toxin to the Cytosol of Target Cells. Pharm. Res. 2019, 37, 16. [Google Scholar] [CrossRef] [Green Version]
- Glauert, A.M.; Dingle, J.T.; Lucy, J.A. The Action of Saponin on Biological membranes. Nature 1962, 196, 953–955. [Google Scholar] [CrossRef]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef]
- Lorent, J.H.; Leonard, C.; Abouzi, M.; Akabi, F.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. alpha-Hederin Induces Apoptosis, Membrane Permeabilization and Morphologic Changes in Two Cancer Cell Lines Through a Cholesterol-Dependent Mechanism. Planta Med. 2016, 82, 1532–1539. [Google Scholar] [CrossRef] [Green Version]
- Bottger, S.; Melzig, M.F. The influence of saponins on cell membrane cholesterol. Bioorg. Med. Chem. 2013, 21, 7118–7124. [Google Scholar] [CrossRef]
- Ikonen, E. Cellular cholesterol trafficking and compartmentalization. Nat. Rev. Mol. Cell Biol. 2008, 9, 125–138. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Subtil, A.; Gaidarov, I.; Kobylarz, K.; Lampson, M.A.; Keen, J.H.; McGraw, T.E. Acute cholesterol depletion inhibits clathrin-coated pit budding. Proc. Natl. Acad. Sci. USA 1999, 96, 6775–6780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.; Lee, S.B.; Yoo, J.H.; Nho, C.W. Flow cytometric fluorescence pulse width analysis of etoposide-induced nuclear enlargement in HCT116 cells. Biotechnol. Lett. 2010, 32, 1045–1052. [Google Scholar] [CrossRef] [Green Version]
- Kucherak, O.A.; Oncul, S.; Darwich, Z.; Yushchenko, D.A.; Arntz, Y.; Didier, P.; Mely, Y.; Klymchenko, A.S. Switchable nile red-based probe for cholesterol and lipid order at the outer leaflet of biomembranes. J. Am. Chem. Soc. 2010, 132, 4907–4916. [Google Scholar] [CrossRef]
- Gilabert-Oriol, R.; Mergel, K.; Thakur, M.; von Mallinckrodt, B.; Melzig, M.F.; Fuchs, H.; Weng, A. Real-time analysis of membrane permeabilizing effects of oleanane saponins. Bioorg. Med. Chem. 2013, 21, 2387–2395. [Google Scholar] [CrossRef] [PubMed]
- Sobo, K.; Le Blanc, I.; Luyet, P.P.; Fivaz, M.; Ferguson, C.; Parton, R.G.; Gruenberg, J.; van der Goot, F.G. Late endosomal cholesterol accumulation leads to impaired intra-endosomal trafficking. PLoS ONE 2007, 2, e851. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.S.; Johnston, D.A.; Holmes, S.E.; Wensley, H.J.; Flavell, S.U.; Flavell, D.J. Augmentation of Saporin-based Immunotoxins for Human Leukaemia and Lymphoma cells by Triterpenoid Saponins: The Modifying Effects of Small Molecule Pharmacological Agents. Toxins 2019, 11, 127. [Google Scholar] [CrossRef] [Green Version]
- Baravalle, G.; Schober, D.; Huber, M.; Bayer, N.; Murphy, R.F.; Fuchs, R. Transferrin recycling and dextran transport to lysosomes is differentially affected by bafilomycin, nocodazole, and low temperature. Cell Tissue Res. 2005, 320, 99–113. [Google Scholar] [CrossRef]
- Wang, S.; Sun, H.; Tanowitz, M.; Liang, X.H.; Crooke, S.T. Intra-endosomal trafficking mediated by lysobisphosphatidic acid contributes to intracellular release of phosphorothioate-modified antisense oligonucleotides. Nucleic Acids Res. 2017, 45, 5309–5322. [Google Scholar] [CrossRef]
- Bissig, C.; Gruenberg, J. Lipid sorting and multivesicular endosome biogenesis. Cold Spring Harb. Perspect. Biol. 2013, 5, a016816. [Google Scholar] [CrossRef] [Green Version]
- Góral, I.; Jurek, I.; Wojciechowski, K. How Does the Surface Activity of Soapwort (Saponaria officinalis L.) Extracts Depend on the Plant Organ? J. Surfactants Deterg. 2018, 21, 797–807. [Google Scholar] [CrossRef]
- Orczyk, M.; Wojciechowski, K.; Brezesinski, G. Disordering Effects of Digitonin on Phospholipid Monolayers. Langmuir 2017, 33, 3871–3881. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, K.; Orczyk, M.; Gutberlet, T.; Trapp, M.; Marcinkowski, K.; Kobiela, T.; Geue, T. Unusual penetration of phospholipid mono- and bilayers by Quillaja bark saponin biosurfactant. Biochim. Biophys. Acta 2014, 1838, 1931–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Konoki, K.; Tachibana, K. Cholesterol-independent membrane disruption caused by triterpenoid saponins. Biochim. Biophys. Acta 1996, 1299, 252–258. [Google Scholar] [CrossRef]
- Verstraeten, S.L.; Deleu, M.; Janikowska-Sagan, M.; Claereboudt, E.J.S.; Lins, L.; Tyteca, D.; Mingeot-Leclercq, M.P. The activity of the saponin ginsenoside Rh2 is enhanced by the interaction with membrane sphingomyelin but depressed by cholesterol. Sci. Rep. 2019, 9, 7285. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, S.L.; Albert, M.; Paquot, A.; Muccioli, G.G.; Tyteca, D.; Mingeot-Leclercq, M.P. Membrane cholesterol delays cellular apoptosis induced by ginsenoside Rh2, a steroid saponin. Toxicol. Appl. Pharmacol. 2018, 352, 59–67. [Google Scholar] [CrossRef]
- Attard, G.S.; Templer, R.H.; Smith, W.S.; Hunt, A.N.; Jackowski, S. Modulation of CTP: Phosphocholine cytidylyltransferase by membrane curvature elastic stress. Proc. Natl. Acad. Sci. USA 2000, 97, 9032–9036. [Google Scholar] [CrossRef] [Green Version]
- Dymond, M.; Attard, G.; Postle, A.D. Testing the hypothesis that amphiphilic antineoplastic lipid analogues act through reduction of membrane curvature elastic stress. J. R. Soc. Interface 2008, 5, 1371–1386. [Google Scholar] [CrossRef] [Green Version]
- Shearman, G.C.; Attard, G.S.; Hunt, A.N.; Jackowski, S.; Baciu, M.; Sebai, S.C.; Mulet, X.; Clarke, J.A.; Law, R.V.; Plisson, C.; et al. Using membrane stress to our advantage. Biochem. Soc. Trans. 2007, 35, 498–501. [Google Scholar] [CrossRef] [Green Version]
- Weng, A.; Thakur, M.; Schindler, A.; Fuchs, H.; Melzig, M.F. Liquid-chromatographic profiling of Saponinum album (Merck). Pharmazie 2011, 66, 744–746. [Google Scholar]
- Flavell, D.J.; Flavell, S.U.; Boehm, D.; Emery, L.; Noss, A.; Ling, N.R.; Richardson, P.R.; Hardie, D.; Wright, D.H. Preclinical studies with the anti-CD19-saporin immunotoxin BU12-SAPORIN for the Treatment of Human B-Cell Tumours. Br. J. Cancer 1995, 72, 1373–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprong, H.; Suchanek, M.; van Dijk, S.M.; van Remoortere, A.; Klumperman, J.; Avram, D.; van der Linden, J.; Leusen, J.H.; van Hellemond, J.J.; Thiele, C. Aberrant receptor-mediated endocytosis of Schistosoma mansoni glycoproteins on host lipoproteins. PLoS Med. 2006, 3, e253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, A.E.; Haynes, M.P.; Phillips, M.C.; Rothblat, G.H. Use of cyclodextrins for manipulating cellular cholesterol content. J. Lipid Res. 1997, 38, 2264–2272. [Google Scholar] [PubMed]
- Hinzey, A.H.; Kline, M.A.; Kotha, S.R.; Sliman, S.M.; Butler, E.S.; Shelton, A.B.; Gurney, T.R.; Parinandi, N.L. Choice of cyclodextrin for cellular cholesterol depletion for vascular endothelial cell lipid raft studies: Cell membrane alterations, cytoskeletal reorganization and cytotoxicity. Indian J. Biochem. Biophys. 2012, 49, 329–341. [Google Scholar] [PubMed]
- Tobert, J.A. Lovastatin and beyond: The history of the HMG-CoA reductase inhibitors. Nat. Rev. Drug Discov. 2003, 2, 517–526. [Google Scholar] [CrossRef]
- Darwich, Z.; Klymchenko, A.S.; Kucherak, O.A.; Richert, L.; Mely, Y. Detection of apoptosis through the lipid order of the outer plasma membrane leaflet. Biochim. Biophys. Acta 2012, 1818, 3048–3054. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.S.; Flavell, S.U.; Flavell, D.J. The Fluorescent Probe NR12S Detects Changes in Plasma Membrane Cholesterol Levels. BMG LabTech 2017. Available online: https://www.bmglabtech.com/the-fluorescent-probe-nr12s-detects-changes-in-plasma-membrane-cholesterol-levels/ (accessed on 19 November 2020).
- Scudiero, D.A.; Shoemaker, R.H.; Paull, K.D.; Monks, A.; Tierney, S.; Nofziger, T.H.; Currens, M.J.; Seniff, D.; Boyd, M.R. Evaluation of a soluble tetrazolium/formazan assay for cell growth and drug sensitivity in culture using human and other tumor cell lines. Cancer Res. 1988, 48, 4827–4833. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, W.S.; Johnston, D.A.; Wensley, H.J.; Holmes, S.E.; Flavell, S.U.; Flavell, D.J. The Role of Cholesterol on Triterpenoid Saponin-Induced Endolysosomal Escape of a Saporin-Based Immunotoxin. Int. J. Mol. Sci. 2020, 21, 8734. https://doi.org/10.3390/ijms21228734
Smith WS, Johnston DA, Wensley HJ, Holmes SE, Flavell SU, Flavell DJ. The Role of Cholesterol on Triterpenoid Saponin-Induced Endolysosomal Escape of a Saporin-Based Immunotoxin. International Journal of Molecular Sciences. 2020; 21(22):8734. https://doi.org/10.3390/ijms21228734
Chicago/Turabian StyleSmith, Wendy S., David A. Johnston, Harrison J. Wensley, Suzanne E. Holmes, Sopsamorn U. Flavell, and David J. Flavell. 2020. "The Role of Cholesterol on Triterpenoid Saponin-Induced Endolysosomal Escape of a Saporin-Based Immunotoxin" International Journal of Molecular Sciences 21, no. 22: 8734. https://doi.org/10.3390/ijms21228734
APA StyleSmith, W. S., Johnston, D. A., Wensley, H. J., Holmes, S. E., Flavell, S. U., & Flavell, D. J. (2020). The Role of Cholesterol on Triterpenoid Saponin-Induced Endolysosomal Escape of a Saporin-Based Immunotoxin. International Journal of Molecular Sciences, 21(22), 8734. https://doi.org/10.3390/ijms21228734