Strand Displacement Activity of PrimPol


 and
and
Abstract
:1. Introduction
2. Results
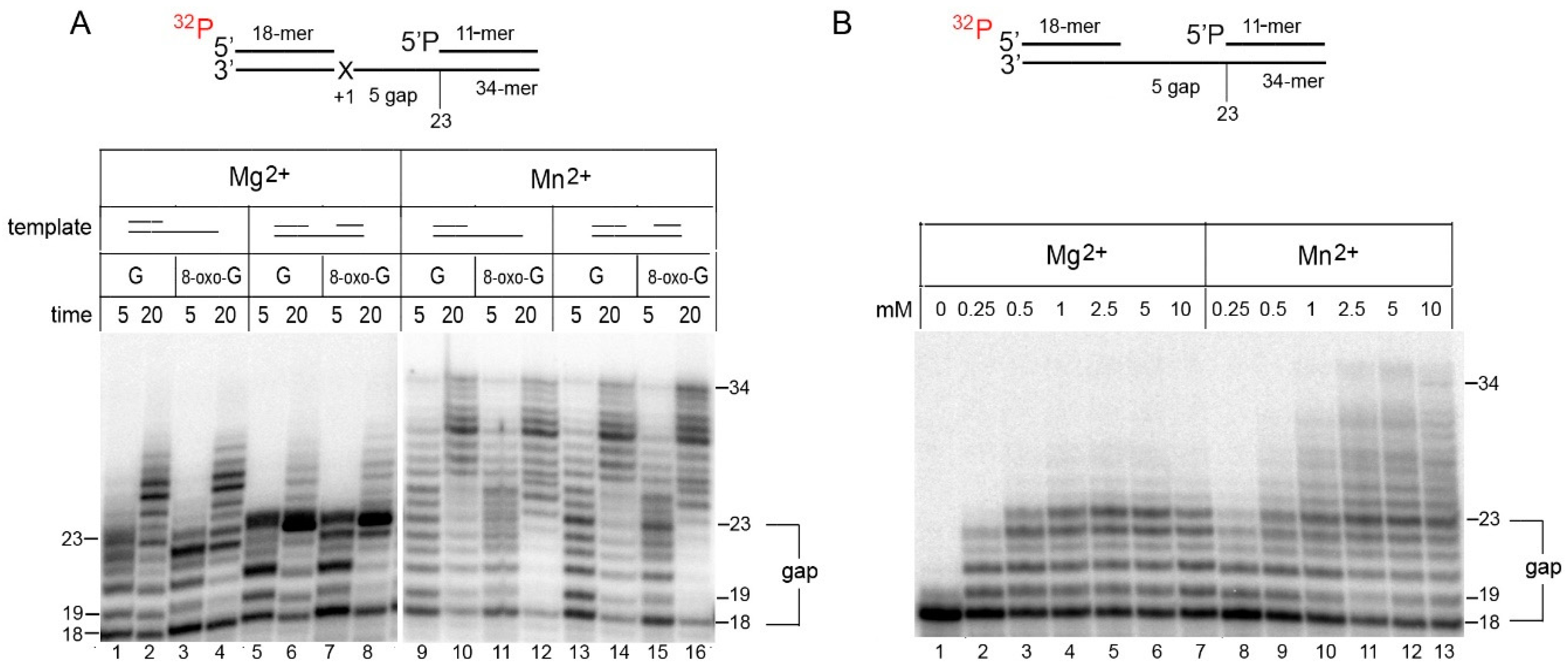
2.1. Mn2+ Ions Stimulate the DNA Strand Displacement Activity of PrimPol
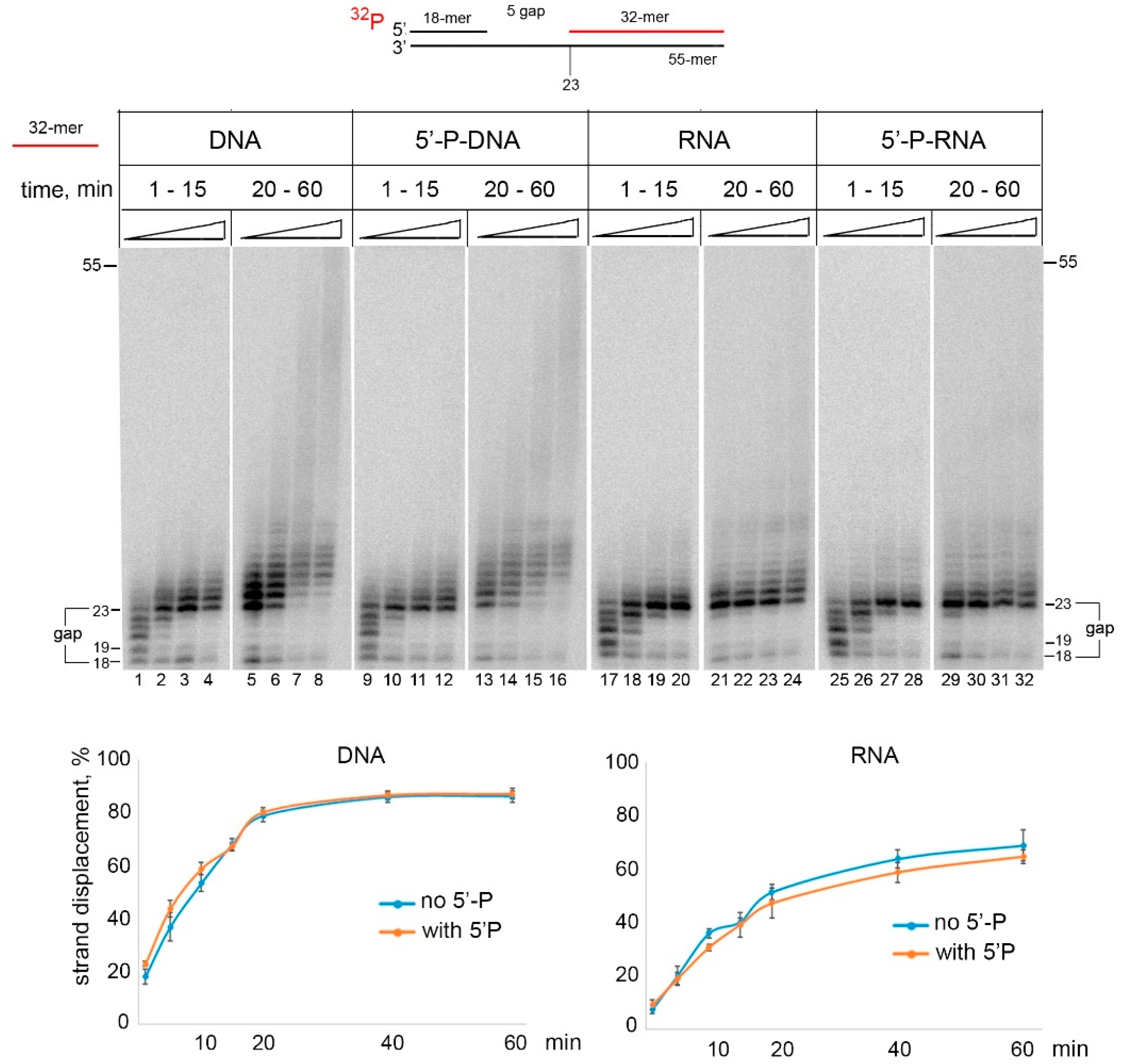
2.2. RNA Strand Displacement Activity of PrimPol Is Not Efficient
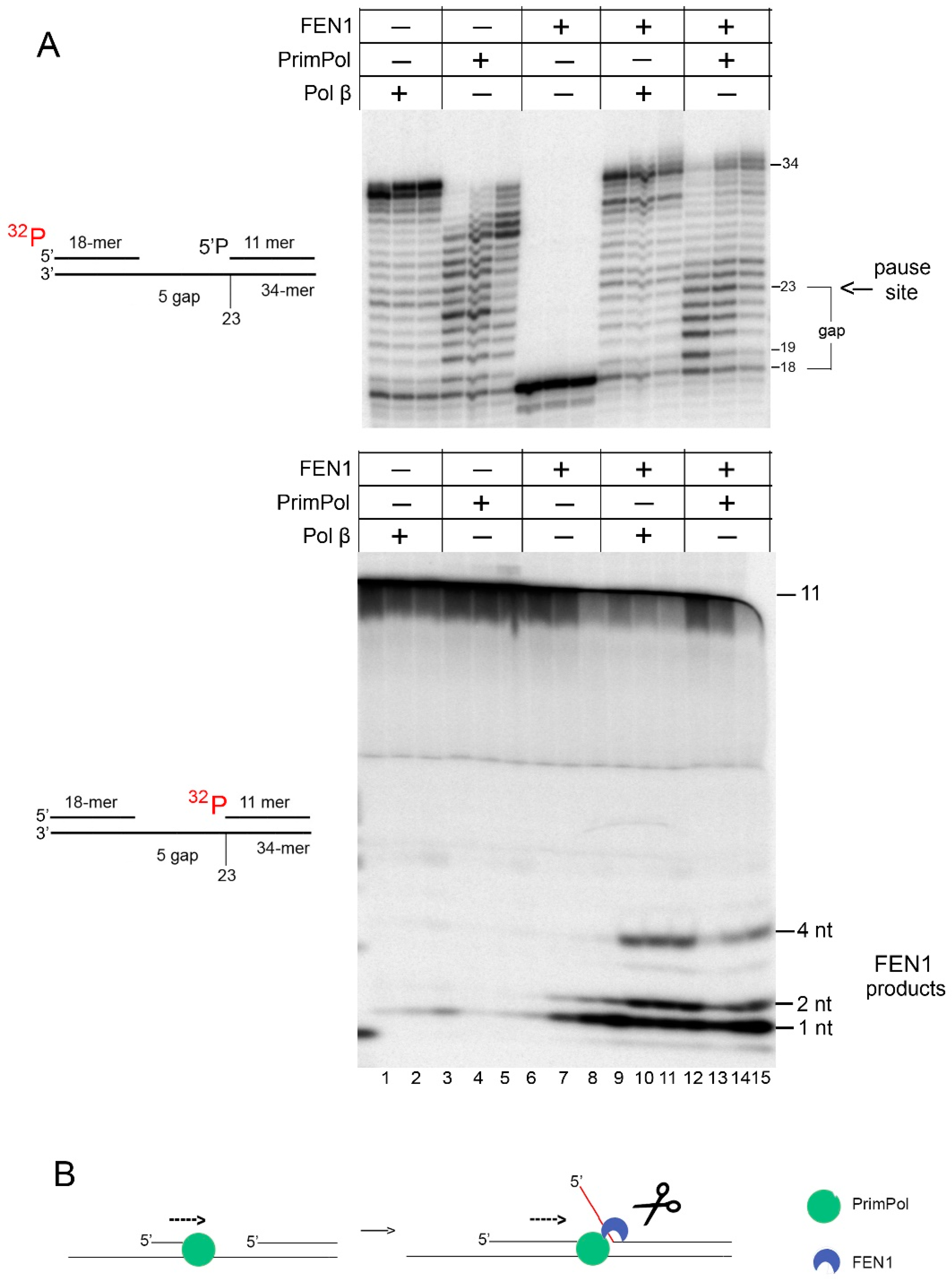
2.3. FEN1 Processes the 5′-Flaps Generated by PrimPol In Vitro
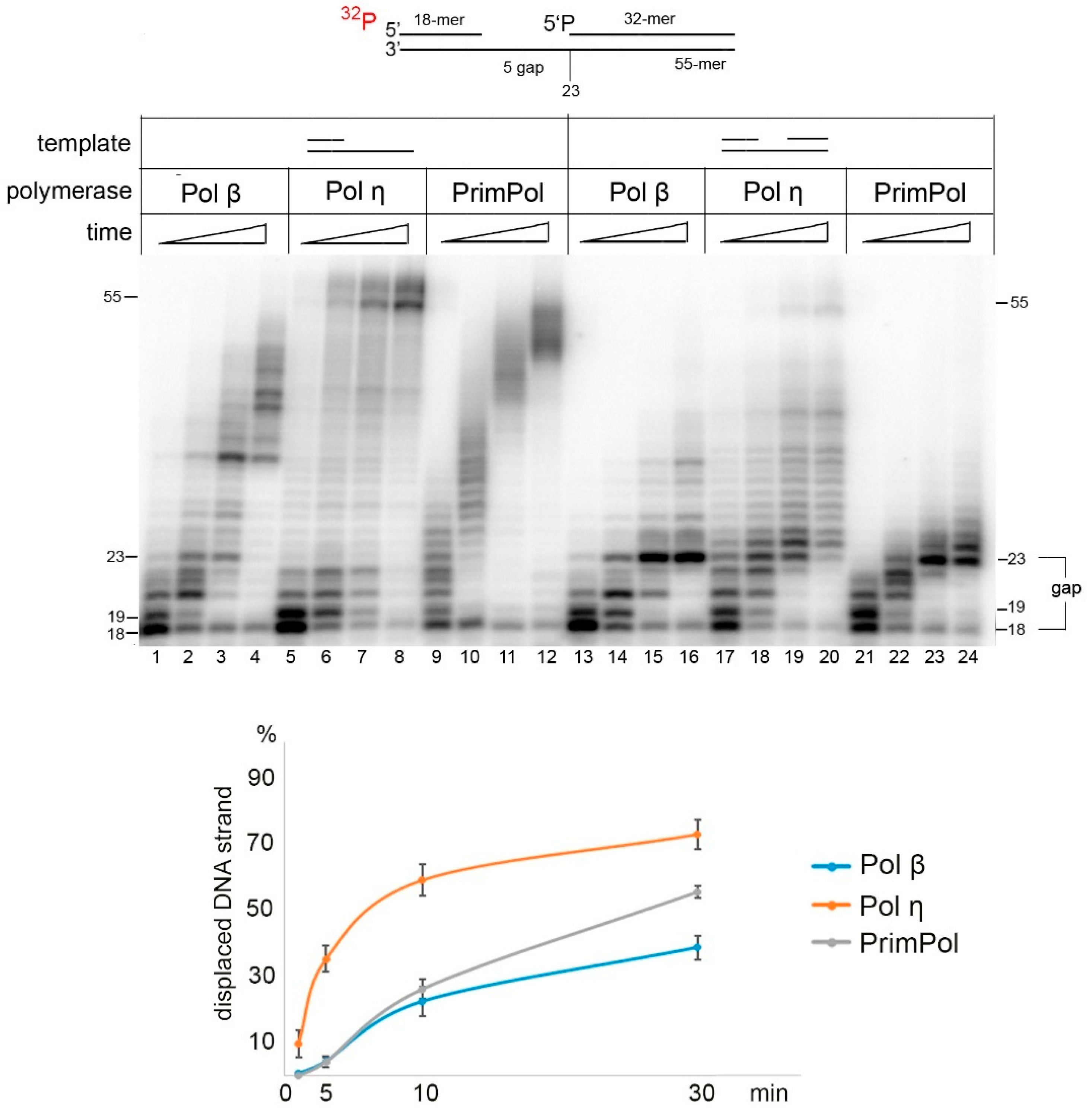
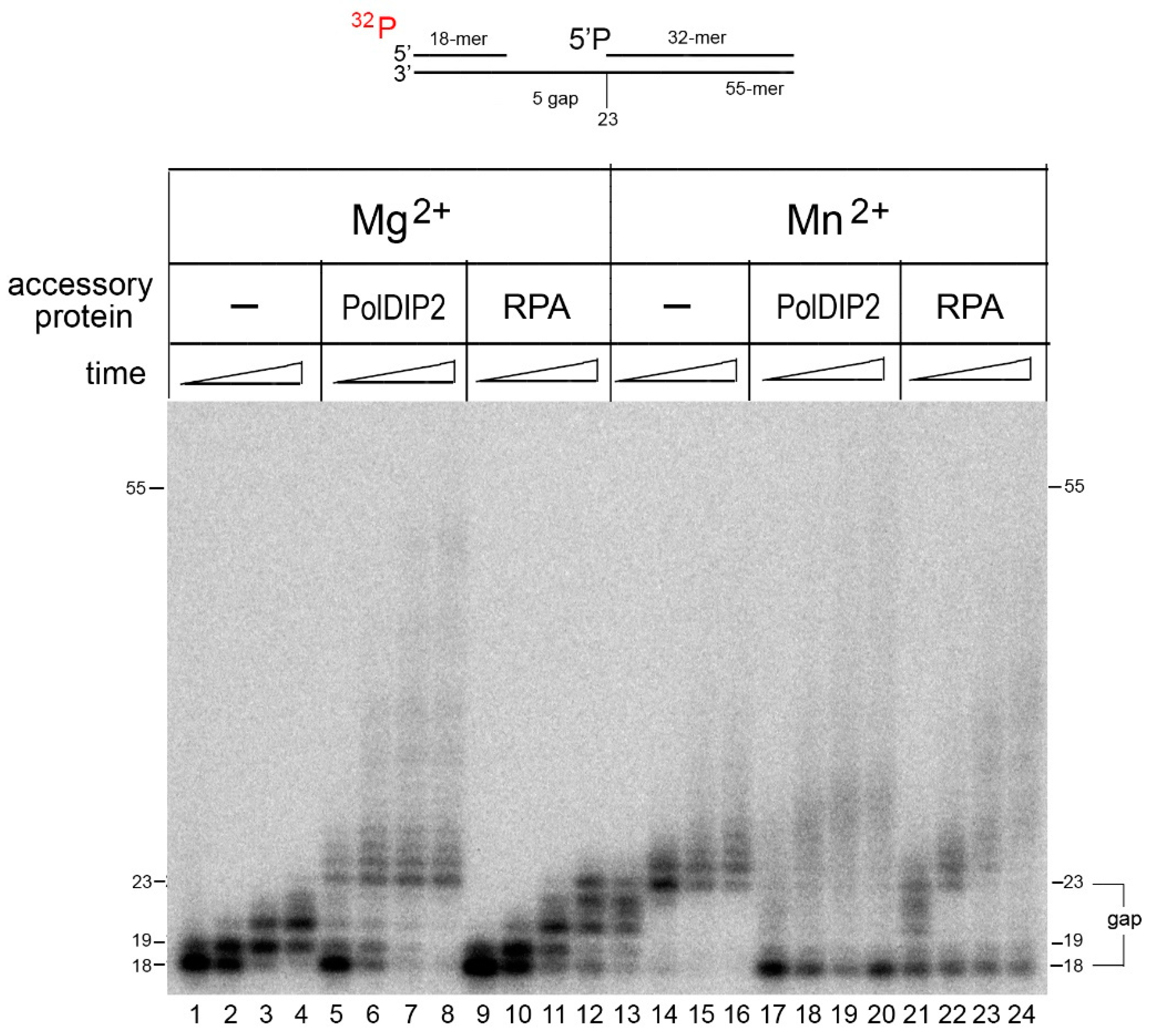
2.4. The Strand Displacement Activity of PrimPol Is Stimulated by PolDIP2 and RPA
3. Discussion
4. Materials and Methods
4.1. Proteins
4.2. DNA Substrates
4.3. Primer Extension Reactions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ssDNA | Single-stranded DNA |
| 8-oxo-G | 8-oxoguanine |
References
- Bianchi, J.J.; Rudd, S.G.; Jozwiakowski, S.K.; Bailey, L.J.; Soura, V.; Taylor, E.; Stevanovic, I.; Green, A.J.; Stracker, T.H.; Lindsay, H.D.; et al. PrimPol bypasses UV photoproducts during eukaryotic chromosomal DNA replication. Mol. Cell 2013, 52, 566–573. [Google Scholar] [CrossRef] [Green Version]
- García-Gómez, S.; Reyes, A.; Martínez-Jiménez, M.I.; Chocrón, S.; Mourón, S.; Terrados, G.; Powell, C.; Salido, E.; Méndez, J.; Holt, I.J.; et al. PrimPol, an archaic primase/polymerase operating in human cells. Mol. Cell 2013, 52, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Jiménez, M.I.; García-Gómez, S.; Bebenek, K.; Sastre-Moreno, G.; Calvo, P.A.; Díaz-Talavera, A.; Kunkel, T.A.; Blanco, L. Alternative solutions and new scenarios for translesion DNA synthesis by human PrimPol. DNA Repair 2015, 29, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Guilliam, T.A.; Tsuda, M.; Yamamoto, J.; Bailey, L.J.; Iwai, S.; Takeda, S.; Doherty, A.J.; Hirota, K. Repriming by PrimPol is critical for DNA replication restart downstream of lesions and chain-terminating nucleosides. Cell Cycle 2016, 15, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- Mourón, S.; Rodriguez-Acebes, S.; Martínez-Jiménez, M.I.; García-Gómez, S.; Chocrón, S.; Blanco, L.; Méndez, J. Repriming of DNA synthesis at stalled replication forks by human PrimPol. Struct. Mol. Biol. 2013, 20, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavone, D.; Jozwiakowski, S.K.; Romanello, M.; Guilbaud, G.; Guilliam, T.A.; Bailey, L.J.; Sale, J.E.; Doherty, A.J. PrimPol Is Required for Replicative Tolerance of G Quadruplexes in Vertebrate Cells. Mol. Cell 2016, 61, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Muñumer, R.; Forslund, J.M.E.; Goffart, S.; Pfeiffer, A.; Stojkovič, G.; Carvalho, G.; Al-Furoukh, N.; Blanco, L.; Wanrooij, S.; Pohjoismäki, J.L.O. PrimPol is required for replication reinitiation after mtDNA damage. Proc. Natl. Acad. Sci. USA 2017, 114, 11398–11403. [Google Scholar] [CrossRef] [Green Version]
- Makarova, A.V.; Boldinova, E.O.; Belousova, E.A.; Lavrik, O.I. In Vitro lesion bypass by human PrimPol. DNA Repair 2018, 70, 18–24. [Google Scholar] [CrossRef]
- Maga, G.; Villani, G.; Tillement, V.; Stucki, M.; Locatelli, G.A.; Frouin, I.; Spadari, S.; Hübscher, U. Okazaki fragment processing: Modulation of the strand displacement activity of DNA polymerase delta by the concerted action of replication protein A, proliferating cell nuclear antigen, and flap endonuclease-1. Proc. Natl. Acad. Sci. USA 2001, 98, 14298–14303. [Google Scholar] [CrossRef] [Green Version]
- Sebesta, M.; Burkovics, P.; Juhasz, S.; Zhang, S.; Szabo, J.E.; Lee, M.Y.W.T.; Haracska, L.; Krejci, L. Role of PCNA and TLS polymerases in D-loop extension during homologous recombination in humans. DNA Repair 2013, 12, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Lebedeva, N.A.; Rechkunova, N.I.; Dezhurov, S.V.; Khodyreva, S.N.; Favre, A.; Blanco, L.; Lavrik, O.I. Comparison of functional properties of mammalian DNA polymerase λ and DNA polymerase β in reactions of DNA synthesis related to DNA repair. Biochim. Biophys. Acta 2005, 1751, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Maga, G.; van Loon, B.; Crespan, E.; Villani, G.; Hübscher, U. The Block of DNA Polymerase δ Strand Displacement Activity by an Abasic Site Can Be Rescued by the Concerted Action of DNA Polymerase β and Flap Endonuclease 1. J. Biol. Chem. 2009, 284, 14267–14275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.; Dianov, G.L.; Bohr, V.A.; Wilson, S.H. FEN1 stimulation of DNA polymerase beta mediates an excision step in mammalian long patch base excision repair. J. Biol. Chem. 2000, 275, 4460–4466. [Google Scholar] [CrossRef] [Green Version]
- Ganai, R.A.; Zhang, X.-P.; Heyer, W.-D.; Johansson, E. Strand displacement synthesis by yeast DNA polymerase ε. Nucleic Acids Res. 2016, 44, 8229–8240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.V.; Guainazzi, A.; Derkunt, S.B.; Enoiu, M.; Schärer, O.D. Structure-dependent bypass of DNA interstrand crosslinks by translesion synthesis polymerases. Nucleic Acids Res. 2011, 39, 7455–7464. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Holzschu, D.L.; Sugiyama, T. PCNA is efficiently loaded on the DNA recombination intermediate to modulate polymerase δ, η, and ζ activities. Proc. Natl. Acad. Sci. USA 2013, 110, 7672–7677. [Google Scholar] [CrossRef] [Green Version]
- McIlwraith, M.J.; Vaisman, A.; Liu, Y.; Fanning, E.; Woodgate, R.; West, S.C. Human DNA polymerase eta promotes DNA synthesis from strand invasion intermediates of homologous recombination. Mol. Cell 2005, 20, 783–792. [Google Scholar] [CrossRef]
- He, Q.; Shumate, C.K.; White, M.A.; Molineux, I.J.; Yin, Y.W. Exonuclease of human DNA polymerase gamma disengages its strand displacement function. Mitochondrion 2013, 13, 592–601. [Google Scholar] [CrossRef] [Green Version]
- Podust, V.N.; Hübscher, U. Lagging strand DNA synthesis by calf thymus DNA polymerases alpha, beta, delta and epsilon in the presence of auxiliary proteins. Nucleic Acids Res. 1993, 21, 841–846. [Google Scholar] [CrossRef]
- Giannattasio, M.; Branzei, D. DNA Replication Through Strand Displacement During Lagging Strand DNA Synthesis in Saccharomyces cerevisiae. Genes 2019, 10, 167. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.A.; Pack, L.R.; Sanman, L.E.; Suo, Z. Efficiency and fidelity of human DNA polymerases λ and β during gap-filling DNA synthesis. DNA Repair 2011, 10, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mislak, A.C.; Anderson, K.S. Insights into the Molecular Mechanism of Polymerization and Nucleoside Reverse Transcriptase Inhibitor Incorporation by Human PrimPol. Agents Chemother. 2015, 60, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, M.K.; Ketkar, A.; Lodeiro, M.F.; Cameron, C.E.; Eoff, R.L. Kinetic analysis of human PrimPol DNA polymerase activity reveals a generally error-prone enzyme capable of accurately bypassing 7,8-dihydro-8-oxo-2′-deoxyguanosine. Biochemistry 2014, 53, 6584–6594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, J.J.; Lieber, M.R. The characterization of a mammalian DNA structure-specific endonuclease. EMBO J. 1994, 13, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Lavrik, O.; Kim, S.J.; Kedar, P.; Yang, X.P.; Vande Berg, B.J.; Wilson, S.H. DNA polymerase beta -mediated long patch base excision repair. Poly(ADP-ribose)polymerase-1 stimulates strand displacement DNA synthesis. J. Biol. Chem. 2001, 276, 32411–32414. [Google Scholar] [CrossRef] [Green Version]
- Stodola, J.L.; Burgers, P.M. Resolving individual steps of Okazaki-fragment maturation at a millisecond timescale. Nat. Struct. Mol. Biol. 2016, 23, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Tom, S.; Henricksen, L.A.; Bambara, R.A. Mechanism whereby proliferating cell nuclear antigen stimulates flap endonuclease 1. J. Biol. Chem. 2000, 275, 10498–10505. [Google Scholar] [CrossRef] [Green Version]
- Guilliam, T.A.; Bailey, L.J.; Brissett, N.C.; Doherty, A.J. PolDIP2 interacts with human PrimPol and enhances its DNA polymerase activities. Nucleic Acids Res. 2016, 44, 3317–3329. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Jiménez, M.I.; Lahera, A.; Blanco, L. Human PrimPol activity is enhanced by RPA. Sci. Rep. 2017, 7, 783. [Google Scholar] [CrossRef] [Green Version]
- Guilliam, T.A.; Brissett, N.C.; Ehlinger, A.; Keen, B.A.; Kolesar, P.; Taylor, E.M.; Bailey, L.J.; Lindsay, H.D.; Chazin, W.J.; Doherty, A.J. Molecular basis for PrimPol recruitment to replication forks by RPA. Nat. Commun. 2017, 8, 15222. [Google Scholar] [CrossRef]
- Gagarinskaya, D.I.; Makarova, A.V. A multifunctional protein PolDIP2 in DNA translesion synthesis. Adv. Exp. Med. Biol. 2020, 1241, 35–45. [Google Scholar] [PubMed]
- Wan, L.; Lou, J.; Xia, Y.; Su, B.; Liu, T.; Cui, J.; Sun, Y.; Lou, H.; Huang, J. hPrimpol1/CCDC111 is a human DNA primase-polymerase required for the maintenance of genome integrity. EMBO Rep. 2013, 14, 1104–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maréchal, A.; Zou, L. RPA-coated single-stranded DNA as a platform for post-translational modifications in the DNA damage response. Cell Res. 2015, 25, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, L.J.; Borowiec, J.A. Human replication protein A binds single-stranded DNA in two distinct complexes. Mol. Cell Biol. 1994, 14, 3993–4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochkarev, A.; Pfuetzner, R.A.; Edwards, A.M.; Frappier, L. Structure of the single-stranded-DNA-binding domain of replication protein A bound to DNA. Nature 1997, 385, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Krasikova, Y.S.; Rechkunova, N.I.; Maltseva, E.A.; Lavrik, O.I. RPA and XPA interaction with DNA structures mimicking intermediates of the late stages in nucleotide excision repair. PLoS ONE 2018, 13, e0190782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavrik, O.I.; Kolpashchikov, D.M.; Weisshart, K.; Nasheuer, H.P.; Khodyreva, S.N.; Favre, A. RPA subunit arrangement near the 3′-end of the primer is modulated by the length of the template strand and cooperative protein interactions. Nucleic Acids Res. 1999, 27, 4235–4240. [Google Scholar] [CrossRef]
- Liu, P.; Qian, L.; Sung, J.S.; de Souza-Pinto, N.C.; Zheng, L.; Bogenhagen, D.F.; Bohr, V.A.; Wilson, D.M., 3rd; Shen, B.; Demple, B. Removal of Oxidative DNA Damage via FEN1-Dependent Long-Patch Base Excision Repair in Human Cell Mitochondria. Mol. Cell Biol. 2008, 28, 4975–4987. [Google Scholar] [CrossRef] [Green Version]
- Rudd, S.G. Cellular and Biochemical Characterisation of PrimPol, a Novel Eukaryotic Primase-Polymerase Involved in DNA Damage Tolerance. Ph.D. Thesis, University of Sussex, Brighton, UK, 2013. [Google Scholar]
- Boldinova, E.O.; Stojkovič, G.; Khairullin, R.; Wanrooij, S.; Makarova, A.V. Optimization of the expression, purification and polymerase activity reaction conditions of recombinant human PrimPol. PLoS ONE 2017, 12, e0184489. [Google Scholar] [CrossRef] [Green Version]
- Binz, S.K.; Dickson, A.M.; Haring, S.J.; Wold, M.S. Functional assays for replication protein A (RPA). Methods Enzymol. 2006, 409, 11–38. [Google Scholar]
- Nazarkina, J.K.; Petrousseva, I.O.; Safronov, I.V.; Lavrik, O.I.; Khodyreva, S.N. Interaction of Flap Endonuclease-1 and Replication Protein A with Photoreactive Intermediates of DNA Repair. Biochemistry (Mosc) 2003, 68, 934–942. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | 5′-CGGTATCCACCAGGTCTG-3′ |
|---|---|
| 34-mer DNA template | 5′-GGCTTCATCGTTGTCXCAGACCTGGTGGATACCG-3′ X = G or 8-oxo-G |
| 55-mer DNA template | 5′-GACTACATTTCATCTGGCTTGGGCTTCATCGTTGTCGCAGACCTGGTGGATACCG-3′ |
| Locking primer 1 (11-mer 5′-P-DNA) | 5′-(P)ACGATGAAGCC-3′(P)—phosphate |
| Locking primer 2 (11-mer DNA) | 5′-ACGATGAAGCC-3′ |
| Locking primer 3 (32-mer 5′-P-DNA) | 5′-(P)ACGATGAAGCCCAAGCCAGATGAAATGTAGTC-3′ (P)—phosphate |
| Locking primer 4 (32-mer DNA) | 5′-ACGATGAAGCCCAAGCCAGATGAAATGTAGTC-3′ |
| Locking primer 5 (32-mer 5′-P-RNA) | 5′-(P)ACGAUGAAGCCCAAGCCAGAUGAAAUGUAGUC-3′ (P)—phosphate |
| Locking primer 6 (32-mer RNA) | 5′- ACGAUGAAGCCCAAGCCAGAUGAAAUGUAGUC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boldinova, E.O.; Belousova, E.A.; Gagarinskaya, D.I.; Maltseva, E.A.; Khodyreva, S.N.; Lavrik, O.I.; Makarova, A.V. Strand Displacement Activity of PrimPol. Int. J. Mol. Sci. 2020, 21, 9027. https://doi.org/10.3390/ijms21239027
Boldinova EO, Belousova EA, Gagarinskaya DI, Maltseva EA, Khodyreva SN, Lavrik OI, Makarova AV. Strand Displacement Activity of PrimPol. International Journal of Molecular Sciences. 2020; 21(23):9027. https://doi.org/10.3390/ijms21239027
Chicago/Turabian StyleBoldinova, Elizaveta O., Ekaterina A. Belousova, Diana I. Gagarinskaya, Ekaterina A. Maltseva, Svetlana N. Khodyreva, Olga I. Lavrik, and Alena V. Makarova. 2020. "Strand Displacement Activity of PrimPol" International Journal of Molecular Sciences 21, no. 23: 9027. https://doi.org/10.3390/ijms21239027
APA StyleBoldinova, E. O., Belousova, E. A., Gagarinskaya, D. I., Maltseva, E. A., Khodyreva, S. N., Lavrik, O. I., & Makarova, A. V. (2020). Strand Displacement Activity of PrimPol. International Journal of Molecular Sciences, 21(23), 9027. https://doi.org/10.3390/ijms21239027