Syringic Acid Alleviates Cesium-Induced Growth Defect in Arabidopsis

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
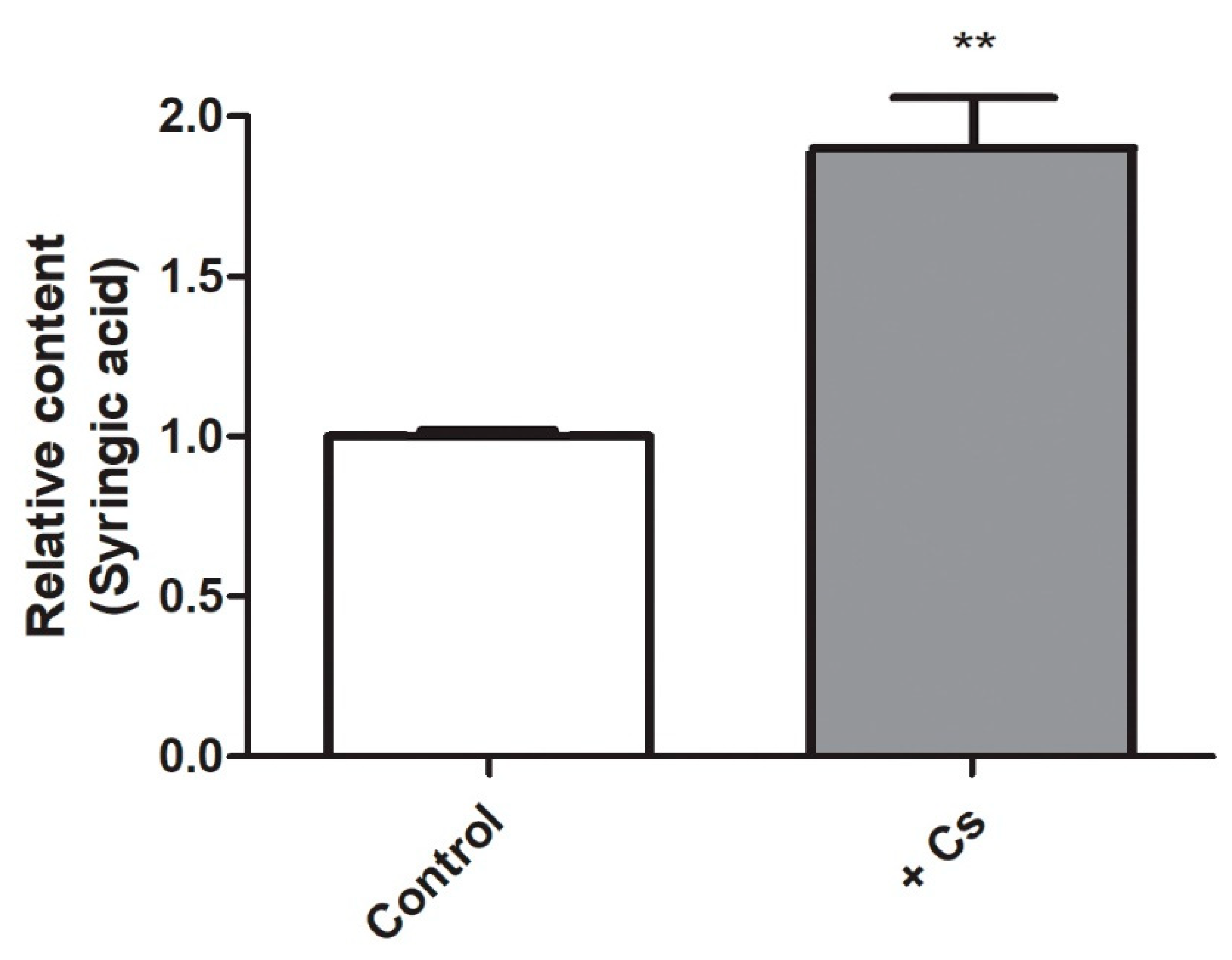
2.1. Increased Levels of Syringic Acid in Cesium-Treated Arabidopsis Shoots
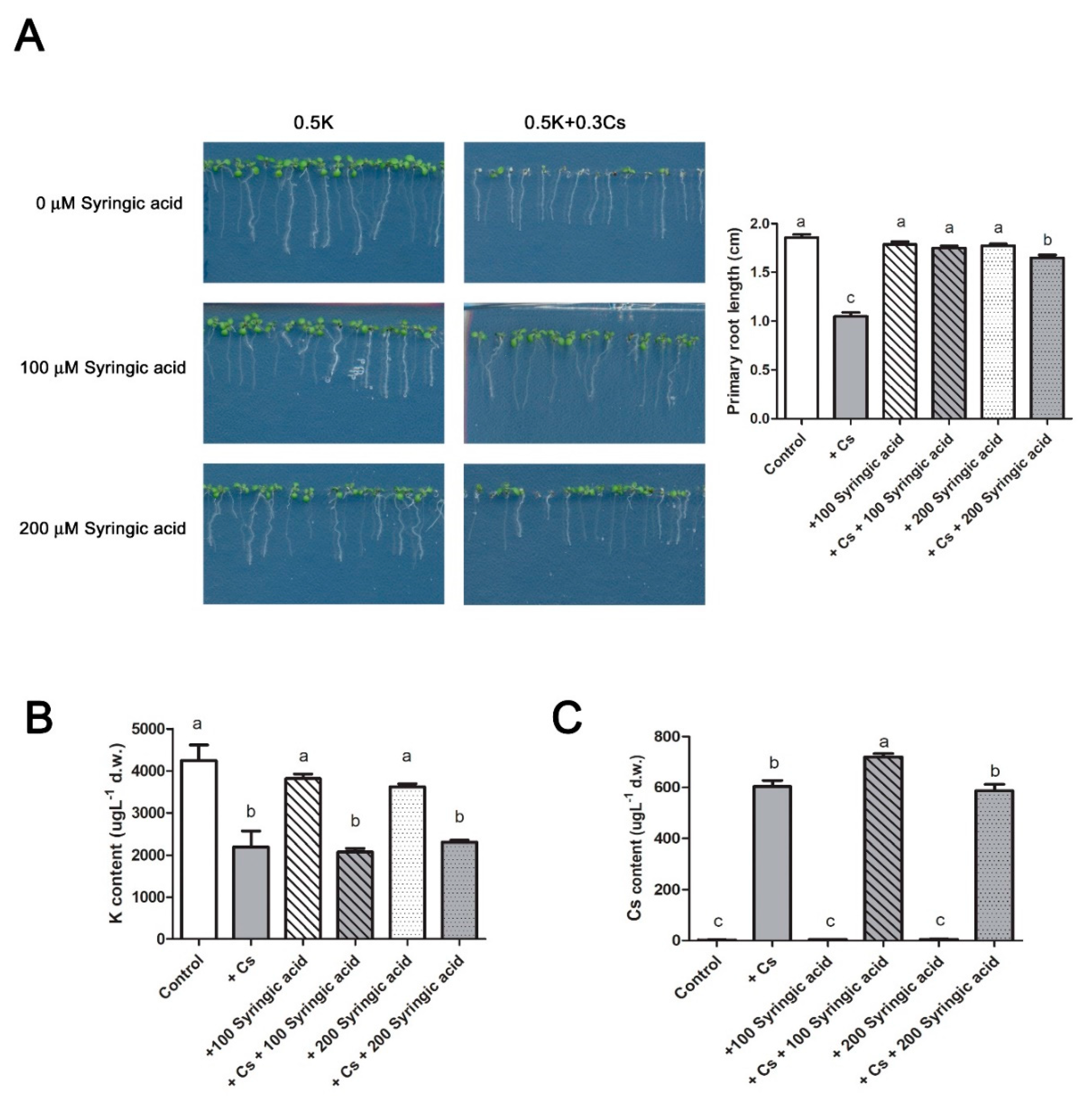
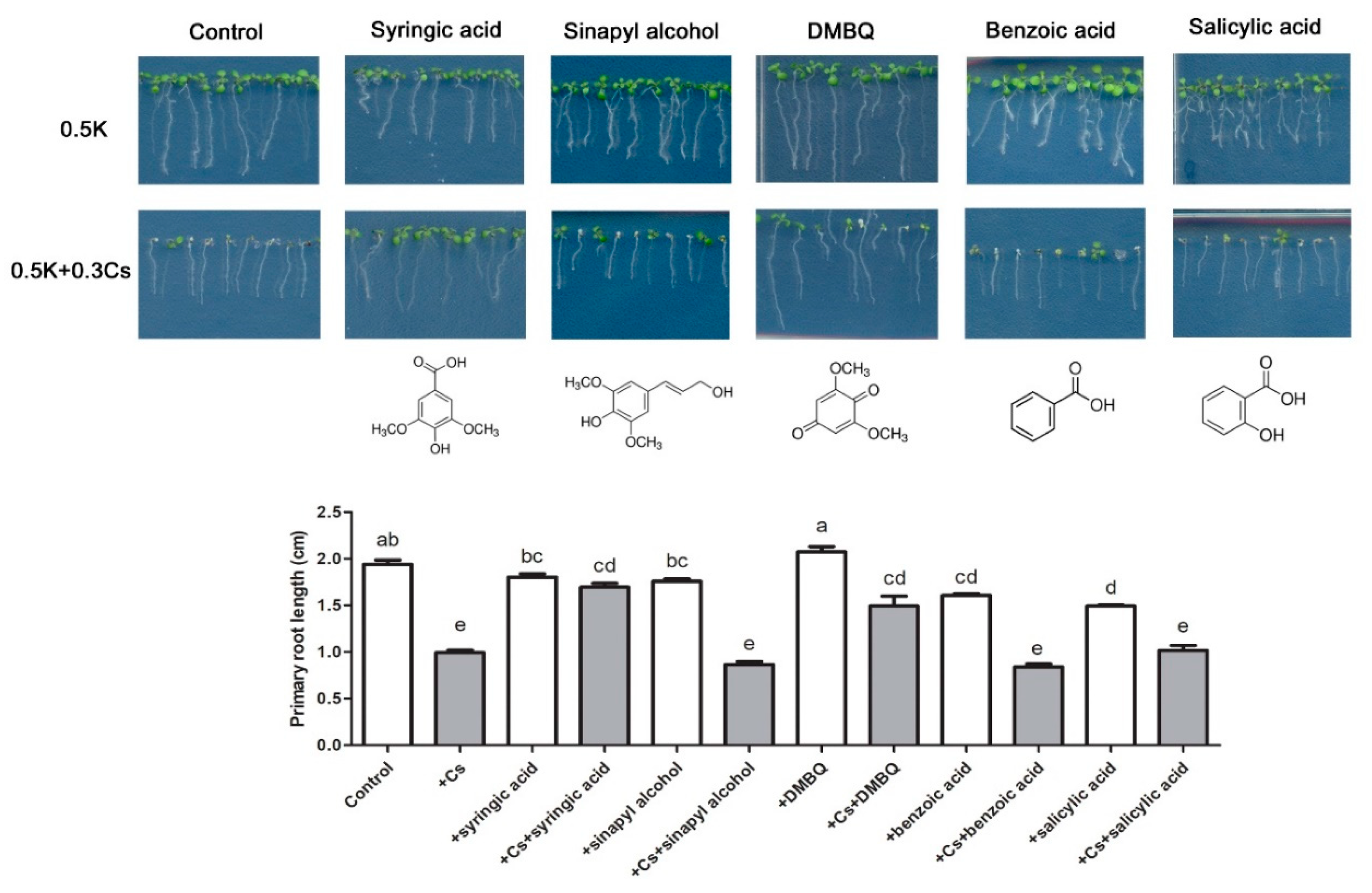
2.2. Syringic Acid Treatment Alleviates Cesium-Induced Growth Inhibition
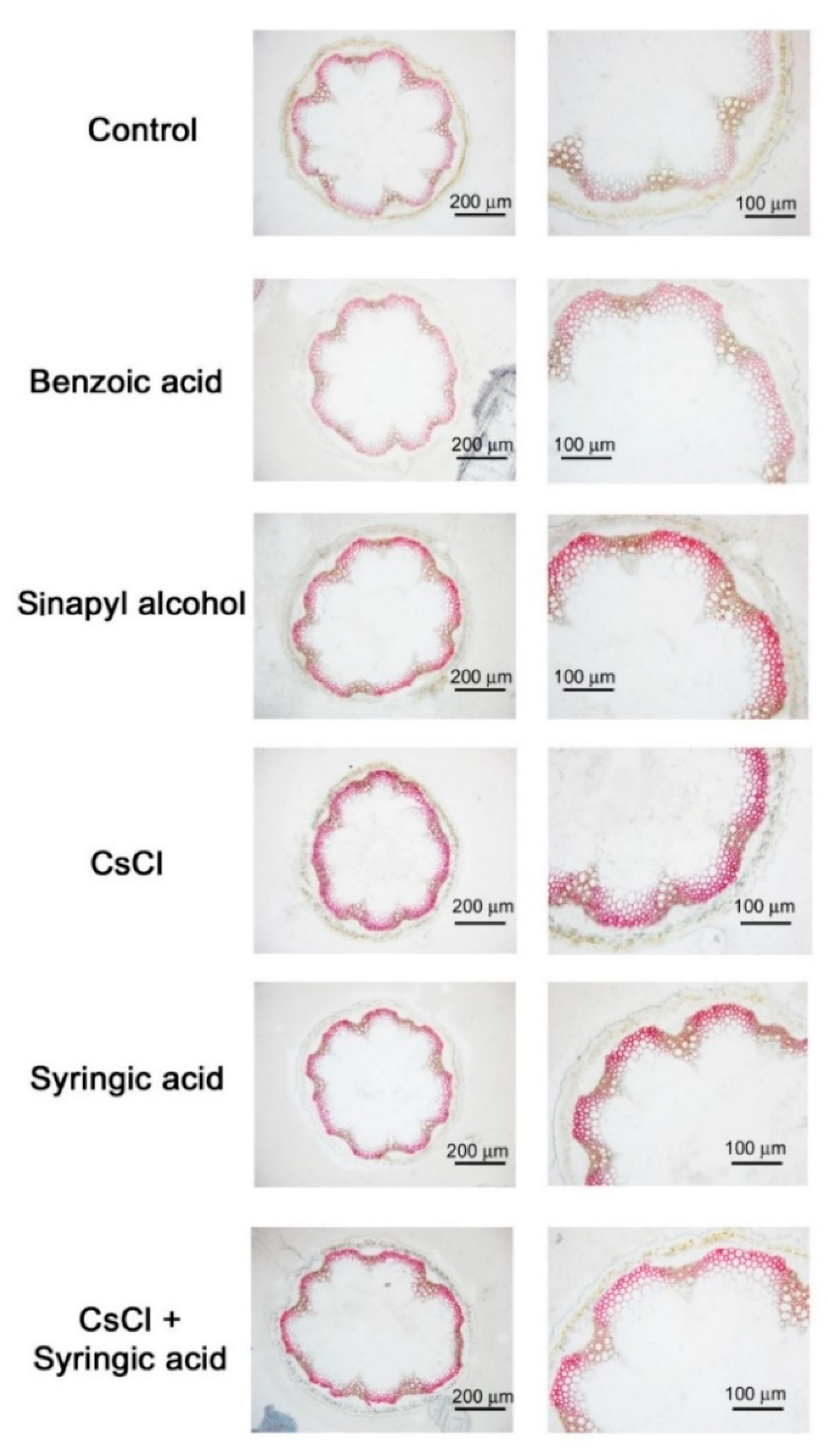
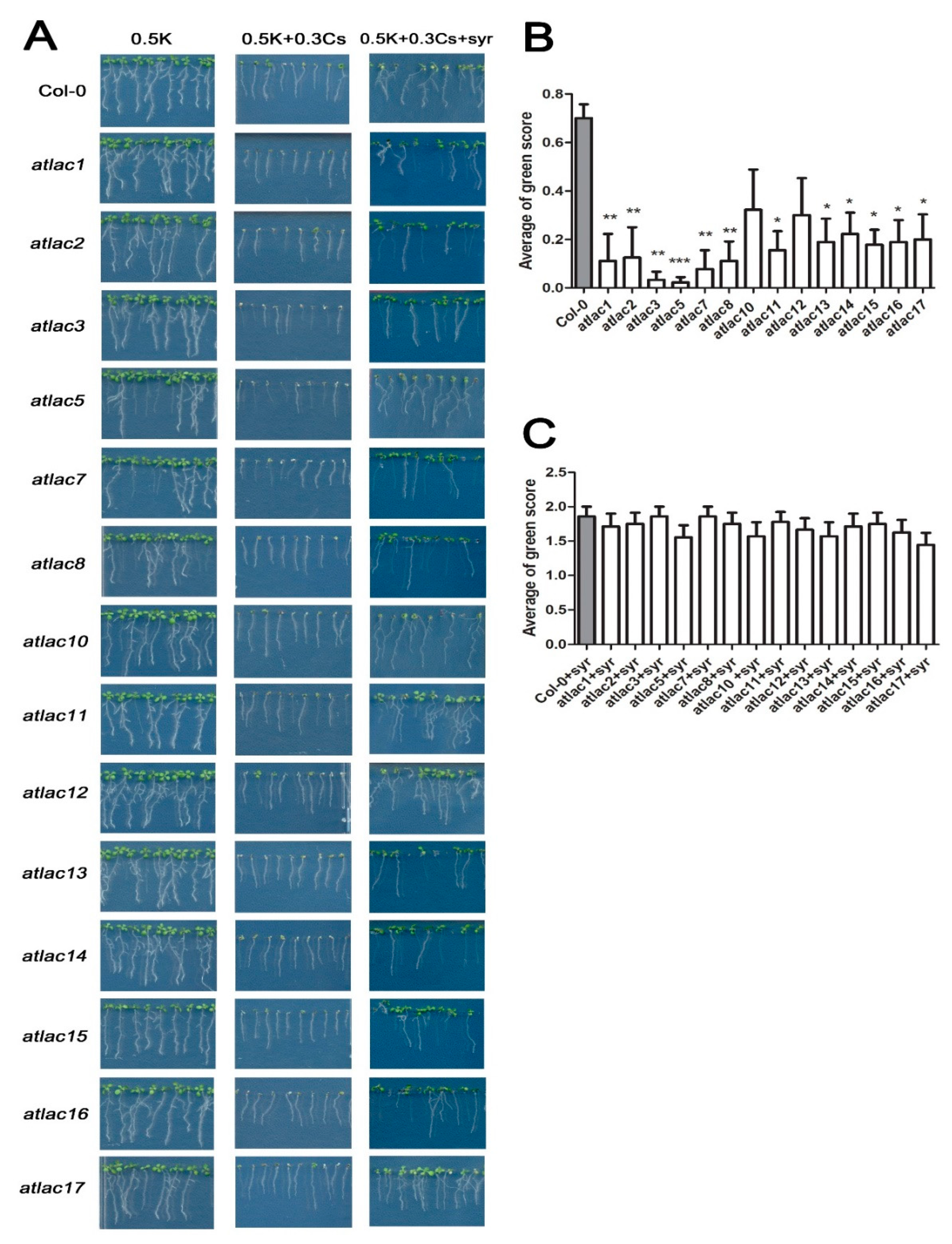
2.3. Cesium and Syringic Acid Treatments Result in Increased Lignin Deposition in Arabidopsis
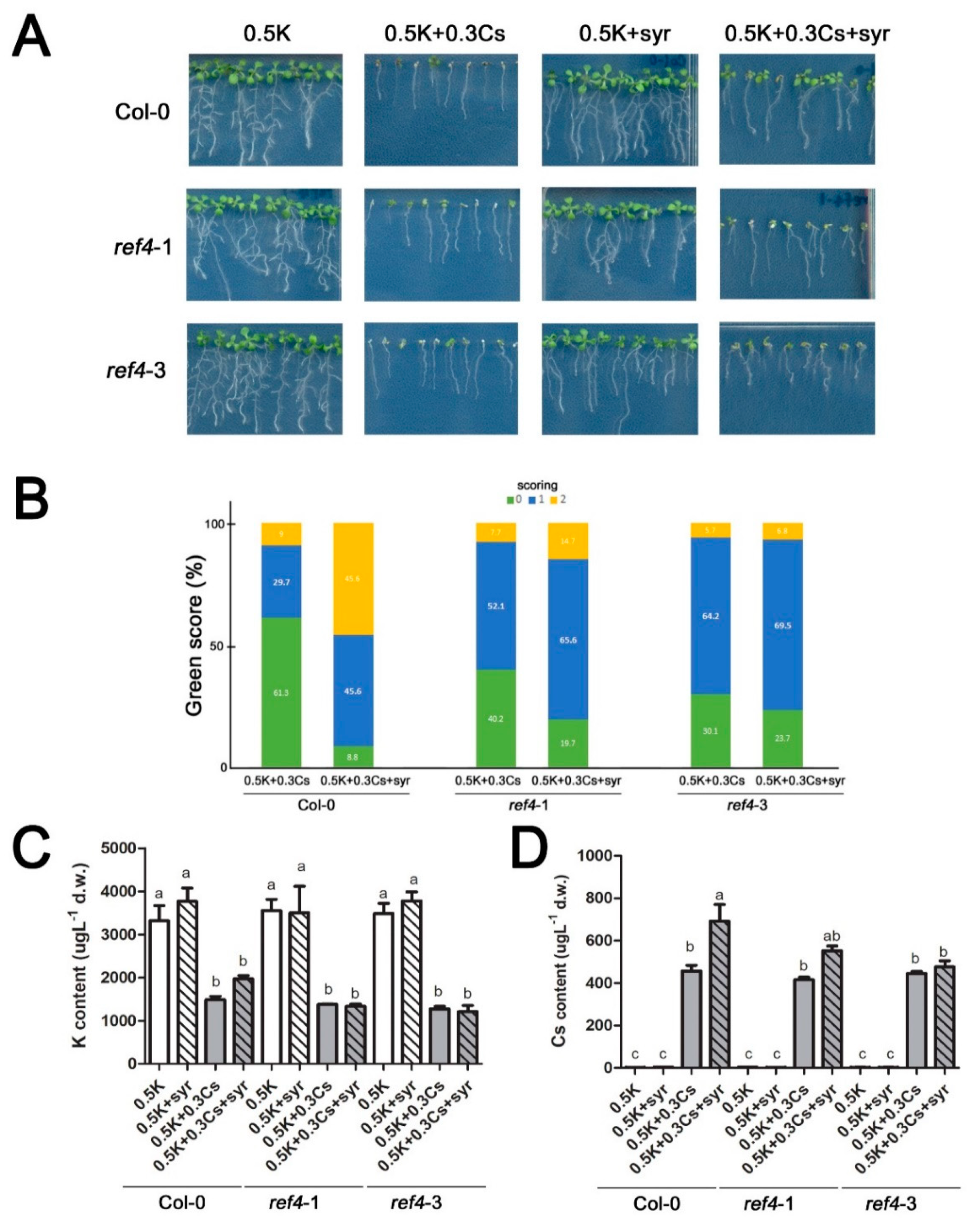
2.4. Absence of REF4 Expression Negates Syringic Acid Alleviation of Cesium-Induced Growth Retardation
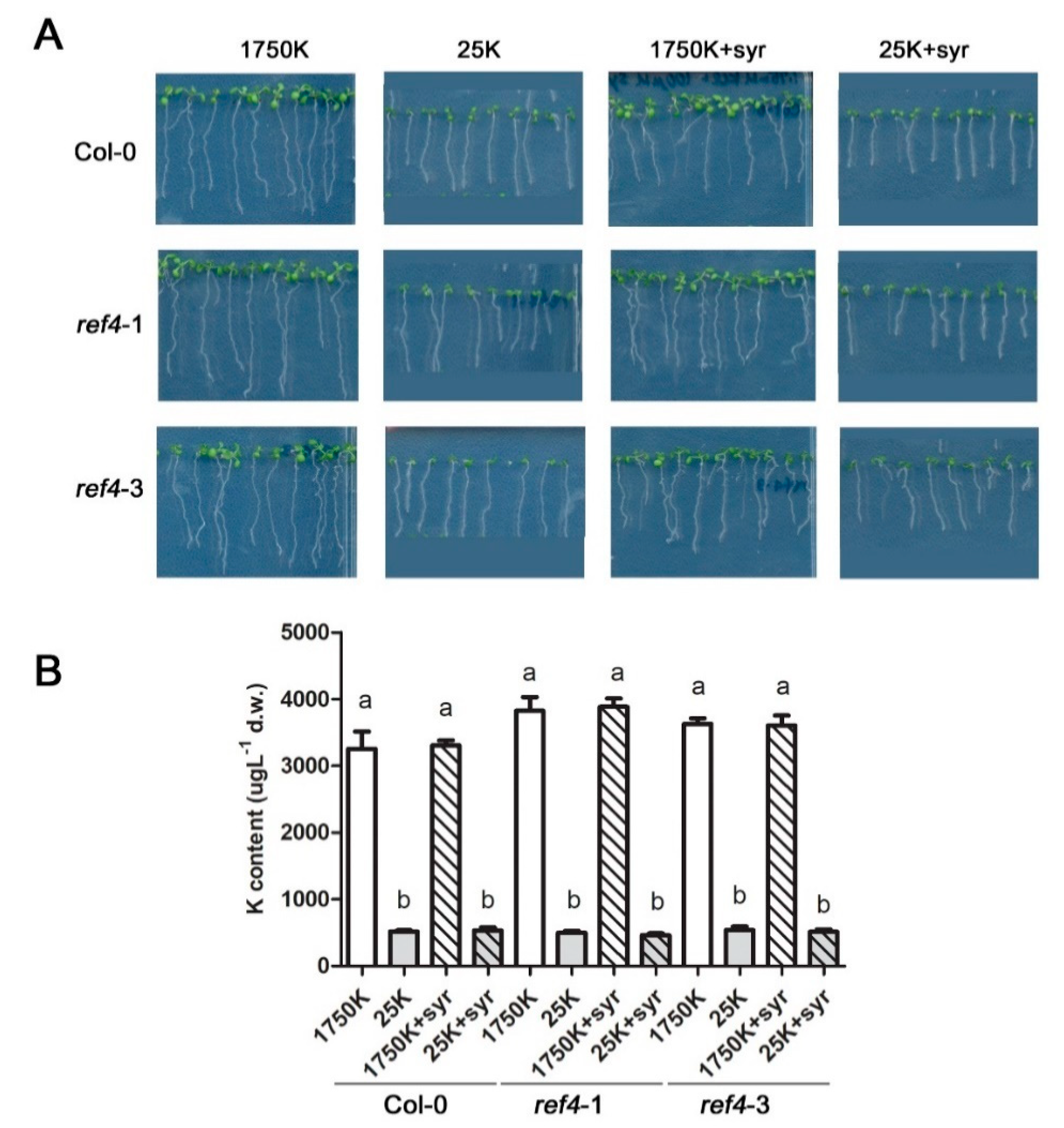
2.5. Syringic Acid Has No Effect on Low Potassium Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. Elemental Analysis
4.3. Phenotyping
4.4. Syringic Acid Quantification
4.5. Lignin Staining
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AtLAC | Arabidopsis LACCASE |
| DMBQ | 1,6-dimethoxybenzoquinone |
| NADPH oxidase | nicotinamide adenine dinucleotide phosphate oxidase |
| REF4 | REDUCED EPIDERMAL FLUORESCENE 4 |
References
- Adams, E.; Abdollahi, P.; Shin, R. Cesium inhibits plant growth through jasmonate signaling in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 4545–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.; Miyazaki, T.; Saito, S.; Uozumi, N.; Shin, R. Cesium inhibits plant growth primarily through reduction of potassium influx and accumulation in Arabidopsis. Plant Cell Physiol. 2019, 60, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.; Miyazaki, T.; Watanabe, S.; Ohkama-Ohtsu, N.; Seo, M.; Shin, R. Glutathione and Its Biosynthetic Intermediates Alleviate Cesium Stress in Arabidopsis. Front. Plant Sci. 2019, 10, 1711. [Google Scholar] [CrossRef] [PubMed]
- Burger, A.; Lichtscheidl, I. Stable and radioactive cesium: A review about distribution in the environment, uptake and translocation in plants, plant reactions and plants’ potential for bioremediation. Sci. Total Environ. 2018, 618, 1459–1485. [Google Scholar] [CrossRef]
- Butterman, W.C.; Brooks, W.E.; Robert, G.; Reese, J. Mineral. Commodity Profiles: Cesium; U.S. Geological Survey: Richmond, VA, USA, 2005.
- Djedidi, S.; Kojima, K.; Yamaya, H.; Ohkama-Ohtsu, N.; Bellingrath-Kimura, S.D.; Watanabe, I.; Yokoyama, T. Stable cesium uptake and accumulation capacities of five plant species as influenced by bacterial inoculation and cesium distribution in the soil. J. Plant Res. 2014, 127, 585–597. [Google Scholar] [CrossRef]
- Hampton, C.R.; Bowen, H.C.; Broadley, M.R.; Hammond, J.P.; Mead, A.; Payne, K.A.; Pritchard, J.; White, P.J. Cesium toxicity in Arabidopsis. Plant Physiol. 2004, 136, 3824–3837. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Liao, S.; Guo, J.; Song, Z.; Wang, R.; Zhou, X. Growth and cesium uptake responses of Phytolacca americana Linn. and Amaranthus cruentus L. grown on cesium contaminated soil to elevated CO2 or inoculation with a plant growth promoting rhizobacterium Burkholderia sp. D54, or in combination. J. Hazard. Mater. 2011, 198, 188–197. [Google Scholar] [CrossRef]
- Adams, E.; Miyazaki, T.; Hayaishi-Satoh, A.; Han, M.; Kusano, M.; Khandelia, H.; Saito, K.; Shin, R. A novel role for methyl cysteinate, a cysteine derivative, in cesium accumulation in Arabidopsis thaliana. Sci. Rep. 2017, 7, 43170. [Google Scholar] [CrossRef] [Green Version]
- Genger, R.K.; Jurkowski, G.I.; McDowell, J.M.; Lu, H.; Jung, H.W.; Greenberg, J.T.; Bent, A.F. Signaling pathways that regulate the enhanced disease resistance of Arabidopsis “defense, no death” mutants. Mol. Plant-Microbe Interact. 2008, 21, 1285–1296. [Google Scholar] [CrossRef] [Green Version]
- Horvat, D.; Simic, G.; Drezner, G.; Lalic, A.; Ledencan, T.; Tucak, M.; Plavsic, H.; Andric, L.; Zdunic, Z. Phenolic Acid Profiles and Antioxidant Activity of Major Cereal Crops. Antioxidants 2020, 9, 527. [Google Scholar] [CrossRef]
- Karamaæ, M.; Kosinska, A.; Pegg, R.B. Comparison of radical–scavenging activities of selected phenolic acids. Pol. J. Food Nutr. Sci. 2005, 14, 165–170. [Google Scholar]
- Cheemanapalli, S.; Mopuri, R.; Golla, R.; Anuradha, C.M.; Chitta, S.K. Syringic acid (SA) A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Cotoras, M.; Vivanco, H.; Melo, R.; Aguirre, M.; Silva, E.; Mendoza, L. In vitro and in vivo evaluation of the antioxidant and prooxidant activity of phenolic compounds obtained from grape (Vitis vinifera) pomace. Molecules 2014, 19, 21154–21167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancak, E.B.; Akbas, A.; Silan, C.; Cakir, D.U.; Turkon, H.; Ozkanli, S.S. Protective effect of syringic acid on kidney ischemia-reperfusion injury. Renal Fail. 2016, 38, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, S.; Mateen, S.; Naeem, S.S.; Akhtar, K.; Rizvi, W.; Moin, S. Syringic acid protects from isoproterenol induced cardiotoxicity in rats. Eur. J. Pharmacol. 2019, 849, 135–145. [Google Scholar] [CrossRef]
- Tanaka, T.; Onuma, H.; Shigihara, T.; Kimura, E.; Fukuta, Y.; Shirasaka, N.; Moriyama, T.; Homma, Y. Anti-osteoporotic effects of syringic acid and vanilic acid in the extracts of waste beds after mushroom cultivation. J. Biosci. Bioeng. 2019, 128, 622–629. [Google Scholar] [CrossRef]
- Tokmak, M.; Sehitoglu, M.H.; Yuksel, Y.; Guven, M.; Akman, T.; Aras, A.B.; Yaka, U.; Gomleksiz, C.; Albayrak, S.B.; Cosar, M. The Axon Protective Effects of Syringic Acid on Ischemia/Reperfusion Injury in a Rat Sciatic Nerve Model. Turk. Neurosurg. 2017, 27, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Velu, P.; Vijayalakshmi, A.; Vinothkumar, V. Syringic acid suppresses oral squamous cell carcinoma SCC131 cell proliferation via modulation of mitochondria-mediated apoptosis signaling pathways. J. Biochem. Mol. Toxicol. 2020, e22586. [Google Scholar] [CrossRef]
- Wei, X.; Chen, D.; Yi, Y.; Qi, H.; Gao, X.; Fang, H.; Gu, Q.; Wang, L.; Gu, L. Syringic Acid Extracted from Herba dendrobii Prevents Diabetic Cataract Pathogenesis by Inhibiting Aldose Reductase Activity. Evid.-Based Complement. Altern. Med. 2012, 2012, 426537. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Dang, H.; Liu, L.; Hu, X.; Li, X.; Ma, Z.; Wang, X.; Ren, T. Effect of syringic acid incorporation on the physical, mechanical, structural and antibacterial properties of chitosan film for quail eggs preservation. Int. J. Biol. Macromol. 2019, 141, 876–884. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, T.; Masai, E.; Miyauchi, K.; Katayama, Y.; Fukuda, M. A tetrahydrofolate-dependent O-demethylase, LigM, is crucial for catabolism of vanillate and syringate in Sphingomonas paucimobilis SYK-6. J. Bacteriol. 2005, 187, 2030–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, T.; Umeda, S.; Kamimura, N.; Kasai, D.; Kumano, S.; Abe, T.; Kawazu, C.; Otsuka, Y.; Nakamura, M.; Katayama, Y.; et al. Regulation of vanillate and syringate catabolism by a MarR-type transcriptional regulator DesR in Sphingobium sp. SYK-6. Sci. Rep. 2019, 9, 18036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.G.; Muchero, W. Regulation of Lignin Biosynthesis and Its Role in Growth-Defense Tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthet, S.; Thevenin, J.; Baratiny, D.; Domont-Caulet, N.; Debeaujon, I.; Bidznski, P.; Leple, J.-C.; Huis, R.; Hawkins, S.; Gomez, L.-D.; et al. Chapter 5—Role of plant laccases in lignin polymerization. Adv. Bot. Res. 2012, 61, 145–172. [Google Scholar] [CrossRef]
- Campos, M.L.; Yoshida, Y.; Major, I.T.; de Oliveira Ferreira, D.; Weraduwage, S.M.; Froehlich, J.E.; Johnson, B.F.; Kramer, D.M.; Jander, G.; Sharkey, T.D.; et al. Rewiring of jasmonate and phytochrome B signalling uncouples plant growth-defense tradeoffs. Nat. Commun. 2016, 7, 12570. [Google Scholar] [CrossRef] [Green Version]
- Bonawitz, N.D.; Soltau, W.L.; Blatchley, M.R.; Powers, B.L.; Hurlock, A.K.; Seals, L.A.; Weng, J.K.; Stout, J.; Chapple, C. REF4 and RFR1, subunits of the transcriptional coregulatory complex mediator, are required for phenylpropanoid homeostasis in Arabidopsis. J. Biol. Chem. 2012, 287, 5434–5445. [Google Scholar] [CrossRef] [Green Version]
- Bonawitz, N.D.; Kim, J.I.; Tobimatsu, Y.; Ciesielski, P.N.; Anderson, N.A.; Ximenes, E.; Maeda, J.; Ralph, J.; Donohoe, B.S.; Ladisch, M.; et al. Disruption of Mediator rescues the stunted growth of a lignin-deficient Arabidopsis mutant. Nature 2014, 509, 376–380. [Google Scholar] [CrossRef]
- Ruegger, M.; Chapple, C. Mutations that reduce sinapoylmalate accumulation in Arabidopsis thaliana define loci with diverse roles in phenylpropanoid metabolism. Genetics 2001, 159, 1741–1749. [Google Scholar]
- Stout, J.; Romero-Severson, E.; Ruegger, M.O.; Chapple, C. Semidominant mutations in reduced epidermal fluorescence 4 reduce phenylpropanoid content in Arabidopsis. Genetics 2008, 178, 2237–2251. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Q.; Li, C. The plant Mediator complex and its role in jasmonate signaling. J. Exp. Bot. 2019, 70, 3415–3424. [Google Scholar] [CrossRef]
- Lai, Z.; Schluttenhofer, C.M.; Bhide, K.; Shreve, J.; Thimmapuram, J.; Lee, S.Y.; Yun, D.J.; Mengiste, T. MED18 interaction with distinct transcription factors regulates multiple plant functions. Nat. Commun. 2014, 5, 3064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef]
- Shin, R.; Schachtman, D.P. Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proc. Natl. Acad. Sci. USA 2004, 101, 8827–8832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkheiri, N.; Bouguerne, B.; Bedos-Belval, F.; Duran, H.; Bemis, C.; Salvayre, R.; Negre-Salvayre, A.; Baltas, M. Synthesis and antioxidant activity evaluation of a syringic hydrazones family. Eur. J. Med. Chem. 2010, 45, 3019–3026. [Google Scholar] [CrossRef] [PubMed]
- Cikman, O.; Soylemez, O.; Ozkan, O.F.; Kiraz, H.A.; Sayar, I.; Ademoglu, S.; Taysi, S.; Karaayvaz, M. Antioxidant Activity of Syringic Acid Prevents Oxidative Stress in l-arginine-Induced Acute Pancreatitis: An Experimental Study on Rats. Int. Surg. 2015, 100, 891–896. [Google Scholar] [CrossRef]
- Keller, T.; Damude, H.G.; Werner, D.; Doerner, P.; Dixon, R.A.; Lamb, C. A plant homolog of the neutrophil NADPH oxidase gp91phox subunit gene encodes a plasma membrane protein with Ca2+ binding motifs. Plant Cell 1998, 10, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A.; Dangl, J.L.; Jones, J.D. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Unmezawa, T. Lignin modification in planta for valorization. Phytochem. Rev. 2018, 17, 1305–1327. [Google Scholar] [CrossRef]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cezard, L.; Le Bris, P.; Borrega, N.; Herve, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Nakashima, J.; Chen, F.; Yin, Y.; Fu, C.; Yun, J.; Shao, H.; Wang, X.; Wang, Z.Y.; Dixon, R.A. Laccase is necessary and nonredundant with peroxidase for lignin polymerization during vascular development in Arabidopsis. Plant Cell 2013, 25, 3976–3987. [Google Scholar] [CrossRef] [Green Version]
- Brogioni, B.; Biglino, D.; Sinicropi, A.; Reijerse, E.J.; Giardina, P.; Sannia, G.; Lubitz, W.; Basosi, R.; Pogni, R. Characterization of radical intermediates in laccase-mediator systems. A multifrequency EPR, ENDOR and DFT/PCM investigation. Phys. Chem. Chem. Phys. 2008, 10, 7284–7292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Iaropolov, A.I. Laccase-mediator systems and their applications: A review. Prikl. Biokhimiia Mikrobiol. 2007, 43, 583–597. [Google Scholar] [CrossRef]
- Cai, X.; Davis, E.J.; Ballif, J.; Liang, M.; Bushman, E.; Haroldsen, V.; Torabinejad, J.; Wu, Y. Mutant identification and characterization of the laccase gene family in Arabidopsis. J. Exp. Bot. 2006, 57, 2563–2569. [Google Scholar] [CrossRef] [Green Version]
- Volkova, N.; Ibrahim, V.; Hatti-Kaul, R. Laccase catalysed oxidation of syringic acid: Calorimetric determination of kinetic parameters. Enzym. Microb. Technol. 2012, 50, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.; Miyazaki, T.; Shin, R. Contribution of KUPs to potassium and cesium accumulation appears complementary in Arabidopsis. Plant Signal. Behav. 2019, 14, 1554468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, I.L.; Ryu, M.; Cho, S.K.; Shah, P.; Lee, J.H.; Bae, H.; Kim, I.G.; Yang, S.W. Cesium toxicity alters microRNA processing and AGO1 expressions in Arabidopsis thaliana. PLoS ONE 2015, 10, e0125514. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, D.; Uozumi, N.; Hisamatsu, S.; Yamagami, M. AtKUP/HAK/KT9, a K+ transporter from Arabidopsis thaliana, mediates Cs+ uptake in Escherichia coli. Biosci. Biotechnol. Biochem. 2010, 74, 203–205. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.; Hampton, C.R.; Shin, R.; Barkla, B.J.; White, P.J.; Schachtman, D.P. The high affinity K+ transporter AtHAK5 plays a physiological role in planta at very low K+ concentrations and provides a caesium uptake pathway in Arabidopsis. J. Exp. Bot. 2008, 59, 595–607. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.Y.; Shin, R.; Schachtman, D.P. Ethylene mediates response and tolerance to potassium deprivation in Arabidopsis. Plant Cell 2009, 21, 607–621. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Ruzicka, D.; Shin, R.; Schachtman, D.P. The Arabidopsis AP2/ERF transcription factor RAP2.11 modulates plant response to low-potassium conditions. Mol. Plant 2012, 5, 1042–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.J.; Tran, L.S.; Kojima, M.; Sakakibara, H.; Nishiyama, R.; Shin, R. Regulatory roles of cytokinins and cytokinin signaling in response to potassium deficiency in Arabidopsis. PLoS ONE 2012, 7, e47797. [Google Scholar] [CrossRef] [Green Version]
- Troufflard, S.; Mullen, W.; Larson, T.R.; Graham, I.A.; Crozier, A.; Amtmann, A.; Armengaud, P. Potassium deficiency induces the biosynthesis of oxylipins and glucosinolates in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behr, M.; Pokorna, E.; Dobrev, P.I.; Motyka, V.; Guignard, C.; Lutts, S.; Hausman, J.F.; Guerriero, G. Impact of jasmonic acid on lignification in the hemp hypocotyl. Plant Signal. Behav. 2019, 14, 1592641. [Google Scholar] [CrossRef] [Green Version]
- Denness, L.; McKenna, J.F.; Segonzac, C.; Wormit, A.; Madhou, P.; Bennett, M.; Mansfield, J.; Zipfel, C.; Hamann, T. Cell wall damage-induced lignin biosynthesis is regulated by a reactive oxygen species- and jasmonic acid-dependent process in Arabidopsis. Plant Physiol. 2011, 156, 1364–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Zhang, L.; Sun, S.; Yi, Z.; Jiang, X.; Jia, D. Neuroprotective effects of syringic acid against OGD/R-induced injury in cultured hippocampal neuronal cells. Int. J. Mol. Med. 2016, 38, 567–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gheena, S.; Ezhilarasan, D. Syringic acid triggers reactive oxygen species-mediated cytotoxicity in HepG2 cells. Hum. Exp. Toxicol. 2019, 38, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.R.; Lee, H.I.; Choi, R.Y.; Sim, M.O.; Seo, K.I.; Lee, M.K. Anti-steatotic and anti-inflammatory roles of syringic acid in high-fat diet-induced obese mice. Food Funct. 2016, 7, 689–697. [Google Scholar] [CrossRef]
- Itoh, A.; Isoda, K.; Kondoh, M.; Kawase, M.; Kobayashi, M.; Tamesada, M.; Yagi, K. Hepatoprotective effect of syringic acid and vanillic acid on concanavalin a-induced liver injury. Biol. Pharm. Bull. 2009, 32, 1215–1219. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Yan, B.C.; Park, J.H.; Yeun, G.H.; Yim, Y.; Ahn, J.H.; Lee, J.C.; Hwang, I.K.; Cho, J.H.; Kim, Y.M.; et al. Neuroprotection of a novel synthetic caffeic acid-syringic acid hybrid compound against experimentally induced transient cerebral ischemic damage. Planta Med. 2013, 79, 313–321. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, J.; Lei, T.; Wang, W.; Lin, A. Anti-endotoxic effects of syringic acid of Radix Isatidis. J. Huazhong Univ. Sci. Technol. Med Sci. 2003, 23, 206–208. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M. Seasonal fluctuations in soil phenolics of a coniferous forest: Effects on seed germination of different coniferous species. Plant Soil 2006, 284, 305–318. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Wu, F.; Zhou, X. Changes in rhizosphere microbial communities in potted cucumber seedlings treated with syringic acid. PLoS ONE 2018, 13, e0200007. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Kaur, R.; Kaur, S.; Baldwin, I.T.; Inderjit. Taking ecological function seriously: Soil microbial communities can obviate allelopathic effects of released metabolites. PLoS ONE 2009, 4, e4700. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Autotoxicity: Concept, Organisms, and Ecological Significance. Critic. Rev. Plant Sci. 2010, 18, 757–772. [Google Scholar] [CrossRef]
- Yu, J.Q.; Matsui, Y. Phytotoxic substances in root exudates of cucumber (Cucumis sativus L.). J. Chem. Ecol. 1994, 20, 21–31. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K.; Jana, M.M.; Gupta, A. Improvement of selective lignin degradation in fungal pretreatment of sweet sorghum bagasse using synergistic CuSO4-syringic acid supplements. J. Environ. Manag. 2017, 193, 558–566. [Google Scholar] [CrossRef]
- Adams, E.; Chaban, V.; Khandelia, H.; Shin, R. Selective chemical binding enhances cesium tolerance in plants through inhibition of cesium uptake. Sci. Rep. 2015, 5, 8842. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q. Lignification: Flexibility, Biosynthesis and Regulation. Trends Plant Sci. 2016, 21, 713–721. [Google Scholar] [CrossRef]
- Laohavisit, A.; Wakatake, T.; Ishihama, N.; Mulvey, H.; Takizawa, K.; Suzuki, T.; Shirasu, K. Quinone perception in plants via leucine-rich-repeat receptor-like kinases. Nature 2020, 587, 92–97. [Google Scholar] [CrossRef]
- Moon, J.Y.; Belloeil, C.; Ianna, M.L.; Shin, R. Arabidopsis CNGC Family Members Contribute to Heavy Metal Ion Uptake in Plants. Int. J. Mol. Sci. 2019, 20, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; Bednarek, P.; Liu, J.; Schneider, B.; Svatos, A.; Hahlbrock, K. Universally occurring phenylpropanoid and species-specific indolic metabolites in infected and uninfected Arabidopsis thaliana roots and leaves. Phytochemistry 2004, 65, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, Y.; Akiyama, K.; Sakata, A.; Kuwahara, A.; Otsuki, H.; Sakurai, T.; Saito, K.; Hirai, M.Y. Widely targeted metabolomics based on large-scale MS/MS data for elucidating metabolite accumulation patterns in plants. Plant Cell Physiol. 2009, 50, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Sun, M.; Wang, F.; Liu, J.; Feng, B.; Si, J.; Zhang, B.; Li, S.; Li, H. Effects of high NH4+ on K+ uptake, culm mechanical strength and grain filling in wheat. Front. Plant Sci. 2014, 5, 703. [Google Scholar] [CrossRef]
- Pradhan Mitra, P.; Loque, D. Histochemical staining of Arabidopsis thaliana secondary cell wall elements. JoVE 2014, 87, e51381. [Google Scholar] [CrossRef] [Green Version]
- Speer, E.O. A method of retaining phloroglucinol proof of lignin. Stain Technol. 1987, 62, 279–280. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adams, E.; Miyazaki, T.; Moon, J.Y.; Sawada, Y.; Sato, M.; Toyooka, K.; Hirai, M.Y.; Shin, R. Syringic Acid Alleviates Cesium-Induced Growth Defect in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 9116. https://doi.org/10.3390/ijms21239116
Adams E, Miyazaki T, Moon JY, Sawada Y, Sato M, Toyooka K, Hirai MY, Shin R. Syringic Acid Alleviates Cesium-Induced Growth Defect in Arabidopsis. International Journal of Molecular Sciences. 2020; 21(23):9116. https://doi.org/10.3390/ijms21239116
Chicago/Turabian StyleAdams, Eri, Takae Miyazaki, Ju Yeon Moon, Yuji Sawada, Muneo Sato, Kiminori Toyooka, Masami Yokota Hirai, and Ryoung Shin. 2020. "Syringic Acid Alleviates Cesium-Induced Growth Defect in Arabidopsis" International Journal of Molecular Sciences 21, no. 23: 9116. https://doi.org/10.3390/ijms21239116
APA StyleAdams, E., Miyazaki, T., Moon, J. Y., Sawada, Y., Sato, M., Toyooka, K., Hirai, M. Y., & Shin, R. (2020). Syringic Acid Alleviates Cesium-Induced Growth Defect in Arabidopsis. International Journal of Molecular Sciences, 21(23), 9116. https://doi.org/10.3390/ijms21239116