Characterization of In Vitro and In Vivo Metabolism of Antazoline Using Liquid Chromatography-Tandem Mass Spectrometry

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
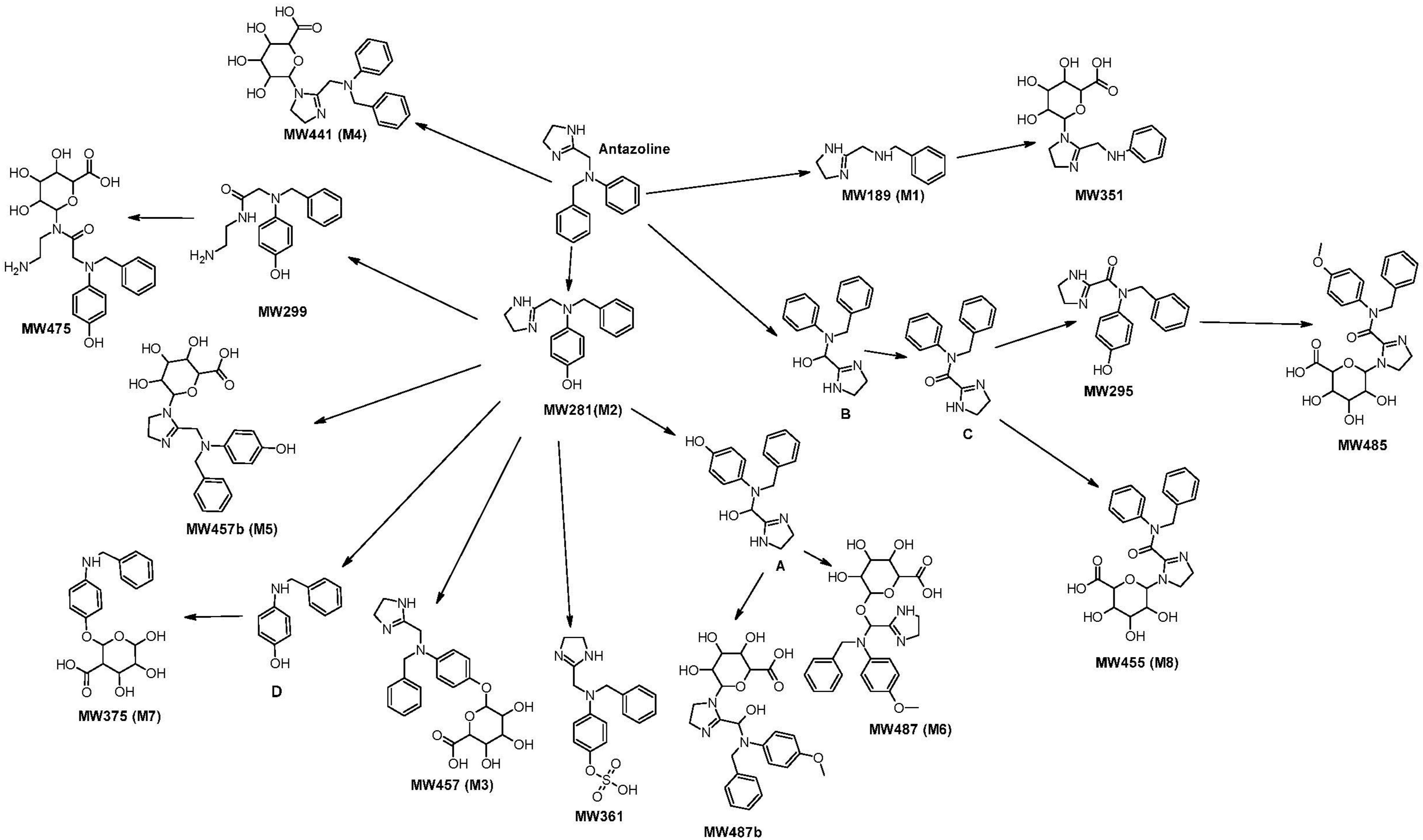
2.1. Determination of Metabolites in Plasma of Volunteers and Hepatocytes Cultures
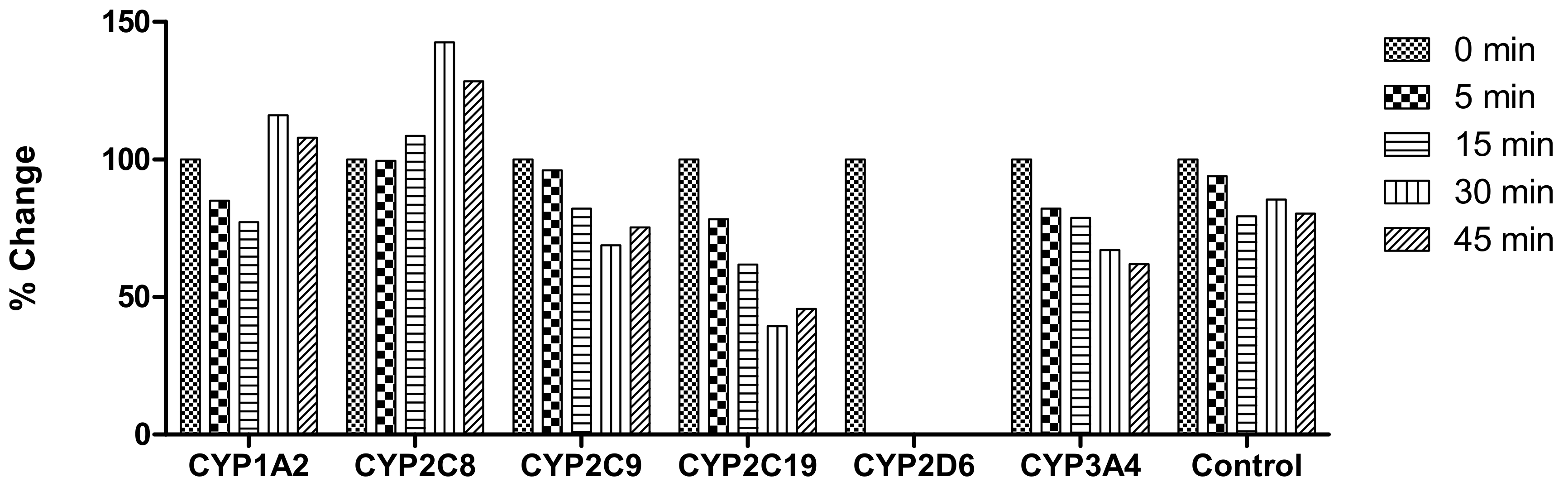
2.2. In Vitro Analysis Using Various CYP Isoforms
3. Materials and Methods
3.1. Sample Preparation
3.1.1. In Vivo Analysis
3.1.2. In Vitro Analysis
3.2. Instrumental Analysis
3.2.1. In Vivo Analysis
- The proposed formula corresponded to products of Phase I or Phase II metabolic biotransformations.
- The peak area of the compound increased at least five times and at least in three sampling timepoints (vs. time 0) with a gradual increase after drug administration in plasma of both volunteers.
- The difference between experimental and theoretical molecular weight was not higher than 5 ppm.
3.2.2. In Vitro Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Experimental Molecular Weight [Da] | RDBE | H/C | SFit [%] | Pattern Cov. [%] |
|---|---|---|---|---|---|
| MW189 | 189.1263 | 6 | 1.4 | 84 | 99.88 |
| MW281 | 281.1525 | 10 | 1.1 | 89 | 99.91 |
| MW295 | 295.1320 | 11 | 1 | 92 | 98.37 |
| MW299 | 299.1634 | 9 | 1.2 | 98 | 98.37 |
| MW351 | 351.1427 | 8 | 1.3 | 82 | 97.77 |
| MW361 | 361.1095 | 10 | 1.1 | 76 | 95.45 |
| MW375 | 375.1318 | 10 | 1.1 | 96 | 97.1 |
| MW441 | 441.1901 | 12 | 1.2 | 65 | 100 |
| MW455 | 455.1702 | 13 | 1.1 | 85 | 99.55 |
| MW457 | 457.1849 | 12 | 1.2 | 78 | 100 |
| MW457b | 457.1850 | 12 | 1.2 | 85 | 100 |
| MW475 | 475.1957 | 11 | 1.3 | 95 | 95.96 |
| MW485 | 485.1795 | 13 | 1.1 | 89 | 95.74 |
| MW487 | 487.1957 | 12 | 1.2 | 82 | 99.47 |
| MW487b | 487.1957 | 12 | 1.2 | 91 | 95.74 |






References
- Heinberg, C.J. A new synthetic anti-histaminic substance, antistine. Eye Ear. Nose Throat Mon. 1947, 26, 639–641. [Google Scholar] [PubMed]
- Maciag, A.; Farkowski, M.M.; Chwyczko, T.; Beckowski, M.; Syska, P.; Kowalik, I.; Pytkowski, M.; Wozniak, J.; Dabrowski, R.; Szwed, H. Efficacy and safety of antazoline in the rapid cardioversion of paroxysmal atrial fibrillation (the AnPAF Study). Europace 2017, 19, 1637–1642. [Google Scholar] [CrossRef] [PubMed]
- Farkowski, M.M.; Maciag, A.; Zurawska, M.; Pytkowski, M.; Kowalik, I.; Wozniak, J.; Sterlinski, M.; Szwed, H. Comparative effectiveness and safety of antazolinebased and propafenonebased strategies for pharmacological cardioversion of shortduration atrial fibrillation in the emergency department. Pol. Arch. Med. Wewn. 2016, 126, 381–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wybraniec, M.T.; Wrobel, W.; Wilkosz, K.; Wrona, K.; Bula, K.; Mizia-Stec, K. Pharmacological Cardioversion With Antazoline in Atrial Fibrillation: The Results of the CANT Study. J. Am. Heart Assoc. 2018, 7, e010153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowski, R.; Krynski, T.; Baran, J.; Futyma, P.; Stec, S.; Kulakowski, P. Antazoline for rapid termination of atrial fibrillation during ablation of accessory pathways. Cardiol. J. 2014, 21, 299–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsam, P.; Kozluk, E.; Peller, M.; Piatkowska, A.; Lodzinski, P.; Kiliszek, M.; Koltowski, L.; Grabowski, M.; Opolski, G. Antazoline for termination of atrial fibrillation during the procedure of pulmonary veins isolation. Adv. Med. Sci. 2015, 60, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Farkowski, M.M.; Maciag, A.; Zurawska, M.; Kowalik, I.; Szwed, H.; Pytkowski, M. Clinical effectiveness and safety of antazoline-based therapy in patients with stable coronary artery disease undergoing pharmacological cardioversion of short-duration atrial fibrillation in the emergency department. Cardiovasc. Ther. 2018, 36, e12469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowski, R.; Giebultowicz, J.; Baran, J.; Sikorska, A.; Gralak-Lachowska, D.; Soszynska, M.; Wroczynski, P.; Kulakowski, P. Antazoline-insights into drug-induced electrocardiographic and hemodynamic effects: Results of the ELEPHANT II substudy. Ann. Noninvasive Electrocardiol. 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Selander, S.; Cramer, K. Agranulocytosis after penicillamine and antazoline. Br. Med. J. 1965, 2, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhof, P.; Benussi, S.; Kotecha, D.; Ahlsson, A.; Atar, D.; Casadei, B.; Castella, M.; Diener, H.-C.; Heidbuchel, H.; Hendriks, J.; et al. 2016 ESC Guidelines for the management of atrial fibrillation developed in collaboration with EACTS. Eur. Heart J. 2016, 37, 2893–2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giebultowicz, J.; Piotrowski, R.; Baran, J.; Kulakowski, P.; Wroczynski, P. Application of a novel liquid chromatography/tandem mass spectrometry method for the determination of antazoline in human plasma: Result of ELEPHANT-I [ELEctrophysiological, pharmacokinetic and hemodynamic effects of PHenazolinum (ANTazoline mesylate)] human pharmacokinetic study. J. Pharm. Biomed. Anal. 2016, 123, 113–119. [Google Scholar] [CrossRef]
- Binkowski, B.J.; Makowski, M.; Kubinski, P.; Lubinski, A. Effect of Antazoline on Electrophysiological Properties of Atrial Muscle and Conduction System of the Heart. Cardiovasc. Drugs Ther. 2018, 32, 169–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkowski, M.M.; Maciag, A.; Kowalik, I.; Konka, M.; Szwed, H.; Pytkowski, M. Intravenous antazoline, a first-generation antihistaminic drug with antiarrhythmic properties, is a suitable agent for pharmacological cardioversion of atrial fibrillation induced during pulmonary vein isolation due to the lack of influence on atrio-venous conduction and high clinical effectiveness (AntaEP Study). Br. J. Clin. Pharmacol. 2019, 85, 1552–1558. [Google Scholar] [CrossRef]
- Frommeyer, G.; Sterneberg, M.; Dechering, D.G.; Kaese, S.; Bogeholz, N.; Pott, C.; Fehr, M.; Bogossian, H.; Milberg, P.; Eckardt, L. Effective suppression of atrial fibrillation by the antihistaminic agent antazoline: First experimental insights into a novel antiarrhythmic agent. Cardiovasc. Ther. 2017, 35. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, T.H.; Madden, J.C.; Naidoo, V.; Walker, C.H. Comparative metabolism as a key driver of wildlife species sensitivity to human and veterinary pharmaceuticals. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Drug Administration. Safety Testing of Drug Metabolites. Guidance for Industry; Food and Drug Administration: Silver Spring, MD, USA, 2020. [Google Scholar]
- European Medicines Agency. ICH Topic M3(R2) Non-Clinical Safety Studies for the Conduct of Human Clinical Trials and Marketing Authorisation for Pharmaceuticals; European Medicines Agency: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Ali, H.M.; Beckett, A.H. Identification of two in vitro metabolic products after liver microsomal incubation of antazoline. J. Chromatogr. 1981, 210, 350–355. [Google Scholar] [CrossRef]
- Yahata, M.; Ishii, Y.; Nakagawa, T.; Watanabe, T.; Bando, K. Species differences in metabolism of a new antiepileptic drug candidate, DSP-0565 [2-(2′-fluoro[1,1′-biphenyl]-2-yl)acetamide]. Biopharm. Drug Dispos. 2019, 40, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Ingelman-Sundberg, M. Genetic polymorphisms of cytochrome P 450 2D6 (CYP2D6): Clinical consequences, evolutionary aspects and functional diversity. Pharmacogenom. J. 2005, 5, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, J.; Pesic, V.; Lauschke, V.M. Frequencies of clinically important CYP2C19 and CYP2D6 alleles are graded across Europe. Eur. J. Hum. Genet. 2020, 28, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Product | Experimental Molecular Weight [Da] | Retention Time [Min] | Molecular Formula | ΔMass [ppm] | Differences with ANT | Detected in vitro 1 | MS/MS Ions |
|---|---|---|---|---|---|---|---|
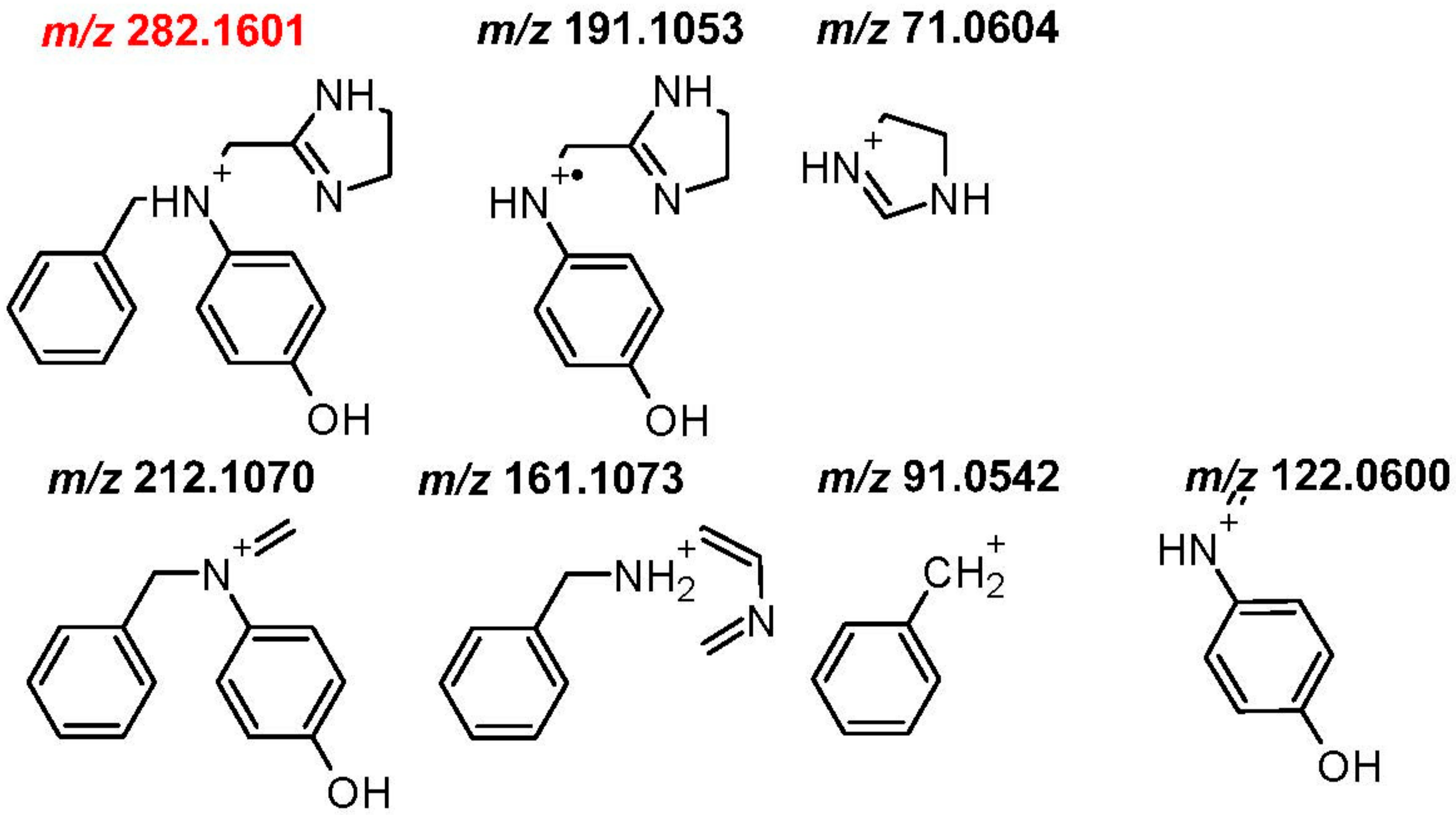
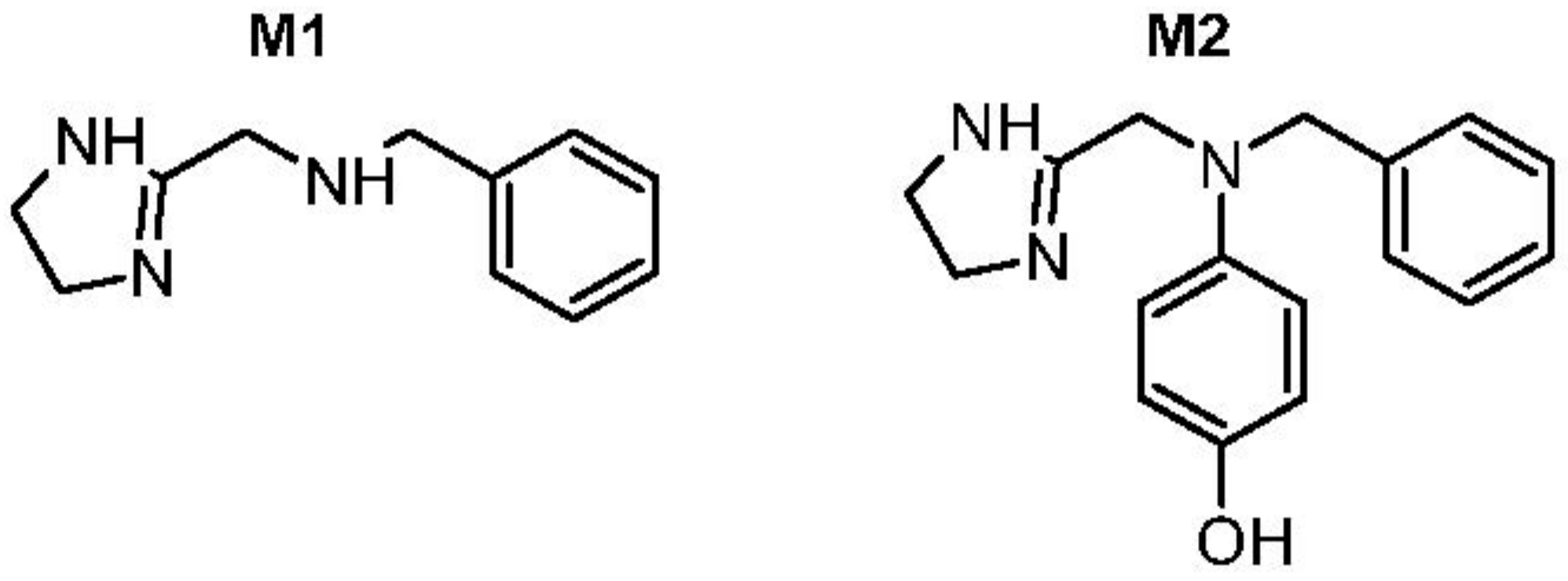
| M1 (MW189) | 189.1263 | 2.2 | C11 H15 N3 | −1.58 | −6C,4H | yes (3.6) | 120.0811, 91.0545, 71.0607 |
| M2 (MW281) | 281.1525 | 11.0 | C17 H19 N3 O | −1.10 | +O | yes (6.6) | 212.1075, 191.1055, 161.1074, 122.0603, 91.0545, 71.0607 |
| MW295 | 295.1320 | 10.4 | C17 H17 N3 O2 | −0.23 | −2H,+2O | yes (6.6) | 205.0856, 188.0835, 91.0545, 71.0601 |
| MW299 | 299.1634 | 11.0 | C17 H21 N3 O2 | 0.06 | +2H,2O | yes (6.7) | 212.1071, 149.0475, 122.0603, 91.0543, 72.0444 |
| MW351 | 351.1427 | 13.5 | C16 H21 N3 O6 | −0.96 | −C,+2H,6O | no | -2 |
| MW361 | 361.1095 | 11.2 | C17 H19 N3 O4 S | −0.19 | +S,4O | yes (5.9) | -2 |
| M7 (MW375) | 375.1318 | 6.4 | C19 H21 N O7 | 0.13 | +2C,2H,7O,−2N | yes (5.6) | 358.1306, 224.1069, 200.1074, 122.0603, 109.0525, 91.0544 |
| M4 (MW441) | 441.1901 | 12.1 | C23 H27 N3 O6 | 0.45 | +6C,8H,6O | yes (7.1) | 266.1654, 196.1125, 161.1073, 91.0545, 71.0606 |
| M8 (MW455) | 455.1702 | 8.7 | C23 H25 N3 O7 | −1.70 | +6C,6H,7O | no | 376.1396, 280.1448, 189.0900, 91.0544, 71.0127 |
| M3 (MW457) | 457.1849 | 8.2 | C23 H27 N3 O7 | 0.08 | +6C,8H,7O | yes (4.8) | 388.1389, 282.1600,212.1071, 122.0600,91.0543,71.0606 |
| M5 (MW457b) | 457.1850 | 9.7 | C23 H27 N3 O7 | 0.32 | +6C,8H,7O | yes (5.3) | 282.1607, 212.1075, 122.0600, 91.0545, 71.0606 |
| MW475 | 475.1957 | 9.1 | C23 H29 N3 O8 | 0.69 | +6C,10H,8O | yes (5.1) | 458.1936, 300.1702, 282.1605, 212.1072, 122.0602, 91.0544, 71.0607 |
| MW485 | 485.1795 | 10.9 | C24 H27 N3 O8 | −0.45 | +7C,8H,8O | no | 310.1551, 257.0769, 242.1178, 91.0544, 81.0449 |
| M6 (MW487) | 487.1957 | 8.9 | C24 H29 N3 O8 | 0.49 | +7C,10H,8O | yes (5.1) | 418.1503, 312.1708, 242.1177, 221.1161, 152.0708, 91.0544, 71.0606 |
| MW487b | 487.1957 | 9.3 | C24 H29 N3 O8 | 0.55 | +7C,10H,8O | yes (5.3) | 312.1709, 176.1187, 137.0600, 106.0655, 71.0605 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giebułtowicz, J.; Korytowska, N.; Piotrowski, R.; Kułakowski, P.; Latacz, G.; Szymańska, E.; Wiśniowska, B.; Polak, S. Characterization of In Vitro and In Vivo Metabolism of Antazoline Using Liquid Chromatography-Tandem Mass Spectrometry. Int. J. Mol. Sci. 2020, 21, 9693. https://doi.org/10.3390/ijms21249693
Giebułtowicz J, Korytowska N, Piotrowski R, Kułakowski P, Latacz G, Szymańska E, Wiśniowska B, Polak S. Characterization of In Vitro and In Vivo Metabolism of Antazoline Using Liquid Chromatography-Tandem Mass Spectrometry. International Journal of Molecular Sciences. 2020; 21(24):9693. https://doi.org/10.3390/ijms21249693
Chicago/Turabian StyleGiebułtowicz, Joanna, Natalia Korytowska, Roman Piotrowski, Piotr Kułakowski, Gniewomir Latacz, Ewa Szymańska, Barbara Wiśniowska, and Sebastian Polak. 2020. "Characterization of In Vitro and In Vivo Metabolism of Antazoline Using Liquid Chromatography-Tandem Mass Spectrometry" International Journal of Molecular Sciences 21, no. 24: 9693. https://doi.org/10.3390/ijms21249693
APA StyleGiebułtowicz, J., Korytowska, N., Piotrowski, R., Kułakowski, P., Latacz, G., Szymańska, E., Wiśniowska, B., & Polak, S. (2020). Characterization of In Vitro and In Vivo Metabolism of Antazoline Using Liquid Chromatography-Tandem Mass Spectrometry. International Journal of Molecular Sciences, 21(24), 9693. https://doi.org/10.3390/ijms21249693