Insulin Modulates the Bioenergetic and Thermogenic Capacity of Rat Brown Adipocytes In Vivo by Modulating Mitochondrial Mosaicism

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Transmission Electron Microscopy
2.2. Expression of Electron Transport Chain Components in MEF
2.3. UCP1 Expression in MEF
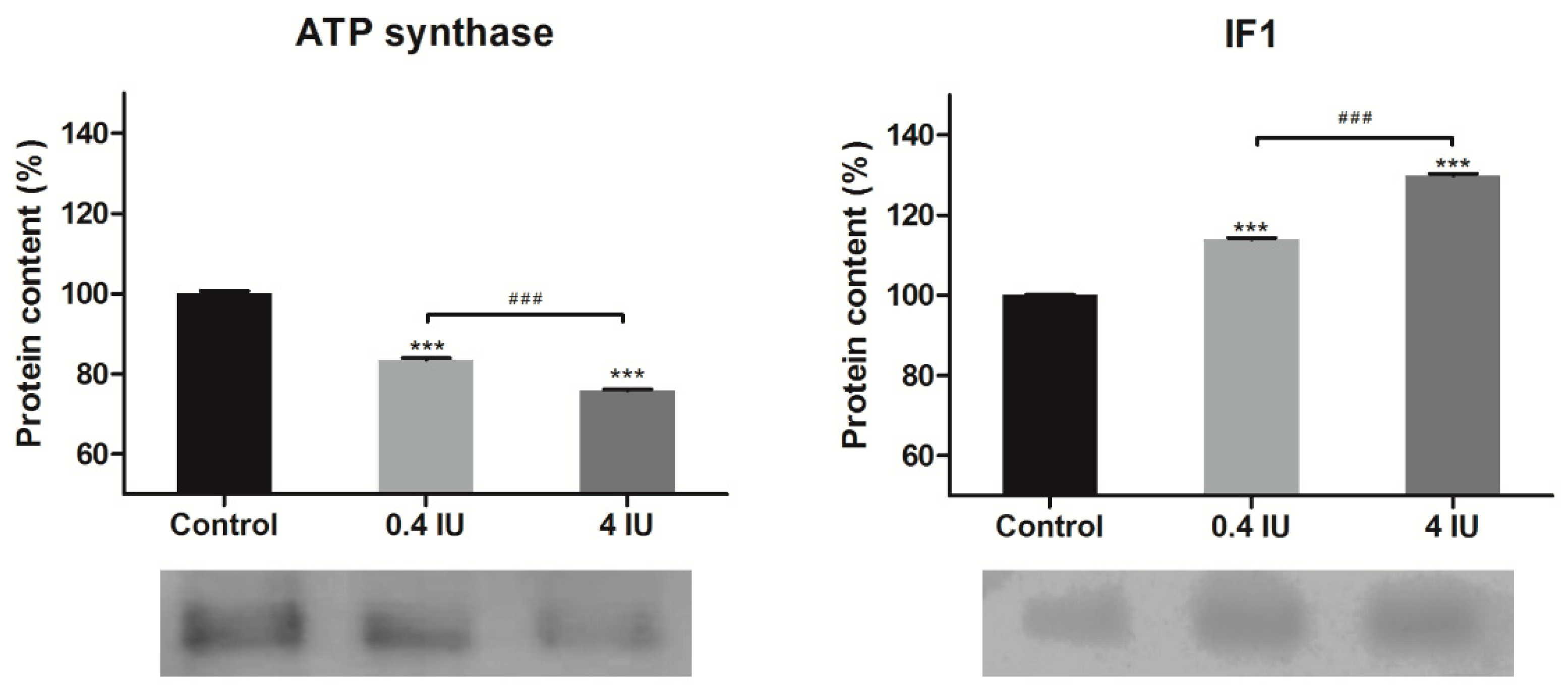
2.4. Expression of ATP Synthase and IF1 in MEF
2.5. Complex IV and ATP Synthase Activity
2.6. Mitochondrial Protein Stoichiometry
2.7. Immunofluorescence of ETC Complexes and UCP1
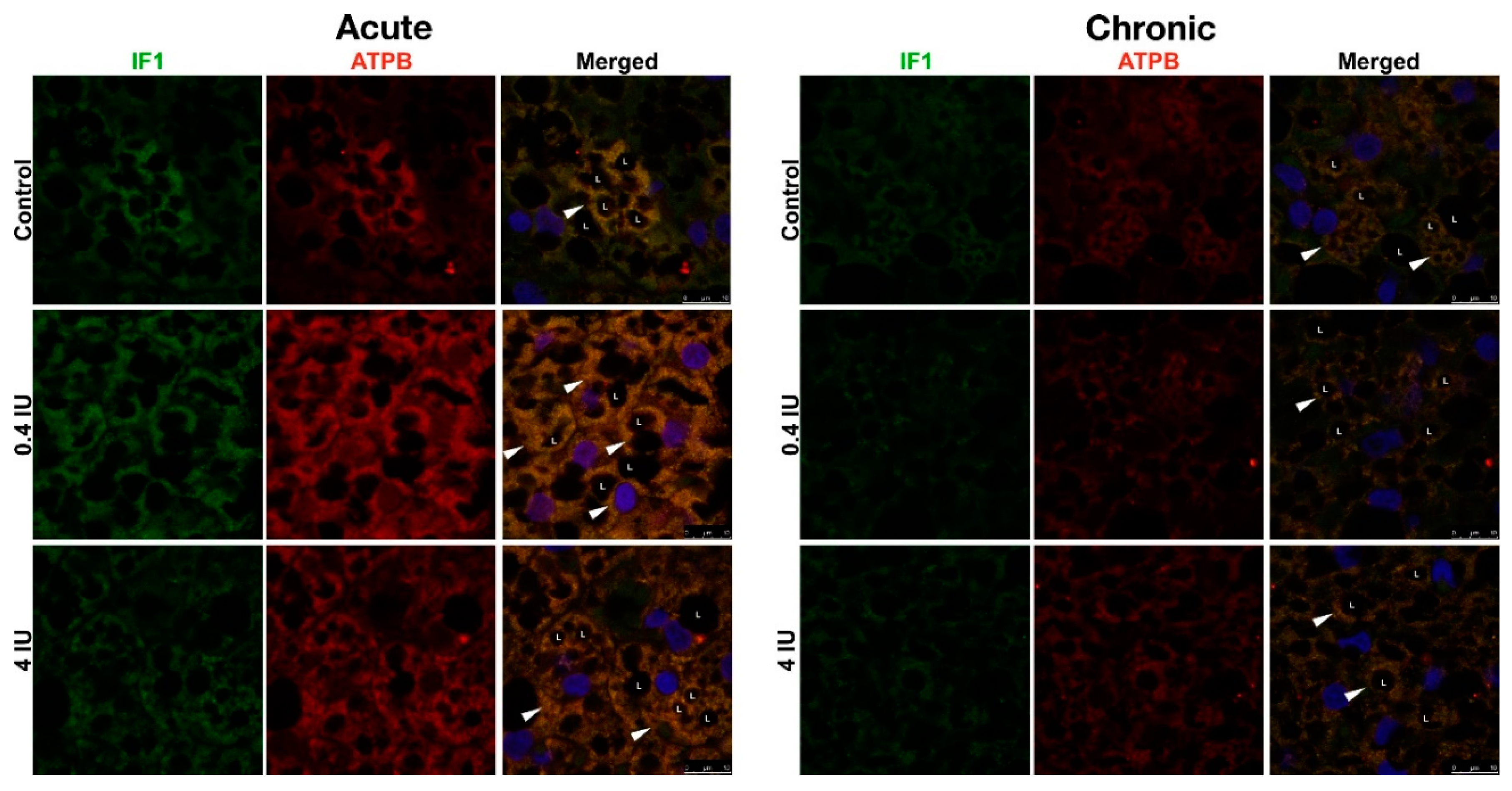
2.8. Immunofluorescence of ATP Synthase/UCP1 and IF1/UCP1
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Isolation of Mitochondria-Enriched Fraction
4.3. Western Blotting
4.4. Determination of Mitochondrial Protein Stoichiometry
4.5. Immunofluorescence
4.6. Colocalization Analysis
4.7. Transmission Electron Microscopy
4.8. Complex IV Activity Assay
4.9. ATP Synthase Activity Assay
4.10. Chemicals
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BAT | Brown adipose tissue |
| BSA | Bovine Serum Albumin |
| EDTA | Ethylenediaminetetraacetic acid |
| ETC | Electron transport chain |
| IF1 | Anti-ATPase inhibitory factor 1 |
| IU | International unit |
| MEF | Mitochondria-enriched fraction |
| NADH | Nicotinamide adenine dinucleotide |
| ROS | Reactive oxygen species |
| UCP1 | Uncoupling protein 1 |
References
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Shore, A.M.; Karamitri, A.; Kemp, P.; Speakman, J.R.; Graham, N.S.; Lomax, M.A. Cold-Induced Changes in Gene Expression in Brown Adipose Tissue, White Adipose Tissue and Liver. PLoS ONE 2013, 8, e68933. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, K.; Bettaieb, A.; Nagata, N.; Matsuo, I.; Keilhack, H.; Haj, F.G. Regulation of Brown Fat Adipogenesis by Protein Tyrosine Phosphatase 1B. PLoS ONE 2011, 6, e16446. [Google Scholar] [CrossRef] [Green Version]
- Mur, C.; Arribas, M.; Benito, M.; Valverde, A.M. Essential Role of Insulin-Like Growth Factor I Receptor in Insulin-Induced Fetal Brown Adipocyte Differentiation. Endocrinology 2003, 144, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Rothwell, N.J.; Stock, M.J. A role for insulin in the diet-induced thermogenesis of cafeteria-fed rats. Metabolism 1981, 30, 673–678. [Google Scholar] [CrossRef]
- Orava, J.; Nuutila, P.; Lidell, M.E.; Oikonen, V.; Noponen, T.; Viljanen, T.; Scheinin, M.; Taittonen, M.; Niemi, T.; Enerbäck, S.; et al. Different Metabolic Responses of Human Brown Adipose Tissue to Activation by Cold and Insulin. Cell Metabol. 2011, 14, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Stanford, K.I.; Middelbeek, R.J.W.; Townsend, K.L.; An, D.; Nygaard, E.B.; Hitchcox, K.M.; Markan, K.R.; Nakano, K.; Hirshman, M.F.; Tseng, Y.-H.; et al. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity. J. Clin. Investig. 2013, 123, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Atgie, C.; Marette, A.; Desautels, M.; Tulp, O.; Bukowiecki, L.J. Specific decrease of mitochondrial thermogenic capacity in brown adipose tissue of obese SHR/N-cp rats. Am. J. Physiol. Cell Physiol. 1993, 265, C1674–C1680. [Google Scholar] [CrossRef]
- Marette, A.; Deshaies, Y.; Collet, A.J.; Tulp, O.; Bukowiecki, L.J. Major thermogenic defect associated with insulin resistance in brown adipose tissue of obese diabetic SHR/N-cp rats. Am. J. Physiol. Endocrinol. Metab. 1991, 261, E204–E213. [Google Scholar] [CrossRef]
- Martin, S.D.; Morrison, S.; Konstantopoulos, N.; McGee, S.L. Mitochondrial dysfunction has divergent, cell type-dependent effects on insulin action. Mol. Metab. 2014, 3, 408–418. [Google Scholar] [CrossRef]
- Von Ballmoos, C.; Wiedenmann, A.; Dimroth, P. Essentials for ATP Synthesis by F1 F0 ATP Synthases. Annu. Rev. Biochem. 2009, 78, 649–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingenberg, M. UCP1—A sophisticated energy valve. Biochimie 2017, 134, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S.; Cancello, R.; Zingaretti, M.C.; Ceresi, E.; Matteis, R.D.; Giordano, A.; Himms-Hagen, J.; Ricquier, D. CL316,243 and Cold Stress Induce Heterogeneous Expression of UCP1 mRNA and Protein in Rodent Brown Adipocytes. J. Histochem. Cytochem. 2002, 50, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benador, I.Y.; Veliova, M.; Mahdaviani, K.; Petcherski, A.; Wikstrom, J.D.; Assali, E.A.; Acín-Pérez, R.; Shum, M.; Oliveira, M.F.; Cinti, S.; et al. Mitochondria Bound to Lipid Droplets Have Unique Bioenergetics, Composition, and Dynamics that Support Lipid Droplet Expansion. Cell Metabol. 2018, 27, 869–885.e6. [Google Scholar] [CrossRef] [Green Version]
- Kacerovsky, M.; Brehm, A.; Chmelik, M.; Schmid, A.I.; Szendroedi, J.; Kacerovsky-Bielesz, G.; Nowotny, P.; Lettner, A.; Wolzt, M.; Jones, J.G.; et al. Impaired insulin stimulation of muscular ATP production in patients with type 1 diabetes: Mitochondria in type 1 diabetes. J. Intern. Med. 2011, 269, 189–199. [Google Scholar] [CrossRef]
- Morrow, R.M.; Picard, M.; Derbeneva, O.; Leipzig, J.; McManus, M.J.; Gouspillou, G.; Barbat-Artigas, S.; Dos Santos, C.; Hepple, R.T.; Murdock, D.G.; et al. Mitochondrial energy deficiency leads to hyperproliferation of skeletal muscle mitochondria and enhanced insulin sensitivity. Proc. Natl. Acad. Sci. USA 2017, 114, 2705–2710. [Google Scholar] [CrossRef] [Green Version]
- Szendroedi, J.; Phielix, E.; Roden, M. The role of mitochondria in insulin resistance and type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2012, 8, 92–103. [Google Scholar] [CrossRef]
- Benard, G.; Rossignol, R. Ultrastructure of the Mitochondrion and Its Bearing on Function and Bioenergetics. Antioxid. Redox Signal. 2008, 10, 1313–1342. [Google Scholar] [CrossRef]
- Hall, J.C.; Sordahl, L.A.; Stefko, P.L. The Effect of Insulin on Oxidative Phosphorylation in Normal and Diabetic Mitochondria. J. Biol. Chem. 1960, 235, 1536–1539. [Google Scholar]
- Nisr, R.B.; Affourtit, C. Insulin acutely improves mitochondrial function of rat and human skeletal muscle by increasing coupling efficiency of oxidative phosphorylation. BBA-Bioenerg. 2014, 1837, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Stump, C.S.; Short, K.R.; Bigelow, M.L.; Schimke, J.M.; Nair, K.S. Effect of insulin on human skeletal muscle mitochondrial ATP production, protein synthesis, and mRNA transcripts. Proc. Natl. Acad. Sci. USA 2003, 100, 7996–8001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boirie, Y.; Short, K.R.; Ahlman, B.; Charlton, M.; Nair, K.S. Tissue-Specific Regulation of Mitochondrial and Cytoplasmic Protein Synthesis Rates by Insulin. Diabetes 2001, 50, 2652–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.-Y.; Cao, S.Y.; Hong, T.; Han, J.; Liu, Z.; Cao, W. Insulin Is a Stronger Inducer of Insulin Resistance than Hyperglycemia in Mice with Type 1 Diabetes Mellitus (T1DM). J. Biol. Chem. 2009, 284, 27090–27100. [Google Scholar] [CrossRef] [Green Version]
- White, M.F. Insulin Signaling in Health and Disease. Science 2003, 302, 1710–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, M.K.; Turner, N. Mitochondrial dysfunction and insulin resistance: An update. Endocr. Connect. 2015, 4, R1–R15. [Google Scholar] [CrossRef] [Green Version]
- Markelic, M.; Velickovic, K.; Golic, I.; Klepal, W.; Otasevic, V.; Stancic, A.; Jankovic, A.; Vucetic, M.; Buzadzic, B.; Korac, B.; et al. The origin of lipofuscin in brown adipocytes of hyperinsulinaemic rats: The role of lipid peroxidation and iron. Histol. Histopathol. 2013, 28, 11. [Google Scholar]
- Korać, A.; Radovanović, J.; Davidović, V.; Koko, V.; Nedeljković, M. Apoptosis in the rat brown adipose tissue after insulin treatment. J. Therm. Biol. 1999, 24, 461–464. [Google Scholar] [CrossRef]
- Klein, J.; Fasshauer, M.; Klein, H.H.; Benito, M.; Kahn, C.R. Novel adipocyte lines from brown fat: A model system for the study of differentiation, energy metabolism, and insulin action. Bioessays 2002, 24, 382–388. [Google Scholar] [CrossRef]
- Inokuma, K.-I.; Ogura-Okamatsu, Y.; Toda, C.; Kimura, K.; Yamashita, H.; Saito, M. Uncoupling Protein 1 Is Necessary for Norepinephrine-Induced Glucose Utilization in Brown Adipose Tissue. Diabetes 2005, 54, 1385–1391. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Mahadev, K.; Zilbering, A.; Zhu, L.; Goldstein, B.J. Insulin-stimulated Hydrogen Peroxide Reversibly Inhibits Protein-tyrosine Phosphatase 1B in Vivo and Enhances the Early Insulin Action Cascade. J. Biol. Chem. 2001, 276, 21938–21942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pi, J.; Bai, Y.; Zhang, Q.; Wong, V.; Floering, L.M.; Daniel, K.; Reece, J.M.; Deeney, J.T.; Andersen, M.E.; Corkey, B.E.; et al. Reactive Oxygen Species as a Signal in Glucose-Stimulated Insulin Secretion. Diabetes 2007, 56, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Yang, Y.; Zhang, S.; Kahn, A.M. Insulin-Stimulated Hydrogen Peroxide Increases Guanylate Cyclase Activity in Vascular Smooth Muscle. Hypertension 2003, 42, 569–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, C.L.; Orr, A.L.; Perevoshchikova, I.V.; Treberg, J.R.; Ackrell, B.A.; Brand, M.D. Mitochondrial Complex II Can Generate Reactive Oxygen Species at High Rates in Both the Forward and Reverse Reactions. J. Biol. Chem. 2012, 287, 27255–27264. [Google Scholar] [CrossRef] [Green Version]
- Lambert, A.J.; Boysen, H.M.; Buckingham, J.A.; Yang, T.; Podlutsky, A.; Austad, S.N.; Kunz, T.H.; Buffenstein, R.; Brand, M.D. Low rates of hydrogen peroxide production by isolated heart mitochondria associate with long maximum lifespan in vertebrate homeotherms. Aging Cell 2007, 6, 607–618. [Google Scholar] [CrossRef]
- Lee, S.; Tak, E.; Lee, J.; Rashid, M.; Murphy, M.P.; Ha, J.; Kim, S.S. Mitochondrial H2O2 generated from electron transport chain complex I stimulates muscle differentiation. Cell Res. 2011, 21, 817–834. [Google Scholar] [CrossRef] [Green Version]
- Persiyantseva, N.A.; Storozhevykh, T.P.; Senilova, Y.E.; Gorbacheva, L.R.; Pinelis, V.G.; Pomytkin, I.A. Mitochondrial H2O2 as an enable signal for triggering autophosphorylation of insulin receptor in neurons. JMS 2013, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Kramarova, T.V.; Shabalina, I.G.; Andersson, U.; Westerberg, R.; Carlberg, I.; Houstek, J.; Nedergaard, J.; Cannon, B. Mitochondrial ATP synthase levels in brown adipose tissue are governed by the c-Fo subunit P1 isoform. FASEB J. 2008, 22, 55–63. [Google Scholar] [CrossRef]
- Schwerzmann, K.; Pedersen, P.L. Regulation of the mitochondrial ATP synthase/ATPase complex. Arch. Biochem. Biophys. 1986, 250, 1–18. [Google Scholar] [CrossRef]
- Forner, F.; Kumar, C.; Luber, C.A.; Fromme, T.; Klingenspor, M.; Mann, M. Proteome Differences between Brown and White Fat Mitochondria Reveal Specialized Metabolic Functions. Cell Metabol. 2009, 10, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Campanella, M.; Parker, N.; Tan, C.H.; Hall, A.M.; Duchen, M.R. IF1: Setting the pace of the F1Fo-ATP synthase. Trends Biochem. Sci. 2009, 34, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Paumard, P.; Vaillier, J.; Coulary, B.; Schaeffer, J.; Soubannier, V.; Mueller, D.M.; Brèthes, D.; di Rago, J.-P.; Velours, J. The ATP synthase is involved in generating mitochondrial cristae morphology. EMBO J. 2002, 21, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Barbato, S.; Sgarbi, G.; Gorini, G.; Baracca, A.; Solaini, G. The Inhibitor Protein (IF1) of the F1 F0 -ATPase Modulates Human Osteosarcoma Cell Bioenergetics. J. Biol. Chem. 2015, 290, 6338–6348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claus, C.; Schonefeld, K.; Hubner, D.; Chey, S.; Reibetanz, U.; Liebert, U.G. Activity Increase in Respiratory Chain Complexes by Rubella Virus with Marginal Induction of Oxidative Stress. J. Virol. 2013, 87, 8481–8492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havlíčková Karbanová, V.; Čížková Vrbacká, A.; Hejzlarová, K.; Nůsková, H.; Stránecký, V.; Potocká, A.; Kmoch, S.; Houštěk, J. Compensatory upregulation of respiratory chain complexes III and IV in isolated deficiency of ATP synthase due to TMEM70 mutation. BBA-Bioenerg. 2012, 1817, 1037–1043. [Google Scholar] [CrossRef] [Green Version]
- Yamamori, T.; Yasui, H.; Yamazumi, M.; Wada, Y.; Nakamura, Y.; Nakamura, H.; Inanami, O. Ionizing radiation induces mitochondrial reactive oxygen species production accompanied by upregulation of mitochondrial electron transport chain function and mitochondrial content under control of the cell cycle checkpoint. Free Radic. Biol. Med. 2012, 53, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Matthias, A.; Ohlson, K.B.E.; Fredriksson, J.M.; Jacobsson, A.; Nedergaard, J.; Cannon, B. Thermogenic Responses in Brown Fat Cells Are Fully UCP1-dependent: UCP2 or UCP3 do not substitute for UCP1 in adrenergically or fatty acid-induced thermogenesis. J. Biol. Chem. 2000, 275, 25073–25081. [Google Scholar] [CrossRef] [Green Version]
- Geloen, A.; Trayhurn, P. Regulation of the level of uncoupling protein in brown adipose tissue by insulin. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1990, 258, R418–R424. [Google Scholar] [CrossRef]
- Dallon, B.W.; Parker, B.A.; Hodson, A.E.; Tippetts, T.S.; Harrison, M.E.; Appiah, M.M.A.; Witt, J.E.; Gibbs, J.L.; Gray, H.M.; Sant, T.M.; et al. Insulin selectively reduces mitochondrial uncoupling in brown adipose tissue in mice. Biochem. J. 2018, 475, 561–569. [Google Scholar] [CrossRef]
- Collins, T.J. Mitochondria are morphologically and functionally heterogeneous within cells. EMBO J. 2002, 21, 1616–1627. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, A.; Margreiter, R. Heterogeneity of Mitochondria and Mitochondrial Function within Cells as Another Level of Mitochondrial Complexity. IJMS 2009, 10, 1911–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikstrom, J.D.; Katzman, S.M.; Mohamed, H.; Twig, G.; Graf, S.A.; Heart, E.; Molina, A.J.A.; Corkey, B.E.; de Vargas, L.M.; Danial, N.N.; et al. Cell Mitochondria Exhibit Membrane Potential Heterogeneity That Can Be Altered by Stimulatory or Toxic Fuel Levels. Diabetes 2007, 56, 2569–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, A.; Dai, W.; Jang, M.J.; Medrano, L.; Li, Z.; Zhao, H.; Shao, M.; Tan, J.; Li, A.; Ning, T.; et al. Low- and high-thermogenic brown adipocyte subpopulations coexist in murine adipose tissue. J. Clin. Investig. 2019, 130, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Golic, I.; Aleksic, M.; Lazarevic, A.; Bogdanovic, M.; Jonic, S.; Korac, A. Methods for studying the localization of mitochondrial complexes III and IV by immunofluorescent and immunogold microscopy. Arch. Biol. Sci. 2016, 68, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Smith, L. Spectrophotometric Assay of Cytochrome c Oxidase. In Methods of Biochemical Analysis; Glick, D., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1955; Volume 2, pp. 427–434. ISBN 978-0-471-30459-3. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acute Treatment | |||||||||
| Control | 0.4 IU | 4 IU | |||||||
| ATP synthase | IF1 | UCP1 | ATP synthase | IF1 | UCP1 | ATP synthase | IF1 | UCP1 | |
| IF1 | 29.97 | 1 | 0.80 | 23.57 | 1 | 0.88 | 25.93 | 1 | 0.86 |
| UCP1 | 37.65 | 1.26 | 1 | 26.88 | 1.14 | 1 | 30.30 | 1.17 | 1 |
| Chronic treatment | |||||||||
| Control | 0.4 IU | 4 IU | |||||||
| ATP synthase | IF1 | UCP1 | ATP synthase | IF1 | UCP1 | ATP synthase | IF1 | UCP1 | |
| IF1 | 26.84 | 1 | 0.80 | 34.55 | 1 | 0.84 | 38.65 | 1 | 0.82 |
| UCP1 | 33.36 | 1.24 | 1 | 41.05 | 1.19 | 1 | 47.10 | 1.22 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golic, I.; Kalezic, A.; Jankovic, A.; Jonic, S.; Korac, B.; Korac, A. Insulin Modulates the Bioenergetic and Thermogenic Capacity of Rat Brown Adipocytes In Vivo by Modulating Mitochondrial Mosaicism. Int. J. Mol. Sci. 2020, 21, 9204. https://doi.org/10.3390/ijms21239204
Golic I, Kalezic A, Jankovic A, Jonic S, Korac B, Korac A. Insulin Modulates the Bioenergetic and Thermogenic Capacity of Rat Brown Adipocytes In Vivo by Modulating Mitochondrial Mosaicism. International Journal of Molecular Sciences. 2020; 21(23):9204. https://doi.org/10.3390/ijms21239204
Chicago/Turabian StyleGolic, Igor, Andjelika Kalezic, Aleksandra Jankovic, Slavica Jonic, Bato Korac, and Aleksandra Korac. 2020. "Insulin Modulates the Bioenergetic and Thermogenic Capacity of Rat Brown Adipocytes In Vivo by Modulating Mitochondrial Mosaicism" International Journal of Molecular Sciences 21, no. 23: 9204. https://doi.org/10.3390/ijms21239204
APA StyleGolic, I., Kalezic, A., Jankovic, A., Jonic, S., Korac, B., & Korac, A. (2020). Insulin Modulates the Bioenergetic and Thermogenic Capacity of Rat Brown Adipocytes In Vivo by Modulating Mitochondrial Mosaicism. International Journal of Molecular Sciences, 21(23), 9204. https://doi.org/10.3390/ijms21239204