ROS and Ions in Cell Signaling during Sexual Plant Reproduction



{kind=link}
Abstract
:1. Introduction
2. Pollen Germination
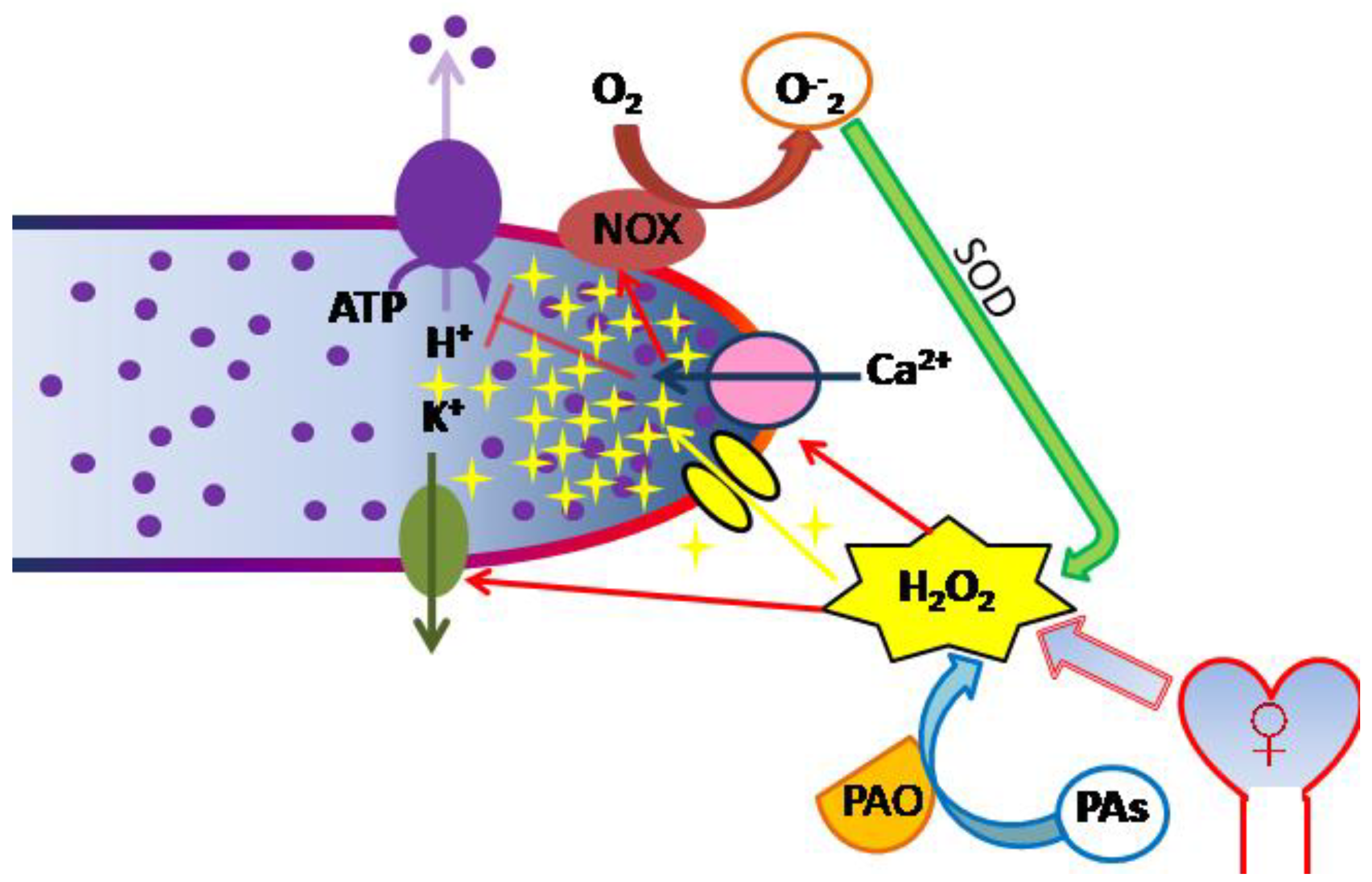
2.1. ROS Production as an Early Event during Germination
2.2. Changes of Ionic Status during Early Germination Stages
2.3. ROS Production on Stigma
2.4. Perception of Exogenous ROS Signal by Pollen
3. Pollen Tube Growth
3.1. Ionic Homeostasis and ROS Production in Growing Pollen Tube
3.2. ROS are Involved in Signal Perception and Mediate Pollen Tube Rupture
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | reactive oxygen species |
| ROP | Rho of plants |
| MnTMPP | Mn-5,10,15,20-tetrakis(1-methyl-4-pyridyl)21H,23H-porphin |
| CMS | cytoplasmic male sterility |
| DPI | diphenyl iodonium chloride |
| NBT | nitro blue tetrazolium |
| SOD | superoxide dismutase |
| NOX | NADPH-oxidase |
| MP | membrane potential |
| PAO | polyamine oxidase |
| DPI | diphenyleneiodonium (NOX inhibitor) |
References
- Bell, E.; Takeda, S.; Dolan, L. Reactive oxygen species in growth and development. In Reactive Oxygen Species in Plant Signaling. Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2009; pp. 43–53. [Google Scholar]
- Kwak, J.M.; Mori, I.C.; Pei, Z.-M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, S.M.B.; Brownlee, C.; Bothwell, J.H.F. A tip-high, Ca2+-interdependent, reactive oxygen species gradient is associated with polarized growth in Fucus serratus zygotes. Planta 2008, 227, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; Harper, J.F.; Palanivelu, R. A Fruitful Journey: Pollen Tube Navigation from Germination to Fertilization. Annu. Rev. Plant Biol. 2019, 70, 809–837. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.L.; Moreira, D.; Ferreira, M.J.; Pereira, A.M.; Coimbra, S. Insights into secrets along the pollen tube pathway in need to be discovered. J. Exp. Bot. 2019, 70, 2979–2992. [Google Scholar] [CrossRef]
- Zhang, M.J.; Zhang, X.S.; Gao, X.-Q. ROS in the Male–Female Interactions During Pollination: Function and Regulation. Front. Plant Sci. 2020, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Cheung, A.Y.; Qu, L.J. Pollen tube integrity regulation in flowering plants: Insights from molecular assemblies on the pollen tube surface. New Phytol. 2019, 222, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Weisenseel, M.H.; Jaffe, L.F. The major growth current through lily pollen tubes enters as K+ and leaves as H+. Planta 1976, 7, 1–7. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Feijó, J.A.; Wudick, M.M. Calcium is life. J. Exp. Bot. 2018, 69, 4147–4150. [Google Scholar] [CrossRef]
- Taylor, L.P.; Hepler, P.K. Pollen germination and tube growth. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 461–491. [Google Scholar] [CrossRef] [PubMed]
- Speranza, A.; Crinelli, R.; Scoccianti, V.; Geitmann, A. Reactive oxygen species are involved in pollen tube initiation in kiwifruit. Plant Biol. 2012, 14, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, A.V.; Matveyeva, N.P.; Polesskaya, O.G.; Yermakov, I.P. Generation of reactive oxygen species during pollen grain germination. Russ. J. Dev. Biol. 2009, 40, 345–353. [Google Scholar] [CrossRef]
- Pasqualini, S.; Cresti, M.; Del Casino, C.; Faleri, C.; Frenguelli, G.; Tedeschini, E.; Ederli, L. Roles for NO and ROS signalling in pollen germination and pollen-tube elongation in Cupressus arizonica. Biol. Plant. 2015, 59, 735–744. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, C.; Fang, M.; Xu, H.; Cao, M. The Cell Death in CMS Plants. Int. J. Biosci. Biochem. Bioinforma. 2011, 1, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Nie, H.; Cheng, C.; Hua, J. Mitochondrial proteomic analysis reveals that proteins relate to oxidoreductase activity play a central role in pollen fertility in cotton. J. Proteom. 2020, 225, 103861. [Google Scholar] [CrossRef]
- Deng, M.-H.; Wen, J.-F.; Huo, J.-L.; Zhu, H.-S.; Dai, X.-Z.; Zhang, Z.-Q.; Zhou, H.; Zou, X.-X. Relationship of metabolism of reactive oxygen species with cytoplasmic male sterility in pepper (Capsicum annuum L.). Sci. Hortic. (Amsterdam) 2012, 134, 232–236. [Google Scholar] [CrossRef]
- Li, S.; Wan, C.; Kong, J.; Zhang, Z.; Li, Y.; Zhu, Y. Programmed cell death during microgenesis in a Honglian CMS line of rice is correlated with oxidative stress in mitochondria. Funct. Plant Biol. 2004, 31, 369–376. [Google Scholar] [CrossRef]
- Maksimov, N.; Evmenyeva, A.; Breygina, M.; Yermakov, I. The role of reactive oxygen species in pollen germination in Picea pungens (blue spruce). Plant Reprod. 2018, 18, 761–767. [Google Scholar] [CrossRef]
- Kaya, H.; Nakajima, R.; Iwano, M.; Kanaoka, M.M.; Kimura, S.; Takeda, S.; Kawarazaki, T.; Senzaki, E.; Hamamura, Y.; Higashiyama, T.; et al. Ca2+-activated reactive oxygen species production by Arabidopsis RbohH and RbohJ is essential for proper pollen tube tip growth. Plant Cell 2014, 26, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.; Iwano, M.; Takeda, S.; Kanaoka, M.M.; Kimura, S. Apoplastic ROS production upon pollination by RbohH and RbohJ in Arabidopsis. Plant Signal. Behav. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, A.; Matveyeva, N.; Yermakov, I. Reactive oxygen species are involved in regulation of pollen wall cytomechanics. Plant Biol. 2013, 16, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Q.; Liu, C.Z.; Li, D.D.; Zhao, T.T.; Li, F.; Jia, X.N.; Zhao, X.Y.; Zhang, X.S. The Arabidopsis KINβγ Subunit of the SnRK1 Complex Regulates Pollen Hydration on the Stigma by Mediating the Level of Reactive Oxygen Species in Pollen. PLoS Genet. 2016, 12, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breygina, M.A.; Matveeva, N.P.; Ermakov, I.P. The role of Cl¯ in pollen germination and tube growth. Russ. J. Dev. Biol. 2009, 40, 157–164. [Google Scholar] [CrossRef]
- Breygina, M.; Smirnova, A.; Matveeva, N.; Yermakov, I. The role of anion channels in pollen germination and tube growth. In Pollen: Structure, Types and Effects; Kaiser, B.J., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2010; pp. 1–19. ISBN 9781616686697. [Google Scholar]
- Matveeva, N.P.; Andreyuk, D.S.; Voitsekh, O.O.; Ermakov, I.P. Regulatory Changes in the Intracellular pH and Cl-- Efflux at Early Stages of Pollen Grain Germination in vitro. Russ. J. Plant Physiol. 2003, 50, 318–323. [Google Scholar] [CrossRef]
- Rodriguez-Rosales, M.P.; Roldan, M.; Belver, A.; Donaire, J.P. Correlation between in vitro germination capacity and proton extrusion in olive pollen. Plant Physiol. Biochem. 1989, 27, 723–728. [Google Scholar]
- Fricker, M.D.; White, N.S.; Obermeyer, G. pH gradients are not associated with tip growth in pollen tubes of Lilium longiflorum. J. Cell Sci. 1997, 110, 1729–1740. [Google Scholar]
- Certal, A.C.; Almeida, R.B.; Carvalho, L.M.; Wong, E.; Moreno, N.; Michard, E.; Carneiro, J.; Rodriguéz-Léon, J.; Wu, H.; Cheung, A.Y.; et al. Exclusion of a proton ATPase from the apical membrane is associated with cell polarity and tip growth in Nicotiana tabacum pollen tubes. Plant Cell 2008, 20, 614–634. [Google Scholar] [CrossRef] [Green Version]
- Mazina, S.E.; Matveyeva, N.P.; Yermakov, I.P. Determination of a functional pore in the pollen grain of tobacco. Tsitologiya 2002, 44, 33–39. [Google Scholar]
- Weisenseel, M.H.; Nuccitelli, R.; Jaffe, L.F. Large electrical currents traverse growing pollen tubes. J. Cell Biol. 1975, 66, 556–567. [Google Scholar] [CrossRef]
- Iwano, M.; Shiba, H.; Miwa, T.; Che, F.-S.; Takayama, S.; Nagai, T.; Miyawaki, A.; Isogai, A. Ca2+ dynamics in a pollen grain and papilla cell during pollination of Arabidopsis. Plant Physiol. 2004, 136, 3562–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafra, A.; Rejón, J.D.; Hiscock, S.J.; Alché, J.D.D. Patterns of ROS accumulation in the stigmas of angiosperms and visions into their multi-functionality in plant reproduction. Front. Plant Sci. 2016, 7, 1112–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnis, S.M.; Desikan, R.; Hancock, J.T.; Hiscock, S.J. Production of reactive oxygen species and reactive nitrogen species by angiosperm stigmas and pollen: Potential signalling crosstalk? New Phytol. 2006, 172, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Hiscock, S.J.; Bright, J.; McInnis, S.M.; Desikan, R.; Hancock, J.T. Signaling on the stigma. Potential new roles for ROS and NO in plant cell signaling. Plant Signal. Behav. 2007, 2, 23–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bredemeijer, G.M.M. The role of peroxidases in pistil-pollen interactions. Theor. Appl. Genet. 1984, 68, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Shivanna, K.R.; Rangaswamy, N.S. Pollen Biology: A Laboratory Manual; Springer Science & Business Media: Berlin, Germany, 2012; ISBN 3642773060. [Google Scholar]
- Žárský, V.; Říhová, L.; Tupý, J. Interference of pollen diffusable substances with peroxidase catalyzed reaction. Plant Sci. 1987, 52, 29–32. [Google Scholar] [CrossRef]
- Kärkönen, A.; Kuchitsu, K. Reactive oxygen species in cell wall metabolism and development in plants. Phytochemistry 2015, 112, 22–32. [Google Scholar] [CrossRef]
- Lindsay, S.E.; Fry, S.C. Redox and Wall-Restructuring. In The Expanding Cell; Verbelen, J.-P., Vissenberg, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 159–190. ISBN 978-3-540-39116-6. [Google Scholar]
- Bunzel, M. Chemistry and occurrence of hydroxycinnamate oligomers. Phytochem. Rev. 2010, 9, 47–64. [Google Scholar] [CrossRef]
- Breygina, M.; Maksimov, N.; Polevova, S.; Evmenyeva, A. Bipolar pollen germination in blue spruce (Picea pungens). Protoplasma 2019, 256, 941–949. [Google Scholar] [CrossRef]
- Çetinbaş-Genç, A.; Vardar, F. Effect of methyl jasmonate on in-vitro pollen germination and tube elongation of Pinus nigra. Protoplasma 2020, 257, 1655–1665. [Google Scholar] [CrossRef]
- Breygina, M.A.; Abramochkin, D.V.; Maksimov, N.M.; Yermakov, I.P. Hydrogen peroxide affects ion channels in lily pollen grain protoplasts. Plant Biol. 2016, 18, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Shang, Z.; Wu, J.; Jiang, X.; Moschou, P.N.; Sun, W.; Roubelakis-Angelakis, K.A.; Zhang, S. Spermidine oxidase-derived H2O2 regulates pollen plasma membrane hyperpolarization-activated Ca2+-permeable channels and pollen tube growth. Plant J. 2010, 63, 1042–1053. [Google Scholar] [CrossRef] [PubMed]
- Maksimov, N.M.; Breygina, M.A.; Ermakov, I.P. Regulation of ion transport across the pollen tube plasmalemma by hydrogen peroxide. Cell Tissue Biol. 2016, 10, 69–75. [Google Scholar] [CrossRef]
- Hafidh, S.; Fila, J.; Honys, D. Male gametophyte development and function in angiosperms: A general concept. Plant Reprod. 2016, 29, 31–51. [Google Scholar] [CrossRef]
- Scholz, P.; Anstatt, J.; Krawczyk, H.E.; Ischebeck, T. Signalling Pinpointed to the Tip: The Complex Regulatory Network That Allows Pollen Tube Growth. Plants 2020, 9, 1098. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.; Ju, Y.; Kessler, S.A. Reactive Oxygen Species as Mediators of Gametophyte Development and Double Fertilization in Flowering Plants. Front. Plant Sci. 2020, 11, 1199. [Google Scholar] [CrossRef]
- Lan, X.; Yang, J.; Abhinandan, K.; Nie, Y.; Li, X.; Li, Y.; Samuel, M.A. Flavonoids and ROS Play Opposing Roles in Mediating Pollination in Ornamental Kale (Brassica oleracea var. acephala). Mol. Plant 2017, 10, 1361–1364. [Google Scholar] [CrossRef] [Green Version]
- Iwano, M.; Entani, T.; Shiba, H.; Kakita, M.; Nagai, T.; Mizuno, H.; Miyawaki, A.; Shoji, T.; Kubo, K.; Isogai, A.; et al. Fine-tuning of the cytoplasmic Ca2+ concentration is essential for pollen tube growth. Plant Physiol. 2009, 150, 1322–1334. [Google Scholar] [CrossRef] [Green Version]
- Gutermuth, T.; Lassig, R.; Portes, M.; Maierhofer, T.; Romeis, T.; Borst, J.; Hedrich, R.; Feijó, J.A.; Konrad, K.R. Pollen tube growth regulation by free anions depends on the interaction between the anion channel SLAH3 and calcium-dependent protein kinases CPK2 and CPK20. Plant Cell 2013, 25, 4525–4543. [Google Scholar] [CrossRef] [Green Version]
- Breygina, M.; Smirnova, A.; Matveeva, N.; Yermakov, I. Membrane potential changes during pollen germination and tube growth. Cell Tissue Biol. 2010, 3, 573–582. [Google Scholar] [CrossRef]
- Podolyan, A.; Maksimov, N.; Breygina, M. Redox-regulation of ion homeostasis in growing lily pollen tubes. J. Plant Physiol. 2019, 243, 153050. [Google Scholar] [CrossRef]
- Hepler, P.K.; Winship, L.J. The pollen tube clear zone: Clues to the mechanism of polarized growth. J. Integr. Plant Biol. 2015, 57, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Feijó, J.A.; Sainhas, J.; Hackett, G.R.; Kunkel, J.G.; Hepler, P.K. Growing pollen tubes possess a constitutive alkaline band in the clear zone and a growth-dependent acidic tip. J. Cell Biol. 1999, 144, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michard, E.; Lima, P.T.; Borges, F.; Silva, A.C.; Portes, M.T.; Carvalho, J.E.; Gilliham, M.; Liu, L.-H.; Obermeyer, G.; Feijó, J. a Glutamate receptor-like genes form Ca2+ channels in pollen tubes and are regulated by pistil D-serine. Science 2011, 332, 434–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, D.D.; Lazzaro, M.D.; Owens, J.N. Growth and development of conifer pollen tubes. Sex. Plant Reprod. 2005, 18, 149–162. [Google Scholar] [CrossRef]
- Cai, G.; Parrotta, L.; Cresti, M. Organelle trafficking, the cytoskeleton, and pollen tube growth. J. Integr. Plant Biol. 2015, 57, 63–78. [Google Scholar] [CrossRef]
- Qian, D.; Xiang, Y. Actin cytoskeleton as actor in upstream and downstream of calcium signaling in plant cells. Int. J. Mol. Sci. 2019, 20, 1403. [Google Scholar] [CrossRef] [Green Version]
- Lovy-Wheeler, A.; Kunkel, J.G.; Allwood, E.G.; Hussey, P.J.; Hepler, P.K. Oscillatory increases in alkalinity anticipate growth and may regulate actin dynamics in pollen tubes of lily. Plant Cell 2006, 18, 2182–2193. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Wong, E.I.; Vidali, L.; Estavillo, A.; Hepler, P.K.; Wu, H.M.; Cheung, A.Y. The regulation of actin organization by actin-depolymerizing factor in elongating pollen tubes. Plant Cell 2002, 14, 2175–2190. [Google Scholar] [CrossRef] [Green Version]
- Muhlemann, J.K.; Younts, T.L.B.; Muday, G.K. Flavonols control pollen tube growth and integrity by regulating ROS homeostasis during high-temperature stress. Proc. Natl. Acad. Sci. USA 2018, 115, E11188–E11197. [Google Scholar] [CrossRef] [Green Version]
- Do, T.H.T.; Choi, H.; Palmgren, M.; Martinoia, E.; Hwang, J.U.; Lee, Y. Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes. Proc. Natl. Acad. Sci. USA 2019, 116, 12540–12549. [Google Scholar] [CrossRef] [Green Version]
- Aloisi, I.; Cai, G.; Tumiatti, V.; Minarini, A.; Del Duca, S. Del Natural polyamines and synthetic analogues modify the growth and the morphology of Pyrus communis pollen tubes affecting ROS levels and causing cell death. Plant Sci. 2015, 239, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, R.L.; Zhang, L.; Wang, Q.L.; Niehaus, K.; Baluška, F.; Šamaj, J.; Lin, J.X. Lipid microdomain polarization is required for NADPH oxidase-dependent ROS signaling in Picea meyeri pollen tube tip growth. Plant J. 2009, 60, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Potocký, M.; Jones, M.A.; Bezvoda, R.; Smirnoff, N.; Žárský, V. Reactive oxygen species produced by NADPH oxidase are involved in pollen tube growth. New Phytol. 2007, 174, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Lassig, R.; Gutermuth, T.; Bey, T.D.; Konrad, K.R.; Romeis, T. Pollen tube NAD(P)H oxidases act as a speed control to dampen growth rate oscillations during polarized cell growth. Plant J. 2014, 78, 94–106. [Google Scholar] [CrossRef]
- Aloisi, I.; Cai, G.; Serafini-Fracassini, D.; Del Duca, S. Polyamines in Pollen: From Microsporogenesis to Fertilization. Front. Plant Sci. 2016, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Kim, D.W.; Niitsu, M.; Maeda, S.; Watanabe, M.; Kamio, Y.; Berberich, T.; Kusano, T. Polyamine Oxidase 7 is a Terminal Catabolism-Type Enzyme in Oryza sativa and is Specifically Expressed in Anthers. Plant Cell Physiol. 2014, 55, 1110–1122. [Google Scholar] [CrossRef] [Green Version]
- Benkő, P.; Jee, S.; Kaszler, N.; Fehér, A.; Gémes, K. Polyamines treatment during pollen germination and pollen tube elongation in tobacco modulate reactive oxygen species and nitric oxide homeostasis. J. Plant Physiol. 2020, 244, 153085. [Google Scholar] [CrossRef]
- Pottosin, I.; Shabala, S. Polyamines control of cation transport across plant membranes: Implications for ion homeostasis and abiotic stress signaling. Front. Plant Sci. 2014, 5, 154. [Google Scholar] [CrossRef] [Green Version]
- Aloisi, I.; Cai, G.; Faleri, C.; Navazio, L.; Serafini-Fracassini, D.; Del Duca, S. Spermine Regulates Pollen Tube Growth by Modulating Ca2+-Dependent Actin Organization and Cell Wall Structure. Front. Plant Sci. 2017, 8, 1701. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, L.; McKenna, S.T.; Kunkel, J.G.; Hepler, P.K. NAD(P)H oscillates in pollen tubes and is correlated with tip growth. Plant Physiol. 2006, 142, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.L.; Wu, J.; Xu, G.H.; Gao, Y.B.; Chen, G.; Wu, J.Y.; Wu, H.Q.; Zhang, S.L. S-RNase disrupts tip-localized reactive oxygen species and induces nuclear DNA degradation in incompatible pollen tubes of Pyrus pyrifolia. J. Cell Sci. 2010, 123, 4301–4309. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.M.; Lopes, A.L.; Coimbra, S. Arabinogalactan proteins as interactors along the crosstalk between the pollen tube and the female tissues. Front. Plant Sci. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mizuta, Y.; Higashiyama, T. Chemical signaling for pollen tube guidance at a glance. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Liu, C.; Fu, R.; Zhang, M.; Li, H.; Shen, L.; Wei, Q.; Sun, X.; Xu, L.; Ni, B.; et al. LORELEI-LIKE GPI-ANCHORED PROTEINS 2/3 Regulate Pollen Tube Growth as Chaperones and Coreceptors for ANXUR/BUPS Receptor Kinases in Arabidopsis. Mol. Plant 2019, 12, 1612–1623. [Google Scholar] [CrossRef]
- Li, H.J.; Yang, W.C. Ligands Switch Model for Pollen-Tube Integrity and Burst. Trends Plant Sci. 2018, 23, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Kita, D.; Johnson, E.A.; Aggarwal, M.; Gates, L.; Wu, H.-M.; Cheung, A.Y. Reactive oxygen species mediate pollen tube rupture to release sperm for fertilization in Arabidopsis. Nat. Commun. 2014, 5, 3129. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Frietsch, S.; Kim, T.-H.; Dizon, M.B.; Schroeder, J.I. The peroxin loss-of-function mutation abstinence by mutual consent disrupts male-female gametophyte recognition. Curr. Biol. 2008, 18, 63–68. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breygina, M.; Klimenko, E. ROS and Ions in Cell Signaling during Sexual Plant Reproduction. Int. J. Mol. Sci. 2020, 21, 9476. https://doi.org/10.3390/ijms21249476
Breygina M, Klimenko E. ROS and Ions in Cell Signaling during Sexual Plant Reproduction. International Journal of Molecular Sciences. 2020; 21(24):9476. https://doi.org/10.3390/ijms21249476
Chicago/Turabian StyleBreygina, Maria, and Ekaterina Klimenko. 2020. "ROS and Ions in Cell Signaling during Sexual Plant Reproduction" International Journal of Molecular Sciences 21, no. 24: 9476. https://doi.org/10.3390/ijms21249476
APA StyleBreygina, M., & Klimenko, E. (2020). ROS and Ions in Cell Signaling during Sexual Plant Reproduction. International Journal of Molecular Sciences, 21(24), 9476. https://doi.org/10.3390/ijms21249476