Neuroprotective Effect of Cudrania tricuspidata Fruit Extracts on Scopolamine-Induced Learning and Memory Impairment

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cytotoxicity of SCO and CTFE on PC12 Cells
2.2. AChE Inhibitory Effect of CTFE on PC12 Cells
2.3. Effect of CTFE on Protein Levels of CREB and ERK1/2 in PC12 Cells
2.4. Effect of CTFE on SCO-Induced Learning and Memory Dysfunction of Mice in BMT
2.5. Effect of CTFE on SCO-Induced Learning and Memory Dysfunction of Mice in the FCT
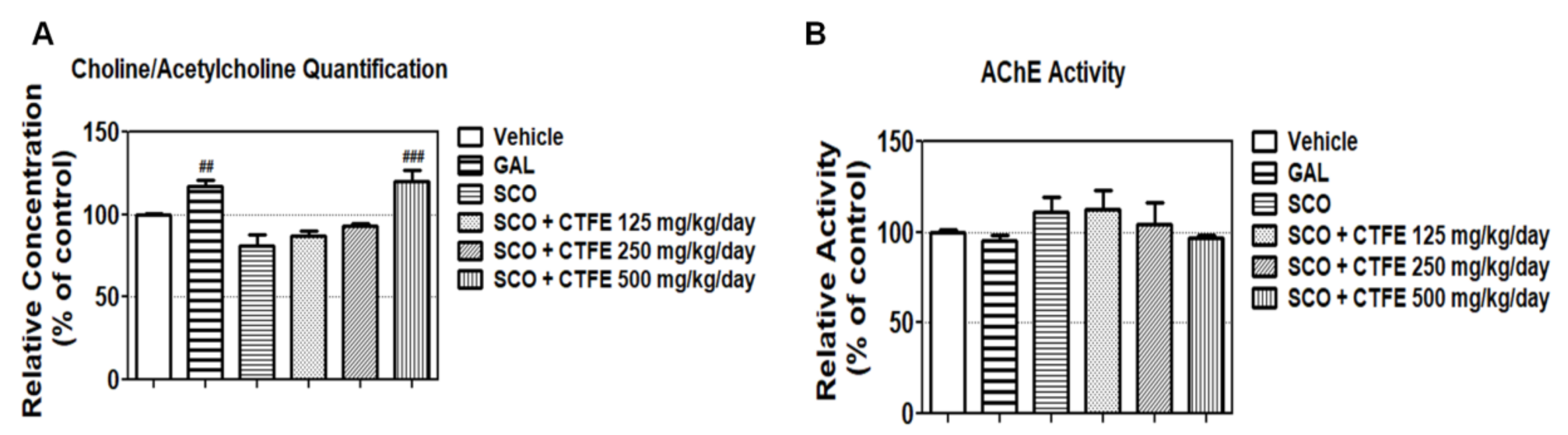
2.6. Effect of CTFE on ACh Concentration and AChE Activity in Mice
2.7. Effect of CTFE on Protein Levels of SIRT3 and AMPK in Hippocampal Tissue of Mice
2.8. Global Gene Expression in Hippocampal Tissue of Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Extract
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. AChE Activity Assay
4.6. Western Blot Analysis of PC12 Cells
4.7. Animals and Drug Treatments
- Group 1:
- water, p.o. and normal saline, i.p. (vehicle control)
- Group 2:
- water, p.o. + 2 mg/kg GAL, i.p. (positive control)
- Group 3:
- water, p.o. + 1 mg/kg SCO, i.p. (negative control)
- Group 4:
- 125 mg/kg/day CTFE, p.o. + 1 mg/kg SCO, i.p.
- Group 5:
- 250 mg/kg/day CTFE, p.o. + 1 mg/kg SCO, i.p.
- Group 6:
- 500 mg/kg/day CTFE, p.o. + 1 mg/kg SCO, i.p.
4.8. Barnes Maze Test (BMT)
4.9. Fear Conditioning Test (FCT)
4.10. Blood Sampling and ACh Quantification
4.11. Hippocampal Tissue Homogenization
4.12. AChE Activity in Hippocampal Tissue
4.13. Microarray Gene Expression Analysis
4.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Pen, A.E.; Jensen, U.B. Current status of treating neurodegenerative disease with induced pluripotent stem cells. Acta Neurol. Scand. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallett, P.J.; Engelender, S.; Isacson, O. Lipid and immune abnormalities causing age-dependent neurodegeneration and Parkinson’s disease. J. Neuroinflamm. 2019, 16, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domaradzki, J. The Impact of Huntington Disease on Family Carers: A Literature Overview. Psychiatr. Pol. 2015, 49, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Lauterbach, E.C.; Victoroff, J.; Coburn, K.L.; Shillcutt, S.D.; Doonan, S.M.; Mendez, M.F. Psychopharmacological neuroprotection in neurodegenerative disease: Assessing the preclinical data. J. Neuropsychiatry Clin. Neurosci. 2010, 22, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Mangialasche, F.; Solomon, A.; Winblad, B.; Mecocci, P.; Kivipelto, M. Alzheimer’s disease: Clinical trials and drug development. Lancet Neurol. 2010, 9, 702–716. [Google Scholar] [CrossRef]
- Polinsky, R.J. Clinical pharmacology of rivastigmine: A new-generation acetylcholinesterase inhibitor for the treatment of Alzheimer’s disease. Clin. Ther. 1998, 20, 634–647. [Google Scholar] [CrossRef]
- Rogers, S.L.; Farlow, M.R.; Doody, R.S.; Mohs, R.; Friedhoff, L.T. A 24-week, double-blind, placebo-controlled trial of donepezil in patients with Alzheimer’s disease. Donepezil Study Group. Neurology 1998, 50, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Weon, J.B.; Yun, B.R.; Lee, J.; Eom, M.R.; Ko, H.J.; Lee, H.Y.; Park, D.S.; Chung, H.C.; Chung, J.Y.; Ma, C.J. Cognitive-Enhancing Effect of Steamed and Fermented Codonopsis lanceolata: A Behavioral and Biochemical Study. Evid. Based Complement. Alternat. Med. 2014, 2014, 319436. [Google Scholar] [CrossRef] [Green Version]
- Kirsch, S.H.; Herrmann, W.; Rabagny, Y.; Obeid, R. Quantification of acetylcholine, choline, betaine, and dimethylglycine in human plasma and urine using stable-isotope dilution ultra performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2010, 878, 3338–3344. [Google Scholar] [CrossRef]
- Shin, Y.; Cho, N.J. The C. elegans VIG-1 and FRM-1 modulate carbachol-stimulated ERK1/2 activation in chinese hamster ovary cells expressing the muscarinic acetylcholine receptor GAR-3. Neurochem. Res. 2014, 39, 777–784. [Google Scholar] [CrossRef]
- Klinkenberg, I.; Blokland, A. The validity of scopolamine as a pharmacological model for cognitive impairment: A review of animal behavioral studies. Neurosci. Biobehav. Rev. 2010, 34, 1307–1350. [Google Scholar] [CrossRef] [PubMed]
- Frumin, M.J.; Herekar, V.R.; Jarvik, M.E. Amnesic actions of diazepam and scopolamine in man. Anesthesiology 1976, 45, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.S.; Rai, S.; Katyal, A. Cholinergic receptor blockade by scopolamine and mecamylamine exacerbates global cerebral ischemia induced memory dysfunction in C57BL/6J mice. Nitric Oxide 2014, 43, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.-T.; Yue, S.-J.; Fan, Y.-C.; Wu, J.-S.; Yan, D.; Guan, H.-S.; Wang, C.-Y. Cudraniatricuspidata: An updated review on ethnomedicine, phytochemistry and pharmacology. RSC Adv. 2017, 7, 31807–31832. [Google Scholar] [CrossRef] [Green Version]
- Song, S.H.; Ki, S.H.; Park, D.H.; Moon, H.S.; Lee, C.D.; Yoon, I.S.; Cho, S.S. Quantitative Analysis, Extraction Optimization, and Biological Evaluation of Cudraniatricuspidata Leaf and Fruit Extracts. Molecules 2017, 22. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.; Kwon, J.; Lee, D.; Mar, W. Effects of Cudraniatricuspidata Fruit Extract and Its Active Compound, 5,7,3′,4′-Tetrahydroxy-6,8-diprenylisoflavone, on the High-Affinity IgE Receptor-Mediated Activation of Syk in Mast Cells. J. Agric. Food Chem. 2015, 63, 5459–5467. [Google Scholar] [CrossRef]
- Li, X.; Yao, Z.; Jiang, X.; Sun, J.; Ran, G.; Yang, X.; Zhao, Y.; Yan, Y.; Chen, Z.; Tian, L.; et al. Bioactive compounds from Cudraniatricuspidata: A natural anticancer source. Crit. Rev. Food Sci. Nutr. 2020, 60, 494–514. [Google Scholar] [CrossRef]
- Hiep, N.T.; Kwon, J.; Kim, D.W.; Hwang, B.Y.; Lee, H.J.; Mar, W.; Lee, D. Isoflavones with neuroprotective activities from fruits of Cudraniatricuspidata. Phytochemistry 2015, 111, 141–148. [Google Scholar] [CrossRef]
- Ebrahimzadeh, A.; Moghadam, S.Y.; Rahimi, H.; Motaghinejad, M.; Motevalian, M.; Safari, S.; Mesrabadi, M.A. Crocin acts as a neuroprotective mediator against methylphenidateinduced neurobehavioral and neurochemical sequelae: Possible role of the CREB-BDNF signaling pathway. Acta Neurobiol. Exp. 2019, 79, 352–366. [Google Scholar]
- Sakamoto, K.; Karelina, K.; Obrietan, K. CREB: A multifaceted regulator of neuronal plasticity and protection. J. Neurochem. 2011, 116, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Saura, C.A.; Valero, J. The role of CREB signaling in Alzheimer’s disease and other cognitive disorders. Rev. Neurosci. 2011, 22, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Pugazhenthi, S.; Wang, M.; Pham, S.; Sze, C.I.; Eckman, C.B. Downregulation of CREB expression in Alzheimer’s brain and in Abeta-treated rat hippocampal neurons. Mol. Neurodegener. 2011, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Medina, J.H.; Viola, H. ERK1/2: A Key Cellular Component for the Formation, Retrieval, Reconsolidation and Persistence of Memory. Front. Mol. Neurosci. 2018, 11, 361. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nan, G. The extracellular signal-regulated kinase 1/2 pathway in neurological diseases: A potential therapeutic target (Review). Int. J. Mol. Med. 2017, 39, 1338–1346. [Google Scholar] [CrossRef] [Green Version]
- Teich, A.F.; Nicholls, R.E.; Puzzo, D.; Fiorito, J.; Purgatorio, R.; Fa, M.; Arancio, O. Synaptic therapy in Alzheimer’s disease: A CREB-centric approach. Neurotherapeutics 2015, 12, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, C.A.; Oliveira, W.H.; Araujo, S.; Nunes, A.K.S. AMPK activation: Role in the signaling pathways of neuroinflammation and neurodegeneration. Exp. Neurol. 2017, 298, 31–41. [Google Scholar] [CrossRef]
- Marinangeli, C.; Didier, S.; Vingtdeux, V. AMPK in Neurodegenerative Diseases: Implications and Therapeutic Perspectives. Curr. Drug Targets 2016, 17, 890–907. [Google Scholar] [CrossRef]
- Liu, S.-G.; Wang, Y.-M.; Zhang, Y.-J.; He, X.-J.; Ma, T.; Song, W.; Zhang, Y.-M. ZL006 protects spinal cord neurons against ischemia-induced oxidative stress through AMPK-PGC-1α-Sirt3 pathway. Neurochem. Int. 2017, 108, 230–237. [Google Scholar] [CrossRef]
- Weir, H.J.; Murray, T.K.; Kehoe, P.G.; Love, S.; Verdin, E.M.; O’Neill, M.J.; Lane, J.D.; Balthasar, N. CNS SIRT3 expression is altered by reactive oxygen species and in Alzheimer’s disease. PLoS ONE 2012, 7, e48225. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.H. Mechanisms and disease implications of sirtuin-mediated autophagic regulation. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Nuttall, J.R.; Oteiza, P.I. Zinc and the ERK kinases in the developing brain. Neurotoxic. Res. 2012, 21, 128–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Jin, J.; Cichewicz, R.H.; Hageman, S.A.; Ellis, T.K.; Xiang, L.; Peng, Q.; Jiang, M.; Arbez, N.; Hotaling, K.; et al. Trans-(-)-epsilon-Viniferin increases mitochondrial sirtuin 3 (SIRT3), activates AMP-activated protein kinase (AMPK), and protects cells in models of Huntington Disease. J. Biol. Chem. 2012, 287, 24460–24472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, D.B.; Zwaans, B.M.M. SIRT3: As simple as it seems? Gerontology 2014, 60, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Pandareesh, M.D.; Anand, T. Neuromodulatory propensity of Bacopa monniera against scopolamine-induced cytotoxicity in PC12 cells via down-regulation of AChE and up-regulation of BDNF and muscarnic-1 receptor expression. Cell Mol. Neurobiol. 2013, 33, 875–884. [Google Scholar] [CrossRef]
- Maurer, S.V.; Williams, C.L. The Cholinergic System Modulates Memory and Hippocampal Plasticity via Its Interactions with Non-Neuronal Cells. Front. Immunol. 2017, 8, 1489. [Google Scholar] [CrossRef] [Green Version]
- Gold, P.E. Acetylcholine modulation of neural systems involved in learning and memory. Neurobiol. Learn. Mem. 2003, 80, 194–210. [Google Scholar] [CrossRef]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a Biomarker in Environmental and Occupational Medicine: New Insights and Future Perspectives. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef]
- Valbonesi, P.; Franzellitti, S.; Bersani, F.; Contin, A.; Fabbri, E. Activity and expression of acetylcholinesterase in PC12 cells exposed to intermittent 1.8 GHz 217-GSM mobile phone signal. Int. J. Radiat. Biol. 2016, 92, 1–10. [Google Scholar] [CrossRef]
- Das, K.P.; Barone, S. Neuronal Differentiation in PC12 Cells Is Inhibited by Chlorpyrifos and Its Metabolites: Is Acetylcholinesterase Inhibition the Site of Action? Toxicol. Appl. Pharmacol. 1999, 160, 217–230. [Google Scholar] [CrossRef]
- Chinnadurai, R.K.; Saravanaraman, P.; Boopathy, R. The significance of aryl acylamidase activity of acetylcholinesterase in osteoblast differentiation and mineralization. Mol. Cell Biochem. 2018, 440, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Gutierres, J.M.; Carvalho, F.B.; Schetinger, M.R.; Agostinho, P.; Marisco, P.C.; Vieira, J.M.; Rosa, M.M.; Bohnert, C.; Rubin, M.A.; Morsch, V.M.; et al. Neuroprotective effect of anthocyanins on acetylcholinesterase activity and attenuation of scopolamine-induced amnesia in rats. Int. J. Dev. Neurosci. 2014, 33, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Sramek, J.J.; Frackiewicz, E.J.; Cutler, N.R. Review of the acetylcholinesterase inhibitor galanthamine. Expert Opin. Investig. Drugs 2000, 9, 2393–2402. [Google Scholar] [CrossRef] [PubMed]
- Seltzer, B. Galantamine-ER for the treatment of mild-to-moderate Alzheimer’s disease. Clin. Interv. Aging 2010, 5, 1–6. [Google Scholar]
- Wu, Z.; Zhao, L.; Chen, X.; Cheng, X.; Zhang, Y. Galantamine attenuates amyloid-beta deposition and astrocyte activation in APP/PS1 transgenic mice. Exp. Gerontol. 2015, 72, 244–250. [Google Scholar] [CrossRef]
- Attar, A.; Liu, T.Y.; Chan, W.T.C.; Hayes, J.; Nejad, M.; Lei, K.C.; Bitan, G. A Shortened Barnes Maze Protocol Reveals Memory Deficits at 4-Months of Age in the Triple-Transgenic Mouse Model of Alzheimer’s Disease. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Stevens, R. Scopolamine impairs spatial maze performance in rats. Physiol. Behav. 1981, 27, 385–386. [Google Scholar] [CrossRef]
- Curzon, P.; Rustay, N.R.; Browman, K.E. Cued and Contextual Fear Conditioning for Rodents. In Methods of Behavior Analysis in Neuroscience, 2nd ed.; Buccafusco, J.J., Ed.; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Tsai, T.H. Separation methods used in the determination of choline and acetylcholine. J. Chromatogr. B 2000, 747, 111–122. [Google Scholar] [CrossRef]
- Cohen, E.L.; Wurtman, R.J. Brain acetylcholine: Control by dietary choline. Science 1976, 191, 561–562. [Google Scholar] [CrossRef]
- Blake, M.G.; Boccia, M.M.; Krawczyk, M.C.; Delorenzi, A.; Baratti, C.M. Choline reverses scopolamine-induced memory impairment by improving memory reconsolidation. Neurobiol. Learn. Mem. 2012, 98, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Talesa, V.N. Acetylcholinesterase in Alzheimer’s disease. Mech. Ageing Dev. 2001, 122, 1961–1969. [Google Scholar] [CrossRef]
- Jeon, S.J.; Rhee, S.Y.; Seo, J.E.; Bak, H.R.; Lee, S.H.; Ryu, J.H.; Cheong, J.H.; Shin, C.Y.; Kim, G.H.; Lee, Y.S.; et al. Oroxylin A increases BDNF production by activation of MAPK-CREB pathway in rat primary cortical neuronal culture. Neurosci. Res. 2011, 69, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C.M. Transcription factors in long-term memory and synaptic plasticity. Physiol. Rev. 2009, 89, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wen, Q.; Ren, L.; Liang, W.; Xia, Y.; Zhang, X.; Zhao, D.; Sun, D.; Hu, Y.; Hao, H.; et al. Neuroprotective effect of arctigenin via upregulation of P-CREB in mouse primary neurons and human SH-SY5Y neuroblastoma cells. Int. J. Mol. Sci. 2013, 14, 18657–18669. [Google Scholar] [CrossRef] [PubMed]
- Igaz, L.M.; Winograd, M.; Cammarota, M.; Izquierdo, L.A.; Alonso, M.; Izquierdo, I.; Medina, J.H. Early activation of extracellular signal-regulated kinase signaling pathway in the hippocampus is required for short-term memory formation of a fear-motivated learning. Cell Mol. Neurobiol. 2006, 26, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Feld, M.; Dimant, B.; Delorenzi, A.; Coso, O.; Romano, A. Phosphorylation of extra-nuclear ERK/MAPK is required for long-term memory consolidation in the crab Chasmagnathus. Behav. Brain Res. 2005, 158, 251–261. [Google Scholar] [CrossRef]
- Cruz, C.D.; Cruz, F. The ERK 1 and 2 pathway in the nervous system: From basic aspects to possible clinical applications in pain and visceral dysfunction. Curr. Neuropharmacol. 2007, 5, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhang, R.; Tang, S.; Ren, Y.; Yang, W.; Liu, X.; Tang, J. Orexin-A-induced ERK1/2 activation reverses impaired spatial learning and memory in pentylenetetrazol-kindled rats via OX1R-mediated hippocampal neurogenesis. Peptides 2014, 54, 140–147. [Google Scholar] [CrossRef]
- Huang, W.; Cao, J.; Liu, X.; Meng, F.; Li, M.; Chen, B.; Zhang, J. AMPK Plays a Dual Role in Regulation of CREB/BDNF Pathway in Mouse Primary Hippocampal Cells. J. Mol. Neurosci. 2015, 56, 782–788. [Google Scholar] [CrossRef]
- Marinangeli, C.; Didier, S.; Ahmed, T.; Caillerez, R.; Domise, M.; Laloux, C.; Bégard, S.; Carrier, S.; Colin, M.; Marchetti, P.; et al. AMP-Activated Protein Kinase Is Essential for the Maintenance of Energy Levels during Synaptic Activation. iScience 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.J.; Li, Y.F.; Liu, F.L.; Deng, J.; Wu, Y.P.; Yuan, W.L.; Tsoi, B.; Chen, J.L.; Wang, Q.; Cai, S.H.; et al. A SIRT3/AMPK/autophagy network orchestrates the protective effects of trans-resveratrol in stressed peritoneal macrophages and RAW 264.7 macrophages. Free Radic. Biol. Med. 2016, 95, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Kesidou, E.; Lagoudaki, R.; Touloumi, O.; Poulatsidou, K.-N.; Simeonidou, C. Autophagy and neurodegenerative disorders. Neural Regen. Res. 2013, 8, 2275–2283. [Google Scholar] [CrossRef] [PubMed]
- Cherra 3rd, S.J.; Chu, C.T. Autophagy in neuroprotection and neurodegeneration: A question of balance. Future Neurol. 2008, 3, 309–323. [Google Scholar] [CrossRef]
- Qin, Q.F.; Li, X.J.; Li, Y.S.; Zhang, W.K.; Tian, G.H.; Shang, H.C.; Tang, H.B. AMPK-ERK/CARM1 Signaling Pathways Affect Autophagy of Hepatic Cells in Samples of Liver Cancer Patients. Front. Oncol. 2019, 9, 1247. [Google Scholar] [CrossRef] [PubMed]
- Thapalia, B.A.; Zhou, Z.; Lin, X. Sauchinone augments cardiomyocyte viability by enhancing autophagy proteins -PI3K, ERK(1/2), AMPK and Beclin-1 during early ischemia-reperfusion injury in vitro. Am. J. Transl. Res. 2016, 8, 3251–3265. [Google Scholar]
- Kim, H.J.; Kim, J.; Kang, K.S.; Lee, K.T.; Yang, H.O. Neuroprotective Effect of Chebulagic Acid via Autophagy Induction in SH-SY5Y Cells. Biomol. Ther. 2014, 22, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.P.; Lang, B.T.; Vemuganti, R.; Dempsey, R.J. Osteopontin is a mediator of the lateral migration of neuroblasts from the subventricular zone after focal cerebral ischemia. Neurochem. Int. 2009, 55, 826–832. [Google Scholar] [CrossRef]
- Yu, H.; Liu, X.; Zhong, Y. The Effect of Osteopontin on Microglia. Biomed. Res. Int. 2017, 2017, 1879437. [Google Scholar] [CrossRef]
- Begcevic, I.; Brinc, D.; Brown, M.; Martinez-Morillo, E.; Goldhardt, O.; Grimmer, T.; Magdolen, V.; Batruch, I.; Diamandis, E.P. Brain-related proteins as potential CSF biomarkers of Alzheimer’s disease: A targeted mass spectrometry approach. J. Proteom. 2018, 182, 12–20. [Google Scholar] [CrossRef]
- Vandell, A.G.; Larson, N.; Laxmikanthan, G.; Panos, M.; Blaber, S.I.; Blaber, M.; Scarisbrick, I.A. Protease-activated receptor dependent and independent signaling by kallikreins 1 and 6 in CNS neuron and astroglial cell lines. J. Neurochem. 2008, 107, 855–870. [Google Scholar] [CrossRef] [Green Version]
- Mella, C.; Figueroa, C.D.; Otth, C.; Ehrenfeld, P. Involvement of Kallikrein-Related Peptidases in Nervous System Disorders. Front. Cell Neurosci. 2020, 14, 166. [Google Scholar] [CrossRef] [PubMed]
- Passamonti, L.; Tsvetanov, K.A.; Jones, P.S.; Bevan-Jones, W.R.; Arnold, R.; Borchert, R.J.; Mak, E.; Su, L.; O’Brien, J.T.; Rowe, J.B. Neuroinflammation and Functional Connectivity in Alzheimer’s Disease: Interactive Influences on Cognitive Performance. J. Neurosci. 2019, 39, 7218–7226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.Y.; Kim, S.J.; Sim, S.J.; Lee, H.J. Development of quantitative analytical method for isoflavonoid compounds from fruits of Cudraniatricuspidata. J. Korean Wood Sci. Technol. 2016, 44, 337–349. [Google Scholar] [CrossRef]
- Barnes, C.A. Memory deficits associated with senescence: A neurophysiological and behavioral study in the rat. J. Comp. Physiol. Psychol. 1979, 93, 74–104. [Google Scholar] [CrossRef]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Contextual and Cued Fear Conditioning Test Using a Video Analyzing System in Mice. JoVE J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [Green Version]
- Anagnostaras, S.G.; Wood, S.C.; Shuman, T.; Cai, D.J.; Leduc, A.D.; Zurn, K.R.; Zurn, J.B.; Sage, J.R.; Herrera, G.M. Automated assessment of pavlovian conditioned freezing and shock reactivity in mice using the video freeze system. Front. Behav. Neurosci. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Hoff, J. Methods of blood collection in the mouse. Lab. Anim. 2000, 29, 47–53. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jee, S.-C.; Lee, K.M.; Kim, M.; Lee, Y.-J.; Kim, S.; Park, J.-O.; Sung, J.-S. Neuroprotective Effect of Cudrania tricuspidata Fruit Extracts on Scopolamine-Induced Learning and Memory Impairment. Int. J. Mol. Sci. 2020, 21, 9202. https://doi.org/10.3390/ijms21239202
Jee S-C, Lee KM, Kim M, Lee Y-J, Kim S, Park J-O, Sung J-S. Neuroprotective Effect of Cudrania tricuspidata Fruit Extracts on Scopolamine-Induced Learning and Memory Impairment. International Journal of Molecular Sciences. 2020; 21(23):9202. https://doi.org/10.3390/ijms21239202
Chicago/Turabian StyleJee, Seung-Cheol, Kwang Min Lee, Min Kim, Yoo-Jung Lee, Soee Kim, Joon-Oh Park, and Jung-Suk Sung. 2020. "Neuroprotective Effect of Cudrania tricuspidata Fruit Extracts on Scopolamine-Induced Learning and Memory Impairment" International Journal of Molecular Sciences 21, no. 23: 9202. https://doi.org/10.3390/ijms21239202
APA StyleJee, S. -C., Lee, K. M., Kim, M., Lee, Y. -J., Kim, S., Park, J. -O., & Sung, J. -S. (2020). Neuroprotective Effect of Cudrania tricuspidata Fruit Extracts on Scopolamine-Induced Learning and Memory Impairment. International Journal of Molecular Sciences, 21(23), 9202. https://doi.org/10.3390/ijms21239202