Statistical Optimisation of Phenol Degradation and Pathway Identification through Whole Genome Sequencing of the Cold-Adapted Antarctic Bacterium, Rhodococcus sp. Strain AQ5-07



 ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Plackett-Burman Design
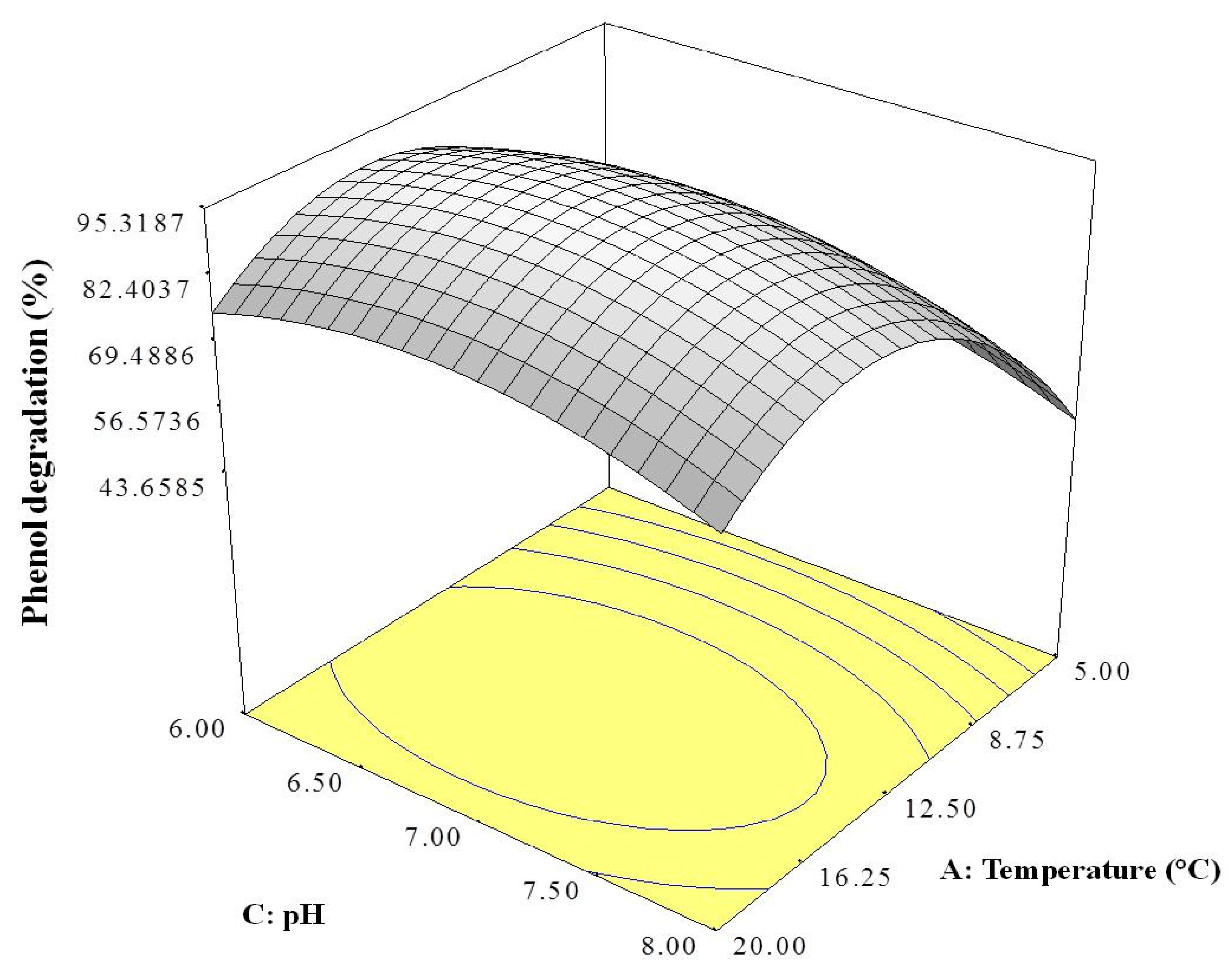
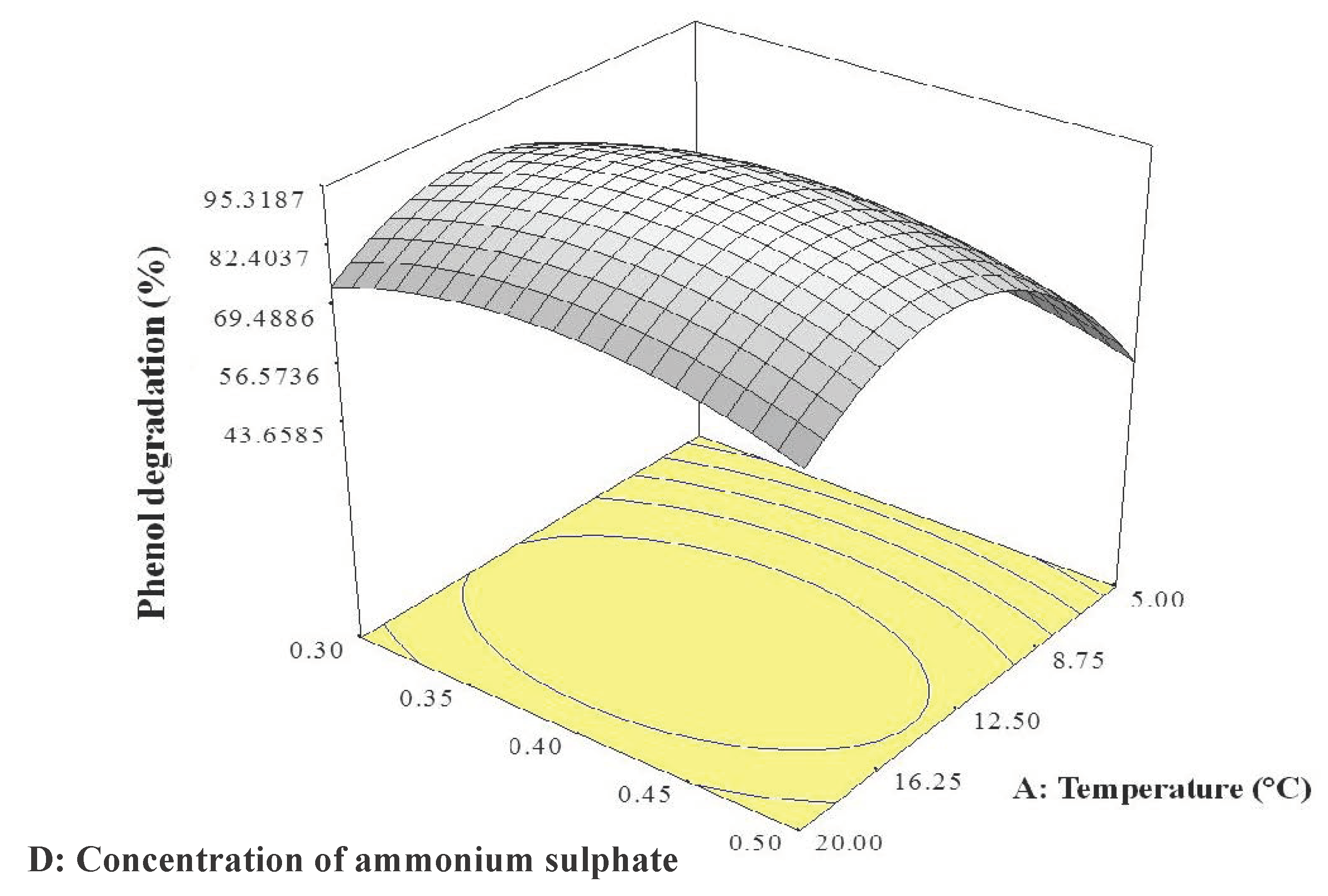
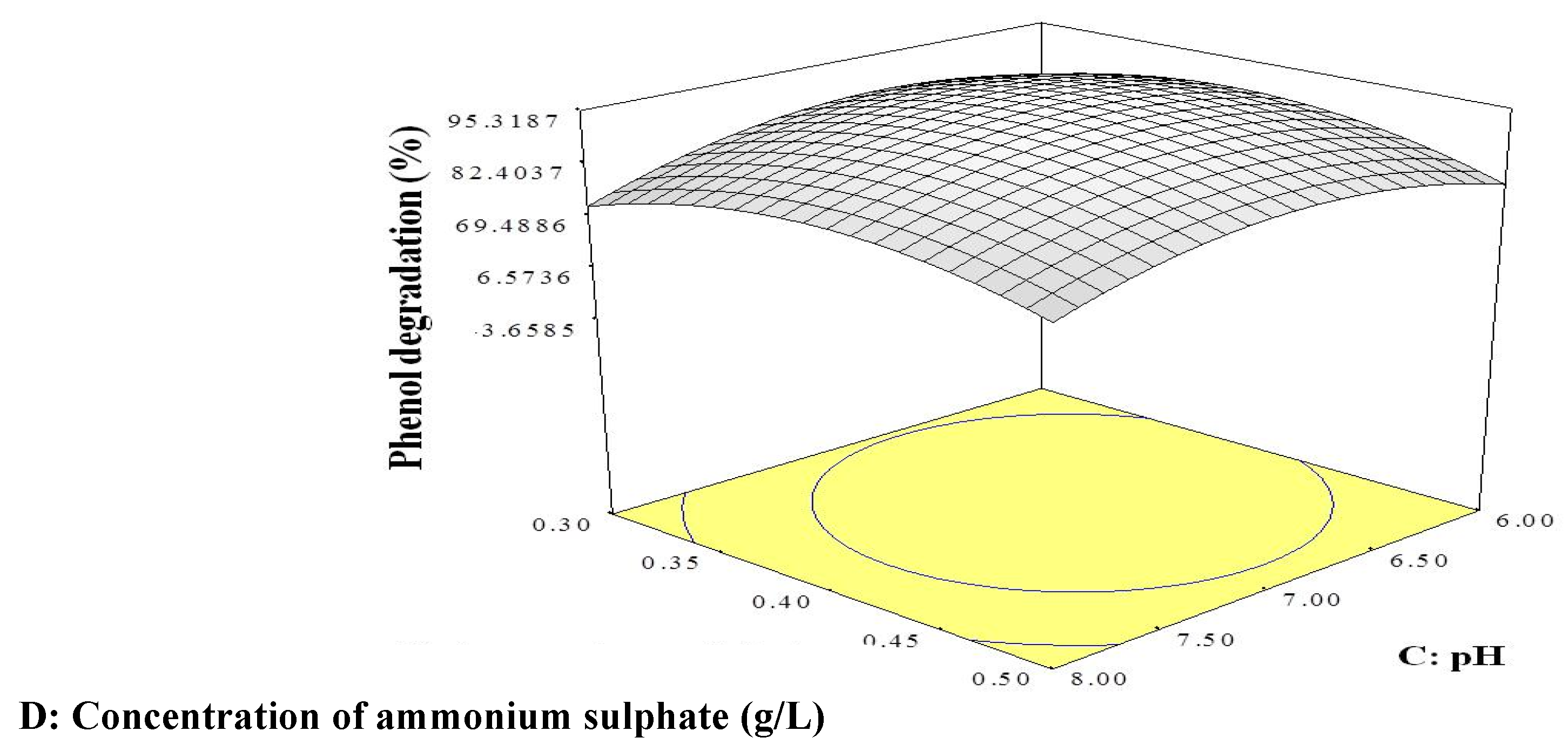
2.2. CCD
2.3. Validation Experiment
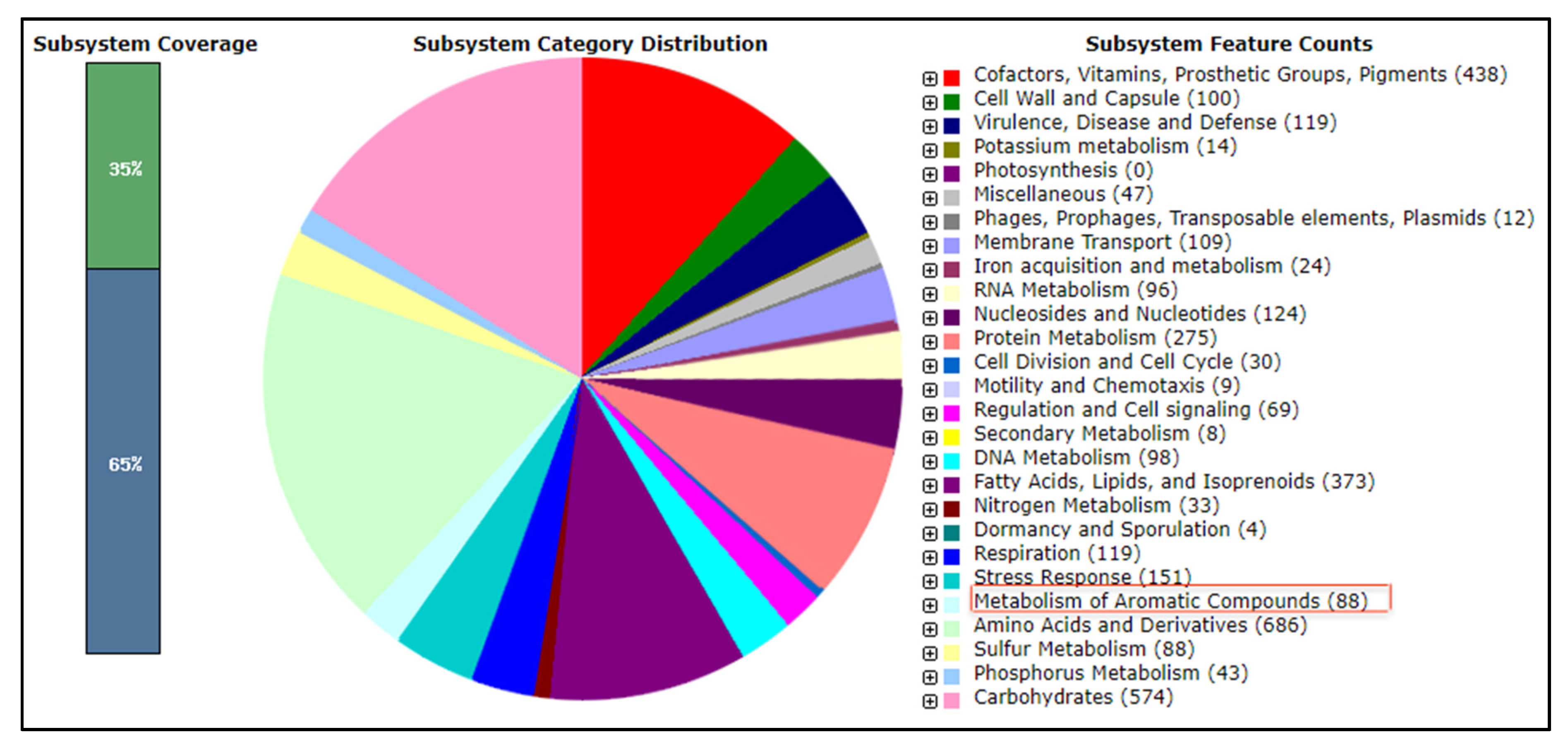
2.4. Genomic Features of Rhodococcus sp. Strain AQ5-07
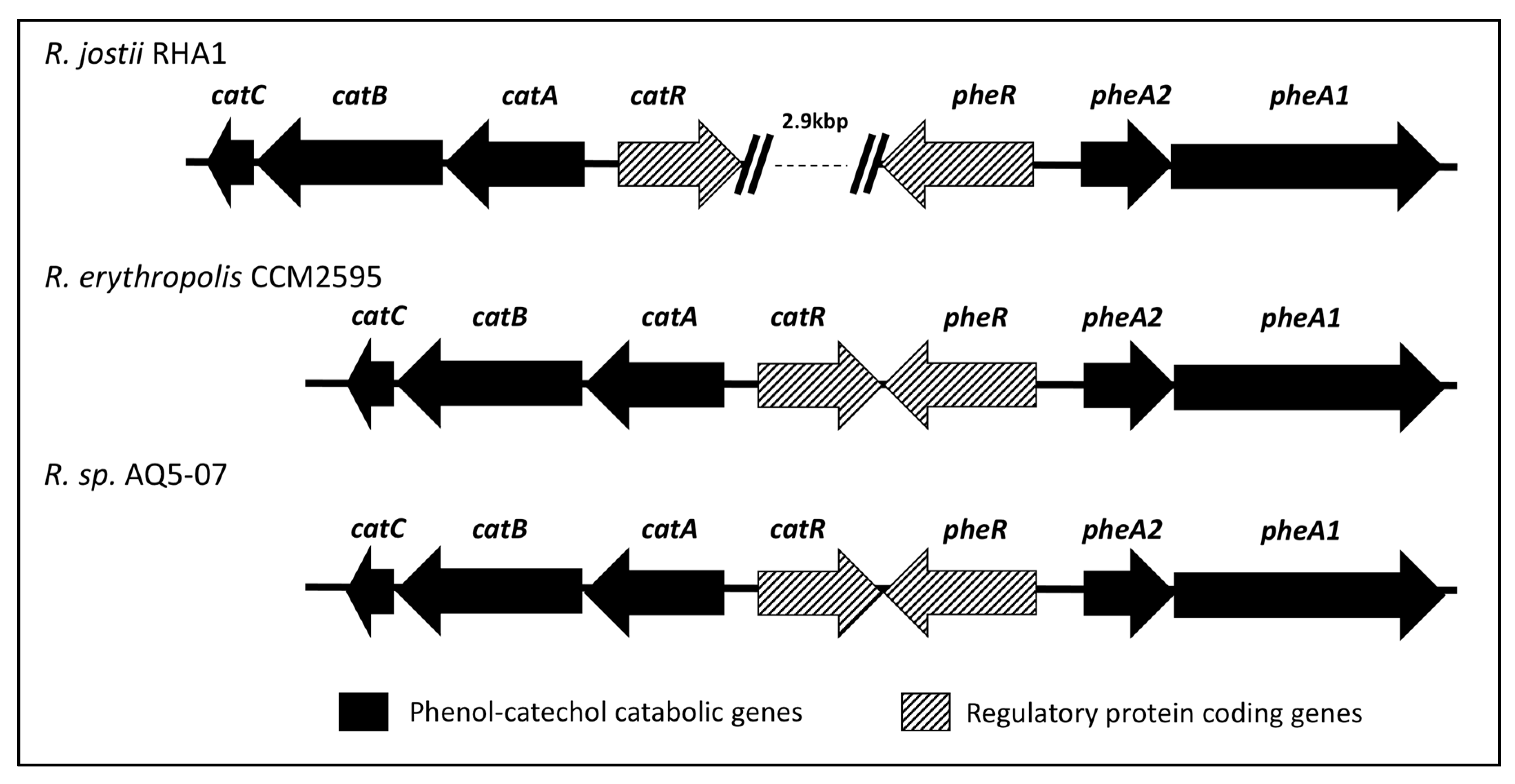
2.5. Identification of Gene Candidates for Phenol Degradation
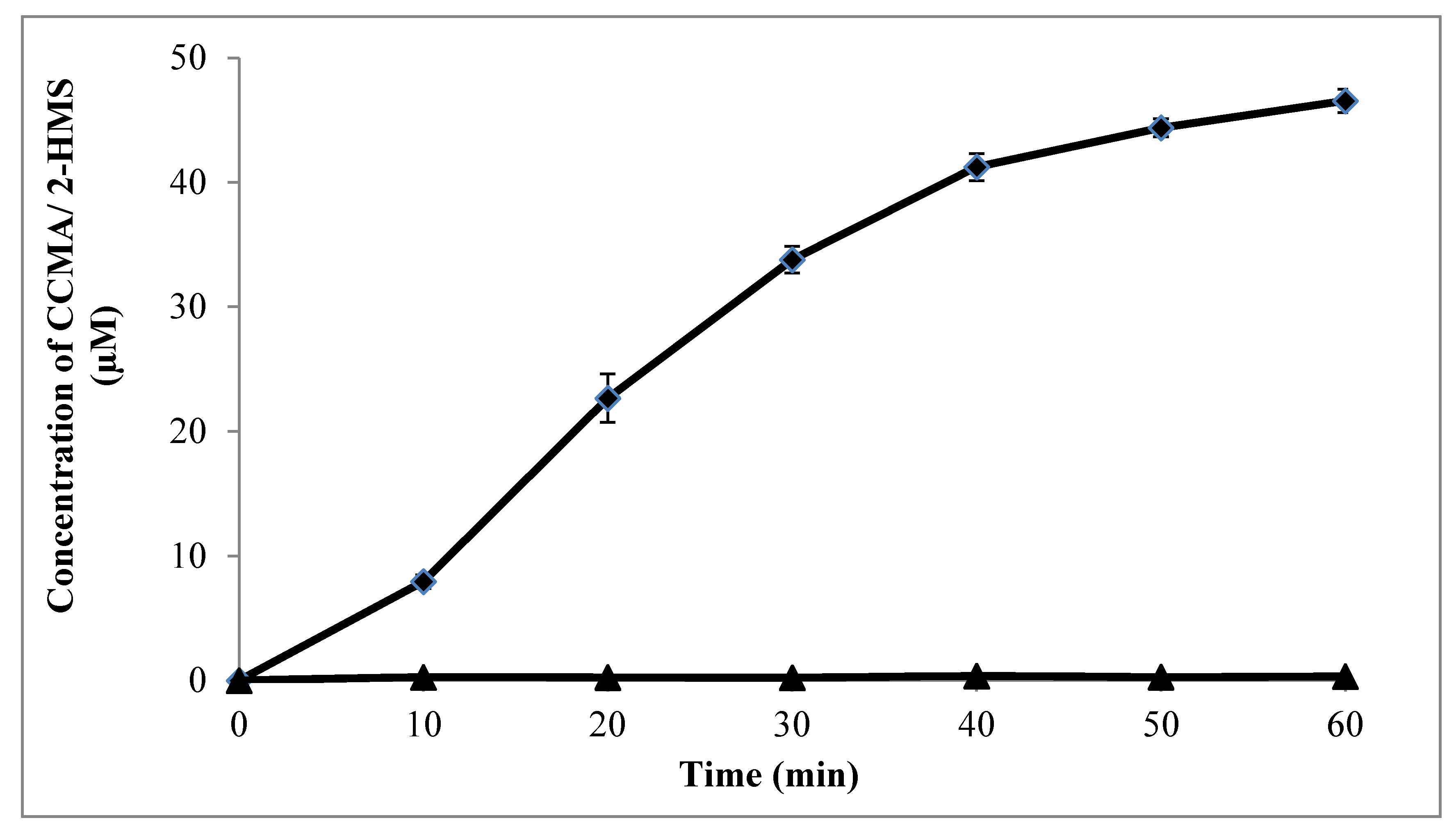
2.6. Enzymatic Assay for catechol 1,2-dioxygenase (C12D) and catechol 2,3-dioxygenase (C23D)
3. Discussion
4. Materials and Methods
4.1. Strain and Phenol Medium
4.2. Analytical Procedure
4.3. Optimisation Using Statistical Approach
4.3.1. Plackett–Burman Design
4.3.2. CCD
4.3.3. Validation of Experiments
4.4. Whole Genome Sequencing
4.4.1. Extraction of Genomic DNA
4.4.2. Genome Sequencing and Assembly
4.4.3. Gene Prediction and Annotation
4.5. Enzyme Assays
4.5.1. Preparation of Cell Extracts
4.5.2. Enzyme Assay of catechol 1, 2 dioxygenase (C12D)
4.5.3. Enzyme Assay of catechol 2, 3 dioxygenase (C23D)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RSM | response surface methodology |
| OFAT | one-factor-at-a-time |
| CCD | central composition design |
| WGS | whole genome sequencing |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| tRNA | transfer RNA |
| rRNA | ribosomal RNA |
| RAST | Rapid Annotations using subsystems Technology |
| BLAST | Basic Local Alignment Search Tool |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| COG | Clusters of Orthologous Groups |
| MSM | minimal salt medium |
| 2-HMS | 2-hydroxymuconic semialdehyde |
| CCMA | cis, cis–muconic acid; CDSs, coding sequences |
References
- Tin, T.; Fleming, Z.L.; Hughes, K.A.; Ainley, D.G.; Convey, P.; Moreno, C.A.; Pfeiffer, S.; Scott, J.; Snape, I. Impacts of local human activities on the Antarctic environment. Antarct. Sci. 2009, 21. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, T.A.; Aislabie, J.; Balks, M.R. Human Impacts on Soils. In The soils of Antarctica. Cham; Bockheim, J.G., Ed.; Springer International Publishing: New York, NY, USA, 2015; pp. 281–303. [Google Scholar] [CrossRef]
- Roslee, A.F.A.; Zakaria, N.N.; Convey, P.; Zulkharnain, A.; Lee, G.L.Y.; Gomez-Fuentes, C.; Ahmad, S.A. Statistical optimisation of growth conditions and diesel degradation by the Antarctic bacterium, Rhodococcus sp. strain AQ5-07. Extremophiles 2020, 24, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.; Abdul Khalil, K.; Zahri, K.N.M.; Gomez-Fuentes, C.; Convey, P.; Zulkharnain, A.; Sabri, S.; Alias, S.A.; González-Rocha, G.; Ahmad, S.A. Biosurfactant production and growth kinetics studies of the waste canola oil-degrading bacterium Rhodococcus erythropolis AQ5-07 from Antarctica. Molecules 2020, 25, 3878. [Google Scholar] [CrossRef]
- Karl, D.M. The grounding of the Bahia Paraiso: Microbial ecology of the 1989 Antarctic oil spill. Microb. Ecol. 1992, 24, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Aislabie, J.M.; Balks, M.R.; Foght, J.M.; Waterhouse, E.J. Hydrocarbon spills on Antarctic soils: Effects and management. Environ. Sci. Technol. 2004, 38, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Abdulrasheed, M.; Zakaria, N.N.; Ahmad Roslee, A.F.; Shukor, M.Y.; Zulkharnain, A.; Napis, S.; Convey, P.; Alias, S.A.; Gonzalez-Rocha, G.; Ahmad, S.A. Biodegradation of diesel oil by cold-adapted bacterial strains of Arthrobacter spp. from Antarctica. Antarct. Sci. 2020, 32, 1–13. [Google Scholar] [CrossRef]
- De Jesus, H.E.; Peixoto, R.S.; Cury, J.C.; van Elsas, J.D.; Rosado, A.S. Evaluation of soil bioremediation techniques in an aged diesel spill at the Antarctic Peninsula. Appl. Microbiol. Biotechnol. 2015, 99, 10815–10827. [Google Scholar] [CrossRef]
- Subramaniam, K.; Shaharuddin, N.A.; Tengku-Mazuki, T.A.; Zulkharnain, A.; Abdul Khalil, K.; Convey, P.; Ahmad, S.A. Statistical optimisation for enhancement of phenol biodegradation by the Antarctic soil bacterium Arthrobacter sp. strain AQ5-15 using response surface methodology. J. Environ. Biol. 2020, 41, 1560–1569. [Google Scholar] [CrossRef]
- Lana, N.B.; Berton, P.; Covaci, A.; Ciocco, N.F.; Barrera-Oro, E.; Atencio, A.; Altamirano, J.C. Fingerprint of persistent organic pollutants in tissues of Antarctic notothenioid fish. Sci. Total Environ. 2014, 499, 89–98. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Fuzisawa, S.; Motakatla, V.R.; Kobayashi, H.; Yoshida, E.; Yajima, Y.; Hoshino, T.; Choi, D. Degradation of toxic compounds at low and medium temperature conditions using isolated fungus. Clean Soil Air Water 2016, 44, 992–1000. [Google Scholar] [CrossRef] [Green Version]
- Zakaria, N.N.; Man, Z.; Zulkharnain, A.; Ahmad, S.A. Psychrotolerant biosurfactant-producing bacteria for hydrocarbon degradation: A mini review. Malays. J. Biochem. Mol. Biol. 2019, 21, 52–59. [Google Scholar]
- Ahmad, S.A.; Syed, M.A.; Arif, N.M.; Shukor, M.Y.A.; Shamaan, N.A. Isolation, identification and characterization of elevated phenol degrading Acinetobacter sp. strain AQ5NOL 1. Aust. J. Basic Appl. Sci. 2011, 5, 1035–1045. [Google Scholar]
- Subramaniam, K.; Ahmad, S.A.; Shaharuddin, N.A. Mini review on phenol biodegradation in Antarctica using native microorganisms. Asia Pac. J. Mol. Biol. Biotechnol. 2020, 28, 77–89. [Google Scholar] [CrossRef]
- Mazuki, T.A.T.; Shukor, M.Y.; Ahmad, S.A. Bioremediation of phenol in Antarctic: A mini review. Malays. J. Biochem. Mol. Biol. 2019, 22, 1–6. [Google Scholar]
- Luz, A.P.; Ciapina, E.M.P.; Gamba, R.C.; Lauretto, M.S.; Farias, E.W.C.; Bicego, M.C.; Taniguchi, S.; Montone, R.C.; Pellizari, V.H. Potential for bioremediation of hydrocarbon polluted soils in the Maritime Antarctic. Antarct. Sci. 2006, 18, 335–343. [Google Scholar] [CrossRef]
- Zahri, K.N.M.; Zulkharnain, A.; Gomez-Fuentes, C.; Sabri, S.; Ahmad, S.A. Effects of heavy metals on Antarctic bacteria cell growth kinetics and degradation of waste canola oil. J. Environ. Biol. 2020, 41, 1433–1441. [Google Scholar] [CrossRef]
- Liu, Z.; Xie, W.; Li, D.; Peng, Y.; Li, Z.; Liu, S. Biodegradation of phenol by bacteria strain Acinetobacter calcoaceticus PA isolated from phenolic wastewater. Int. J. Environ. Res. Public Health 2016, 13, 300. [Google Scholar] [CrossRef] [Green Version]
- Tengku-Mazuki, T.A.; Subramanian, K.; Zakaria, N.N.; Convey, P.; Abdul Khalil, K.; Lee, G.L.Y.; Zulkharnain, A.; Shaharuddin, N.A.; Ahmad, S.A. Optimization of phenol degradation by Antarctic bacterium Rhodococcus sp. Antarc. Sci. 2020, 1–10. [Google Scholar] [CrossRef]
- Chandana Lakshmi, M.V.V.; Sridevi, V.; Narasimha Rao, M.; Swamy, A.V.N. Optimization of phenol degradation from Pseudomonas aeruginosa (NCIM 2074) using response surface methodology. Int. J. Res. Pharm. Chem. 2011, 1, 925–935. [Google Scholar]
- Ahmad, S.A.; Asokan, G.; Yasid, N.A.; Nawawi, N.M.; Subramaniam, K.; Zakaria, N.N.; Shukor, M.Y. Effect of heavy metals on biodegradation of phenol by Antarctic bacterium: Arthrobacter bambusae strain AQ5-003. Malays. J. Biochem. Mol. Biol. 2018, 21, 47–51. [Google Scholar]
- Feller, G. Cryosphere and psychrophiles: Insights into a cold origin of life? Life 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, D.; Tiedje, J. Coping with our cold planet. Appl. Environ. Microbiol. 2008, 74, 1677–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaria, N.N.; Ahmad, S.A.; Yin, G.L.L.; Yasid, N.A.; Manogaran, M.; Nawawi, N.M.; Shukor, M.Y. Biodegradation of phenol by Antarctic bacterium Rhodococcus baikonurensis strain AQ5-001 in the presence of heavy metals. Malays. J. Biochem. Mol. Biol. 2018, 21, 29–36. [Google Scholar]
- Gerginova, M.; Manasiev, J.; Yemendzhiev, H.; Terziyska, A.; Peneva, N.; Alexieva, Z. Biodegradation of phenol by Antarctic strains of Aspergillus fumigatus. Z. Für Nat. C. 2014, 68, 384–393. [Google Scholar] [CrossRef]
- Margesin, R.; Fonteyne, P.-A.; Redl, B. Low-temperature biodegradation of high amounts of phenol by Rhodococcus spp. and Basidiomycetous yeasts. Res. Microbiol. 2005, 156, 68–75. [Google Scholar] [CrossRef]
- Subramaniam, K.; Mazuki, T.A.T.; Shukor, M.Y.; Ahmad, S.A. Isolation and optimisation of phenol degradation by Antarctic isolate using one factor at time. Malays. J. Biochem. Mol. Biol. 2019, 22, 79–86. [Google Scholar]
- Aislabie, J.; Saul, D.J.; Foght, J.M. Bioremediation of hydrocarbon-contaminated Polar soils. Extremophiles 2006, 10, 171–179. [Google Scholar] [CrossRef]
- Ibrahim, S.; Zahri, K.N.M.; Convey, P.; Abdul Khalil, K.; Gomez-Fuentes, C.; Zulkarnain, A.; Sabri, S.; Alias, S.A.; González-Rocha, G.; Ahmad, S.A. Optimisation of biodegradation conditions for waste canola oil by cold-adapted Rhodococcus sp. AQ5-07 from Antarctica. Electron. J. Biotechnol. 2020, 48, 1–12. [Google Scholar] [CrossRef]
- Lee, G.L.Y.; Ahmad, S.A.; Yasid, N.A.; Zulkharnain, A.; Convey, P.; Johari, W.L.W.; Alias, S.A.; Gonzalez-Rocha, G.; Shukor, M.Y. Biodegradation of phenol by cold-adapted bacteria from Antarctic soils. Polar Biol. 2018, 41, 553–562. [Google Scholar] [CrossRef]
- Pandimadevi, M.; Venkatesh Prabhu, M.; Vinod Kumar, V. Optimization of phenol degradation using Pseudomonas aeruginosa (MTCC 7814) by Plackett-Burman design and response surface methodology. J. Bioremed. Biodeg. 2014, 5, 261. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Ma, T.; Li, D.; Liang, F.; Liu, R.-L.; Li, G. Optimization of nutrient component for diesel oil degradation by Rhodococcus erythropolis. Mar. Pollut. Bull. 2008, 56, 1714–1718. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, X.; Ding, C.; Wang, Z.; Zhou, Q.; Pao, H.; Cai, W. Optimization of phenol degradation by Candida tropicalis Z-04 using Plackett-Burman design and response surface methodology. J. Environ. Sci. 2011, 23, 22–30. [Google Scholar] [CrossRef]
- Sivasubramanian, S.; Namasivayam, S.K.R. Statistical optimization of physical conditions for phenol degradation using effective microorganism-I. Indian J. Med. Res. 2014, 21, 14–20. [Google Scholar]
- Fernández, P.M.; Martorell, M.M.; Blaser, M.G.; Ruberto, L.A.M.; de Figueroa, L.I.C.; Mac Cormack, W.P. Phenol degradation and heavy metal tolerance of Antarctic yeasts. Extremophiles 2017, 21, 445–457. [Google Scholar] [CrossRef]
- Moyer, C.L.; Eric Collins, R.; Morita, R.Y. Psychrophiles and Psychrotrophs. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Bihari, Z. Current trends in bioremediation and biodegradation: Next-generation sequencing. J. Bioremediat. Biodegrad. 2013, 4, e138. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Jin, H.M.; Lee, S.H.; Math, R.K.; Madsen, E.L.; Jeon, C.O. Comparative genomic analysis and benzene, toluene, ethylbenzene, and o-, m, and p-xylene (btex) degradation pathways of Pseudoxanthomonas spadix BD-a59. Appl. Environ. Microbiol. 2013, 79, 663–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostka, J.E.; Teske, A.P.; Joye, S.B.; Head, I.M. The metabolic pathways and environmental controls of hydrocarbon biodegradation in marine ecosystems. Front Microbiol. 2014, 5, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehsani, E.; Jauregui, R.; Geffers, R.; Jareck, M.; Boon, N.; Pieper, D.H.; Vilchez-Vargas, R. Draft genome sequence of Rhodococcus sp. strain 311R. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goordial, J.; Raymond-Bouchard, I.; Ronholm, J.; Shapiro, N.; Woyke, T.; Whyte, L.; Bakermans, C. Improved-high-quality draft genome sequence of Rhodococcus sp. JG-3, a eurypsychrophilic Actinobacteria from Antarctic Dry Valley permafrost. Stand Genom. Sci. 2015, 10, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summer, E.J.; Liu, M.; Gill, J.J.; Grant, M.; Chan-Cortes, T.N.; Ferguson, L.; Janes, C.; Lange, K.; Bertoli, M.; Moore, C.; et al. Genomic and functional analyses of Rhodococcus equi phages ReqiPepy6, ReqiPoco6, ReqiPine5, and ReqiDocB7. Appl. Environ. Microbiol. 2011, 77, 669–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahar, A.; Baker, A.L.; Charleston, M.A.; Bowman, J.P.; Britz, M.L. Draft genome sequences of three sub-Antarctic Rhodococcus spp., including two novel psychrophilic genomospecies. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.Y.; Yin, W.-F.; Chan, K.-G. Enterobacter asburiae strain L1: Complete genome and whole genome optical mapping analysis of a quorum sensing bacterium. Sensors 2014, 14, 13913–13924. [Google Scholar] [CrossRef]
- Song, W.; Lin, X.; Huang, X. Characterization and expression analysis of three cold shock protein (CSP) genes under different stress conditions in the Antarctic bacterium Psychrobacter sp. G. Polar Biol. 2012, 35, 1515–1524. [Google Scholar] [CrossRef]
- Keto-Timonen, R.; Hietala, N.; Palonen, E.; Hakakorpi, A.; Lindström, M.; Korkeala, H. Cold shock proteins: A minireview with special emphasis on csp-family of Enteropathogenic Yersinia. Front Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Phadtare, S. Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 2004, 6, 125–136. [Google Scholar] [PubMed]
- Muñoz, P.A.; Márquez, S.L.; González-Nilo, F.D.; Márquez-Miranda, V.; Blamey, J.M. Structure and application of antifreeze proteins from Antarctic bacteria. Microb. Cell Factories 2017, 16. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Davies, P.L.; Laybourn-Parry, J. A hyperactive, Ca2+-dependent antifreeze protein in an Antarctic bacterium. FEMS Microbiol. Lett. 2005, 245, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Ghoshal, A.K. Phenol degradation performance by isolated Bacillus cereus immobilized in alginate. Int. Biodeterior. Biodegrad. 2011, 65, 1052–1060. [Google Scholar] [CrossRef]
- Martinkova, L.; Uhnakova, B.; Patek, M.; Nesvera, J.; Kren, V. Biodegradation potential of the genus Rhodococcus. Environ. Int. 2009, 35, 162–177. [Google Scholar] [CrossRef]
- Strnad, H.; Patek, M.; Fousek, J.; Szokol, J.; Ulbrich, P.; Nesvera, J.; Paces, V.; Vlcek, C. Genome sequence of Rhodococcus erythropolis strain CCM2595, a phenol derivative-degrading bacterium. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Patrauchan, M.A.; Florizone, C.; Dosanjh, M.; Mohn, W.W.; Davies, J.; Eltis, L.D. Catabolism of benzoate and phthalate in Rhodococcus sp. strain rha1: Redundancies and convergence. J. Bacteriol. 2005, 187, 4050–4063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás-Gallardo, L.; Santero, E.; Camafeita, E.; Calvo, E.; Schlömann, M.; Floriano, B. Molecular and biochemical characterization of the tetralin degradation pathway in Rhodococcus sp. strain TFB. Microb. Biotechnol. 2009, 2, 262–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Chae, J.-C.; Zylstra, G.J.; Sohn, H.-Y.; Kwon, G.-S.; Kim, E. Identification of two-component regulatory genes involved in o-xylene degradation by Rhodococcus sp. strain DK17. J. Microbiol. Seoul Korea 2005, 43, 49–53. [Google Scholar]
- Szőköl, J.; Rucká, L.; Šimčíková, M.; Halada, P.; Nešvera, J.; Pátek, M. Induction and carbon catabolite repression of phenol degradation genes in Rhodococcus erythropolis and Rhodococcus jostii. Appl. Microbiol. Biotechnol. 2014, 98, 8267–8279. [Google Scholar] [CrossRef] [PubMed]
- Veselý, M.; Knoppová, M.; Nesvera, J.; Pátek, M. Analysis of catRABC operon for catechol degradation from phenol-degrading Rhodococcus erythropolis. Appl. Microbiol. Biotechnol. 2007, 76, 159–168. [Google Scholar] [CrossRef]
- Eulberg, D.; Schlömann, M. The putative regulator of catechol catabolism in Rhodococcus opacus 1CP—An IclR-type, not a LysR-type transcriptional regulator. Antonie Van Leeuwenhoek 1998, 74, 71–82. [Google Scholar] [CrossRef]
- Jiménez, J.I.; Miñambres, B.; García, J.L.; Díaz, E. Genomic insights in the metabolism of aromatic compounds in Pseudomonas. In Pseudomonas; Springer: Boston, MA, USA, 2004; pp. 425–462. [Google Scholar] [CrossRef]
- Nojiri, H.; Maeda, K.; Sekiguchi, H.; Urata, M.; Shintani, M.; Yoshida, T.; Habe, H.; Omori, T. Organization and transcriptional characterization of catechol degradation genes involved in carbazole degradation by Pseudomonas resinovorans strain CA10. Biosci. Biotechnol. Biochem. 2002, 66, 897–901. [Google Scholar] [CrossRef] [Green Version]
- Cámara, B.; Bielecki, P.; Kaminski, F.; dos Santos, V.M.; Plumeier, I.; Nikodem, P.; Pieper, D.H. A gene cluster involved in degradation of substituted salicylates via ortho cleavage in Pseudomonas sp. strain MT1 encodes enzymes specifically adapted for transformation of 4-methylcatechol and 3-methylmuconate. J. Bacteriol. 2007, 189, 1664–1674. [Google Scholar] [CrossRef] [Green Version]
- Tropel, D.; van der Meer, J.R. Bacterial transcriptional regulators for degradation pathways of aromatic compounds. Microbiol. Mol. Biol. Rev. 2004, 68, 474–500. [Google Scholar] [CrossRef] [Green Version]
- Gröning, J.A.D.; Eulberg, D.; Tischler, D.; Kaschabek, S.R.; Schlömann, M. Gene redundancy of two-component (chloro)phenol hydroxylases in Rhodococcus opacus 1CP. FEMS Microbiol. Lett. 2014, 361, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Saa, L.; Jaureguibeitia, A.; Largo, E.; Llama, M.J.; Serra, J.L. Cloning, purification and characterization of two components of phenol hydroxylase from Rhodococcus erythropolis UPV-1. Appl. Microbiol. Biotechnol. 2010, 86, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Bej, A.K.; Saul, D.; Aislabie, J. Cold-tolerant alkane-degrading Rhodococcus species from Antarctica. Polar Biol. 2000, 23, 100–105. [Google Scholar] [CrossRef]
- de Jesus, H.E.; Peixoto, R.S.; Rosado, A.S. Bioremediation in Antarctic soils. J. Pet. Environ. Biotechnol. 2015, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Ruberto, L.; Vazquez, S.; Lobalbo, A.; Mac Cormack, W.P. Psychrotolerant hydrocarbon-degrading Rhodococcus strains isolated from polluted Antarctic soils. Antarct Sci. 2005, 17, 47–56. [Google Scholar] [CrossRef]
- Nair, C.I.; Jayachandran, K.; Shashidhar, S. Biodegradation of phenol. Afr. J. Biotechnol. 2008, 7, 4951–4958. [Google Scholar]
- Nadaf, N.H.; Ghosh, J.S. Purification and characterization of catechol 1, 2-dioxygenase from Rhodococcus sp. NCIM 289. Res. J. Environ. Earth Sci. 2011, 3, 608–613. [Google Scholar]
- Táncsics, A.; Szoboszlay, S.; Kriszt, B.; Kukolya, J.; Baka, E.; Márialigeti, K.; Révész, S. Applicability of the functional gene catechol 1,2-dioxygenase as a biomarker in the detection of BTEX-degrading Rhodococcus species. J. Appl. Microbiol. 2008, 105, 1026–1033. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Y.; Shi, Y.; Song, W.; Zhang, C. Cloning, expression and characterization of a mesophilic catechol 1,2-dioxygenase from Rhodococcus ruber OA1. Biotechnol. Faisalabad 2016, 16, 10–18. [Google Scholar] [CrossRef]
- Tavakoli, A.; Hamzah, A. Characterization and evaluation of catechol oxygenases by twelve bacteria, isolated from oil contaminated soils in Malaysia. Biol. J. Microorg. 2017, 5, 71–80. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Abdulrasheed, M.; Zulkharnain, A.; Zakaria, N.N.; Roslee, A.F.A.; Abdul Khalil, K.; Napis, S.; Convey, P.; Gomez-Fuentes, C.; Ahmad, S.A. Response surface methodology optimization and kinetics of diesel degradation by a cold-adapted Antarctic bacterium, Arthrobacter sp. strain AQ5-05. Sustainability 2020, 12, 6966. [Google Scholar] [CrossRef]
- Xie, Y.; Wu, G.; Tang, J.; Luo, R.; Patterson, J.; Liu, S.; Huang, W.; He, G.; Gu, S.; Li, S.; et al. SOAPdenovo-Trans: De novo transcriptome assembly with short RNA-Seq reads. Bioinformatics 2014, 30, 1660–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedell, J.A.; Korf, I.; Gish, W. MaskerAid: A performance enhancement to RepeatMasker. Bioinformatics 2000, 16, 1040–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Stærfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Moxon, S.; Marshall, M.; Khanna, A.; Eddy, S.R.; Bateman, A. Rfam: Annotating non-coding RNAs in complete genomes. Nucleic Acids Res. 2005, 33, D121–D124. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Margesin, R.; Bergauer, P.; Gander, S. Degradation of phenol and toxicity of phenolic compounds: A comparison of cold-tolerant Arthrobacter sp. and mesophilic Pseudomonas putida. Extremophiles 2004, 8, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Guzik, U.; Hupert-Kocurek, K.; Sitnik, M.; Wojcieszyńska, D. High activity catechol 1,2-dioxygenase from Stenotrophomonas maltophilia strain KB2 as a useful tool in cis,cis-muconic acid production. Antonie Van Leeuwenhoek 2013, 103, 1297–1307. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.A.; Shamaan, N.A.; Syed, M.A.; Khalid, A.; Ab Rahman, N.A.; Khalil, K.A.; Dahalan, F.A.; Shukor, M.Y. Meta-cleavage pathway of phenol degradation by Acinetobacter sp. strain AQ5NOL 1. Rend. Fis. Acc. Lincei 2017, 28, 1–9. [Google Scholar] [CrossRef]
- Hupert-Kocurek, K.; Guzik, U.; Wojcieszynska, D. Characterization of catechol 2, 3-dioxygenase from Planococcus sp. strain S5 induced by high phenol concentration. Acta Biochim. Pol. 2012, 59, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinaru, E.; Truu, J.; Stottmeister, U.; Heinaru, A. Three types of phenol and p-cresol catabolism in phenol- and p-cresol-degrading bacteria isolated from river water continuously polluted with phenolic compounds. FEMS Microbiol. Ecol. 2000, 31, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Yokota, T. Benzoate metabolism in Pseudomonas putida(arvilla) MT-2: Demonstration of two benzoate pathways. J. Bacteriol. 1973, 115, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Wojcieszyńska, D.; Guzik, U.; Greń, I.; Perkosz, M.; Hupert-Kocurek, K. Induction of aromatic ring: Cleavage dioxygenases in Stenotrophomonas maltophilia strain KB2 in cometabolic systems. World J. Microbiol. Biotechnol. 2011, 27, 805–811. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | A | B | C | D | Phenol Degradation (%) |
|---|---|---|---|---|---|
| 1 | 25 | 0.30 | 8 | 0.1 | 53.04 |
| 2 | 5 | 0.05 | 8 | 0.5 | 49.31 |
| 3 | 5 | 0.30 | 6 | 0.1 | 49.08 |
| 4 | 25 | 0.30 | 6 | 0.5 | 61.08 |
| 5 | 5 | 0.30 | 8 | 0.1 | 44.12 |
| 6 | 5 | 0.05 | 6 | 0.5 | 50.34 |
| 7 | 25 | 0.05 | 8 | 0.5 | 56.64 |
| 8 | 5 | 0.05 | 6 | 0.1 | 46.43 |
| 9 | 25 | 0.30 | 6 | 0.5 | 59.36 |
| 10 | 25 | 0.05 | 8 | 0.1 | 52.68 |
| 11 | 25 | 0.05 | 6 | 0.1 | 56.52 |
| 12 | 5 | 0.30 | 8 | 0.5 | 44.58 |
| Source | Sum of Squares | Degree of Freedom | Mean Square | F Value | Prob > F |
|---|---|---|---|---|---|
| Model | 344.5220 | 8 | 43.0653 | 57.7867 | 0.0034 ** |
| A | 129.9352 | 1 | 129.9352 | 174.3524 | 0.0009 *** |
| B | 3.0380 | 1 | 3.0380 | 4.0765 | 0.1368 |
| C | 37.8766 | 1 | 37.8766 | 50.8245 | 0.0057 ** |
| D | 11.7988 | 1 | 11.7988 | 15.8322 | 0.0284 * |
| AC | 2.5548 | 1 | 2.5548 | 3.4281 | 0.1612 |
| AD | 0.0870 | 1 | 0.0870 | 0.1168 | 0.7551 |
| BC | 5.6012 | 1 | 5.6012 | 7.5159 | 0.0712 |
| BD | 7.2682 | 1 | 7.2682 | 9.7528 | 0.0524 |
| Residual | 2.2357 | 3 | 0.7452 | ||
| Lack of Fit | 0.7565 | 2 | 0.3783 | 0.2557 | 0.8134 |
| Pure Error | 1.4792 | 1 | 1.4792 | ||
| Cor Total | 346.7578 | 11 | |||
| Std dev | 0.86 | R2 | 0.9936 | ||
| Mean | 51.93 | Adjusted R2 | 0.9764 | ||
| C.V | 1.66 | Predicted R2 | 0.9290 | ||
| PRESS | 24.63 | Adeq Precision | 21.1694 |
| Run Order | A | C | D | Phenol Degradation (%) | |
|---|---|---|---|---|---|
| Experimental Value | Predicted Value | ||||
| 1 | 12.5 | 7.0 | 0.40 | 93.91 | 93.45 |
| 2 | 12.5 | 7.0 | 0.40 | 94.10 | 93.45 |
| 3 | 12.5 | 7.0 | 0.23 | 69.72 | 69.42 |
| 4 | 25.0 | 7.0 | 0.40 | 47.40 | 46.21 |
| 5 | 20.0 | 6.0 | 0.50 | 63.48 | 64.11 |
| 6 | 12.5 | 7.0 | 0.40 | 92.53 | 93.45 |
| 7 | 20.0 | 8.0 | 0.30 | 60.60 | 61.10 |
| 8 | 12.5 | 5.3 | 0.40 | 73.56 | 73.10 |
| 9 | 12.5 | 8.7 | 0.40 | 62.64 | 62.60 |
| 10 | 5.0 | 8.0 | 0.30 | 33.96 | 33.69 |
| 11 | 12.5 | 7.0 | 0.57 | 67.20 | 67.00 |
| 12 | 12.5 | 7.0 | 0.40 | 93.52 | 93.45 |
| 13 | 12.5 | 7.0 | 0.40 | 93.72 | 93.45 |
| 14 | 5.0 | 6.0 | 0.30 | 43.32 | 43.23 |
| 15 | 0 | 7.0 | 0.40 | 2.76 | 3.43 |
| 16 | 20.0 | 6.0 | 0.30 | 68.76 | 69.44 |
| 17 | 12.5 | 7.0 | 0.40 | 92.82 | 93.45 |
| 18 | 5.0 | 8.0 | 0.50 | 36.48 | 36.16 |
| 19 | 20.0 | 8.0 | 0.50 | 60.84 | 61.30 |
| 20 | 5.0 | 6.0 | 0.50 | 40.32 | 40.18 |
| Source | Sum of Squares | DF | Mean Square | F Value | Prob > F |
|---|---|---|---|---|---|
| Model | 12,042.1628 | 9 | 1338.0181 | 2358.7788 | <0.0001 *** |
| A | 2233.4754 | 1 | 2233.4754 | 3937.3716 | <0.0001 *** |
| C | 131.4727 | 1 | 131.4727 | 231.7720 | <0.0001 *** |
| D | 6.9752 | 1 | 6.9752 | 12.2965 | 0.0057 ** |
| A2 | 8534.5562 | 1 | 8534.5562 | 15,045.4843 | <0.0001 *** |
| C2 | 1169.5896 | 1 | 1169.5896 | 2061.8578 | <0.0001 *** |
| D2 | 1136.9217 | 1 | 1136.9217 | 2004.2680 | <0.0001 *** |
| AC | 0.7200 | 1 | 0.7200 | 1.2693 | 0.2862 |
| AD | 2.5992 | 1 | 2.5992 | 4.5821 | 0.0580 |
| CD | 15.2352 | 1 | 15.2352 | 26.8580 | 0.0004 *** |
| Residual | 5.6725 | 10 | 0.5673 | ||
| Lack of Fit | 3.7190 | 5 | 0.7438 | 1.9037 | 0.2484 |
| Pure Error | 1.9535 | 5 | 0.3907 | ||
| Cor Total | 12,047.8353 | 19 | |||
| Std dev | 0.75 | R2 | 0.9995 | ||
| Mean | 64.58 | Adjusted R2 | 0.9991 | ||
| Coefficient of Variance | 1.17 | Predicted R2 | 0.9975 | ||
| PRESS | 30.56 | Adeq Precision | 169.0312 |
| Feature | Count/Value |
|---|---|
| Genome size (bp) | 6,749,221 |
| GC content (%) | 62.4 |
| Number of contigs | 34 |
| Length of the longest contig (bp) | 1,378,316 |
| Number of Subsystems | 436 |
| Number of coding sequences (CDSs) | 6545 |
| Number of rRNAs | 4 |
| Number of tRNAs | 75 |
| Gene Name (Locus Tag) | Gene Products | Accession No. | Amino Acid Residues (Aa) | COG No. | KEGG No. |
|---|---|---|---|---|---|
| pheA1 (S2GM001986) | Phenol hydroxylase large subunit A1 | RAL35045 | 542 | COG2368 | K03380 |
| pheA2 (S2GM001985) | Phenol hydroxylase small subunit A2 | RAL35044 | 189 | COG1853 | K03380 |
| catA (S2GM001982) | Catechol 1,2-dioxygenase | RAL35043 | 279 | COG3485 | K03381 |
| catB (S2GM001981) | Muconate cycloisomerase | RAL35564 | 373 | COG4948 | K01856 |
| catC (S2GM001980) | Muconolactone isomerase | RAL35042 | 93 | COG4829 | K03464 |
| praD (S2GM002436) | 3-oxoadipate enol-lactonase | RAL34922 | 270 | COG0596 | K01055 |
| pcaI (S2GM003023) | 3-oxoadipate coa-transferase, alpha subunit | RAL34191 | 303 | COG1788 | K01031 |
| pcaJ (S2GM003024) | 3-oxoadipate coa-transferase, beta subunit | RAL34192 | 261 | COG2057 | K01032 |
| fadA/ fadI (S2GM005247) | Acetyl-coa acyltransferase | RAL31833 | 411 | COG0183 | K00632 |
| Variables | Symbol | Unit | Experimental Value | |
|---|---|---|---|---|
| Low (−1) | High (+1) | |||
| Temperature | A | °C | 5 | 25 |
| Concentration of NaCl | B | g/L | 0.05 | 0.25 |
| pH | C | - | 6 | 8 |
| Concentration of (NH4)2SO4 | D | g/L | 0.1 | 0.5 |
| Variables | Symbol | Unit | Experimental Value | ||||
|---|---|---|---|---|---|---|---|
| −2 | −1 | 0 | +1 | +2 | |||
| Temperature | A | °C | 0 | 5.0 | 10.0 | 20.0 | 25.0 |
| pH | C | - | 5.3 | 6.0 | 7.0 | 8.0 | 8.7 |
| Concentration of (NH4)2SO4 | D | g/L | 0.23 | 0.30 | 0.40 | 0.50 | 0.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, G.L.Y.; Zakaria, N.N.; Convey, P.; Futamata, H.; Zulkharnain, A.; Suzuki, K.; Abdul Khalil, K.; Shaharuddin, N.A.; Alias, S.A.; González-Rocha, G.; et al. Statistical Optimisation of Phenol Degradation and Pathway Identification through Whole Genome Sequencing of the Cold-Adapted Antarctic Bacterium, Rhodococcus sp. Strain AQ5-07. Int. J. Mol. Sci. 2020, 21, 9363. https://doi.org/10.3390/ijms21249363
Lee GLY, Zakaria NN, Convey P, Futamata H, Zulkharnain A, Suzuki K, Abdul Khalil K, Shaharuddin NA, Alias SA, González-Rocha G, et al. Statistical Optimisation of Phenol Degradation and Pathway Identification through Whole Genome Sequencing of the Cold-Adapted Antarctic Bacterium, Rhodococcus sp. Strain AQ5-07. International Journal of Molecular Sciences. 2020; 21(24):9363. https://doi.org/10.3390/ijms21249363
Chicago/Turabian StyleLee, Gillian Li Yin, Nur Nadhirah Zakaria, Peter Convey, Hiroyuki Futamata, Azham Zulkharnain, Kenshi Suzuki, Khalilah Abdul Khalil, Noor Azmi Shaharuddin, Siti Aisyah Alias, Gerardo González-Rocha, and et al. 2020. "Statistical Optimisation of Phenol Degradation and Pathway Identification through Whole Genome Sequencing of the Cold-Adapted Antarctic Bacterium, Rhodococcus sp. Strain AQ5-07" International Journal of Molecular Sciences 21, no. 24: 9363. https://doi.org/10.3390/ijms21249363
APA StyleLee, G. L. Y., Zakaria, N. N., Convey, P., Futamata, H., Zulkharnain, A., Suzuki, K., Abdul Khalil, K., Shaharuddin, N. A., Alias, S. A., González-Rocha, G., & Ahmad, S. A. (2020). Statistical Optimisation of Phenol Degradation and Pathway Identification through Whole Genome Sequencing of the Cold-Adapted Antarctic Bacterium, Rhodococcus sp. Strain AQ5-07. International Journal of Molecular Sciences, 21(24), 9363. https://doi.org/10.3390/ijms21249363