1. Introduction
Bacterial infections represent one of the major problems for human health, causing different types of diseases. In the last century, although the discovery and use of antibiotics have improved the possibility to counteract harmful microorganisms, their poorly controlled and inappropriate application has led to an effective bacterial resistance against these molecules, based on various mechanisms of defense, in order to avoid antibiotic effects [
1]. In this context, hospital nosocomial infections constitute an ongoing serious danger, especially for immunocompromised individuals or those with critical illnesses, of which different pathogenic bacteria often represent the main primary cause due to their ability to prevent any potential drug effect. Despite the development of novel antibiotic therapies still representing an efficient method, this approach not only presents an elevated cost but also risks causing further pathogen resistance [
2]. Thus, in order to improve the research of innovative strategies, in particular in clinical systems, both the study of alternative antimicrobial molecules and the identification of new models appear necessary. In recent decades, the discovery of many antimicrobial peptides opened the door to the possibility to effectively design new therapeutic tools. However, although these molecules show both high validity and selectivity for different pathogens, their cytotoxicity against host cells often indicates a disadvantage of their possible use [
3].
In the investigation of potential solutions, it has been observed that many enzymes are efficiently active against different harmful agents, and their application has been widely examined. Among these enzymes, ribonucleases recently acquired a certain importance due to their ability to take part in the regulation of several biological processes, including immune response. In particular, enzymes belonging to the T2 ribonuclease family (T2 RNases), representing a widespread group of proteins present in almost all organisms and possessing a highly conserved RNAse-regulating housekeeping role, are also involved in several key cellular processes [
4]. Indeed, by showing a high pleiotropic nature, T2 RNases have apparently acquired specific functionalities during their evolution in different organisms [
5,
6].
Interestingly, the involvement of T2 RNases in immune processes has been already demonstrated in plants, in which several ribonucleases are implicated in the defensive mechanisms of seeds and branches. Indeed, these proteins are able to counteract external harmful agents, such as viruses or bacteria, independently from the catalytic activity [
4,
6]. In humans, overexpression of the RNASET2 enzyme regulates the immune response in mice carcinoma models, promoting a pro-inflammatory response, recruiting immunocompetent cells and inducing M1/M2 macrophage phenotype transition in the microenvironment surrounding the tumor mass, which in turns triggers a marked oncosuppressive activity [
7,
8]. Interestingly, the ability to chemoattract macrophages has also been observed in the medicinal leech
H. verbana, in which the T2 RNase enzyme
HvRNASET2 appears directly involved in the modulation of innate immunity [
9,
10].
As observed in vertebrate models, the injection in the leech body wall of LPS and LTA, the main antigens of Gram-negative and Gram-positive, respectively, triggered an increased expression of the endogenous
HvRNASET2 in both host type I granulocytes and macrophages, indicating a direct modulating role of the innate immune response during bacterial infections [
9,
10]. In
H. verbana, type I granulocytes represent one of the first cellular populations activated during bacterial challenges. Once stimulated, these cells degranulate the content of their cytoplasmatic granules in the extracellular environment, releasing different types of immune molecules [
11]. When released,
HvRNASET2 agglutinated Gram-positive bacteria
Staphylococcus aureus both in vivo and in vitro, also altering the cell integrity. Indeed, the close interaction with the microorganism cell walls was confirmed by its specific localization on the bacterial external surfaces [
10]. The
HvRNASET2 expression pattern during the earlier phases of leech inflammation, coupled with the simultaneous burst of macrophages recruitment, confirmed its crucial role in counteracting pathogens [
9]. In particular, the simultaneous chemoattractant role of
HvRNASET2 against macrophages and the formation of bacterial clusters seem to be critically involved not only in increasing the number of pathogen-targeting immune cells but also in improving the macrophage phagocytic activity itself, a key prerequisite for the elimination of pathogens and possible cellular debris [
10]. These results suggest that T2 RNases represent evolutionarily conserved key molecules implicated in the regulation of pathogen–host defense processes [
10,
12], whose role should be better investigated in the search for new solutions against different types of infections.
Based on this evidence and on the results previously achieved with S. aureus, in the present study, we assessed the HvRNASET2 effects on the Gram-negative bacteria E. coli. 3D structure analysis has been predicted to further confirm the high level of conservation existing between T2 RNase family members. Furthermore, HvRNASET2 bioinformatics characterization was crucial not only for demonstrating the preservation of conserved peptide motifs, but also to identify particular amino acid residues that might explain the obtained results. Subsequently, the agglutinating ability of the leech RNase T2 enzyme was evaluated both in vitro and in vivo, revealing that it indifferently recognizes any type of bacterium, affecting bacterial viability and promoting macrophages phagocytosis.
3. Discussion
The emergence of novel antimicrobial agents to counteract possible harmful agents is derived from the necessity to adopt alternative and innovative therapeutic solutions. Indeed, the capacity of many bacterial strains to acquire multi-drug resistance has made most common treatments ineffective, causing an increase in problems both for public health and the economic status of several national health programs, thus stimulating the search for innovative molecules to be used as alternative powerful tools to replace the expensive and poorly effective treatments already used [
17,
18].
In this context, it has long been known that many enzymes are involved in host defense against different pathogens, affecting microorganisms’ viability and modulating the immune response. Among them, it was recently demonstrated in the medicinal leech H. verbana that the HvRNASET2 enzyme plays a pivotal role in host defense, especially during bacterial infections. Indeed, injection in the leech body wall of both the bacterial antigens LPS and LTA induces a rapid increase in the endogenous expression of this enzyme [
9,
10]. Moreover, in vivo and in vitro experiments carried out on Gram-positive S. aureus cells revealed that HvRNASET2 agglutinates living cells and triggers the formation of several bacterial clusters. In addition, several blebs were clearly visible on cell membranes, indicating a local destabilization [
10].
Based on these data, in the present study, the antibacterial role of HvRNASET2 has been better evaluated by means of in silico, in vitro and in vivo analyses. In particular, we assessed its ability to affect Gram-negative bacteria, in particular the E. coli BL21(DE3) strain, revealing how HvRNASET2 actually interacts independently with different types of microorganisms.
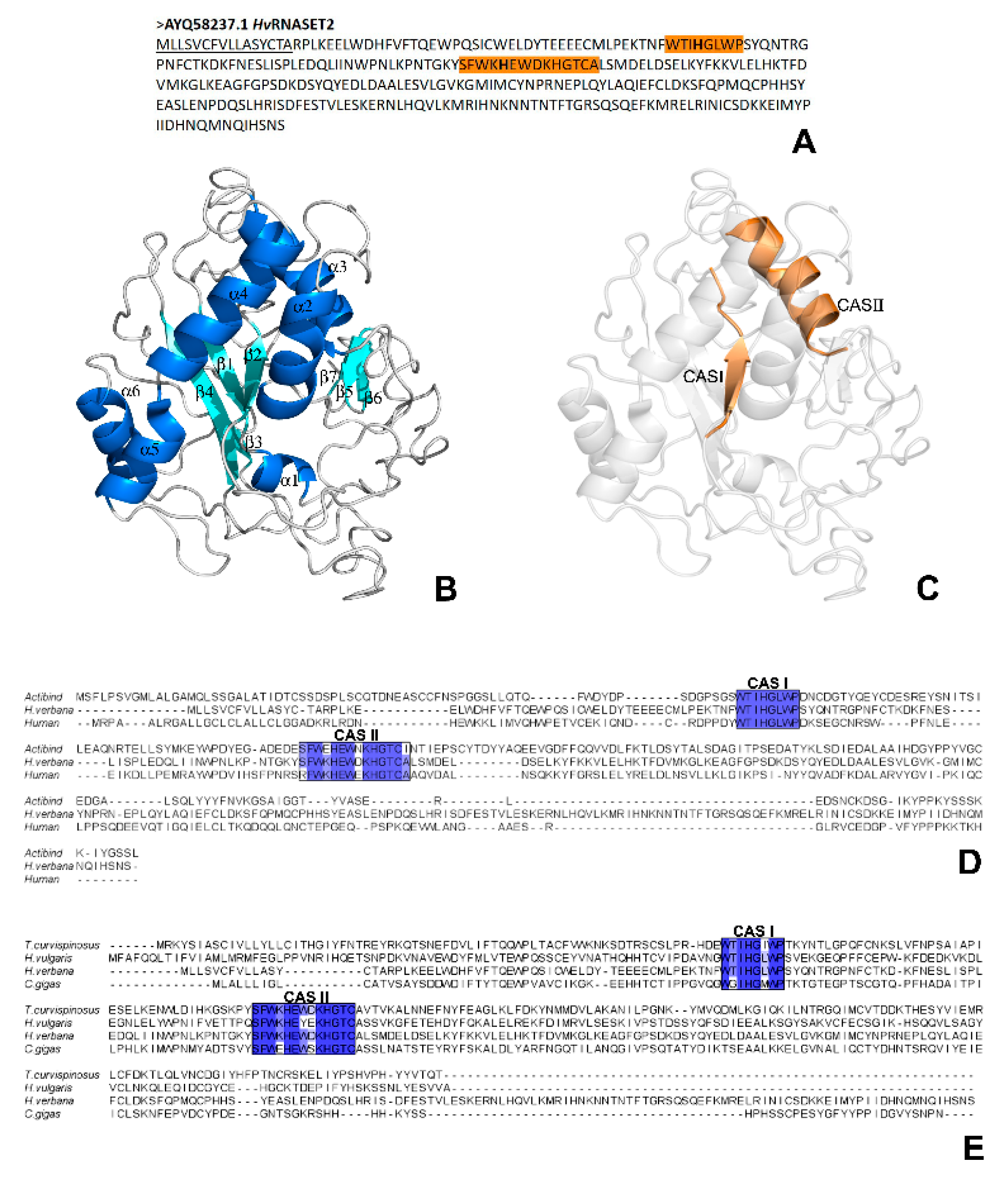
A first bioinformatic characterization revealed that the
H. verbana T2 ribonuclease presents a high level of conservation with other members of the T2 RNase family and, by analyzing both the primary amino acid sequence and the three-dimensional secondary structure, many similarities were clearly detected. Indeed, the presence of the two conserved Rh/T2/S ribonuclease-specific active sites, CAS I and CAS II, was confirmed by the PROSITE server. These fundamental domains, respectively localized in positions Trp60-Pro67 and Ser110-Ala123 in the HvRNASET2 protein, are normally involved in the enzymatic housekeeping role of T2 RNases, which consists in the control of RNA processing or degradation. In particular, the three-dimensional conformation predicted with I-TASSER not only confirmed a typical α + β structural organization, with a central core of β-sheets surrounded by several α-helices, but also identified the precise localization of the CAS I and CAS II domains, respectively, on the central β2-strand and on the parallel α3-helix, as for other RNase T2 enzymes [
19,
20,
21]. The functional relevance of these putative catalytic sites was further confirmed by the presence of the two His residues (His61 and His117), which were critically involved in the catalytic activity in both active sites [
22,
23]. This evidence was also validated by multiple sequence alignments to compare the leech HvRNASET2 sequence with those from different T2 RNases. In particular, the alignment with both the human RNASET2 and A. niger ACTIBIND or with several invertebrate T2 members highlighted the current similarities.
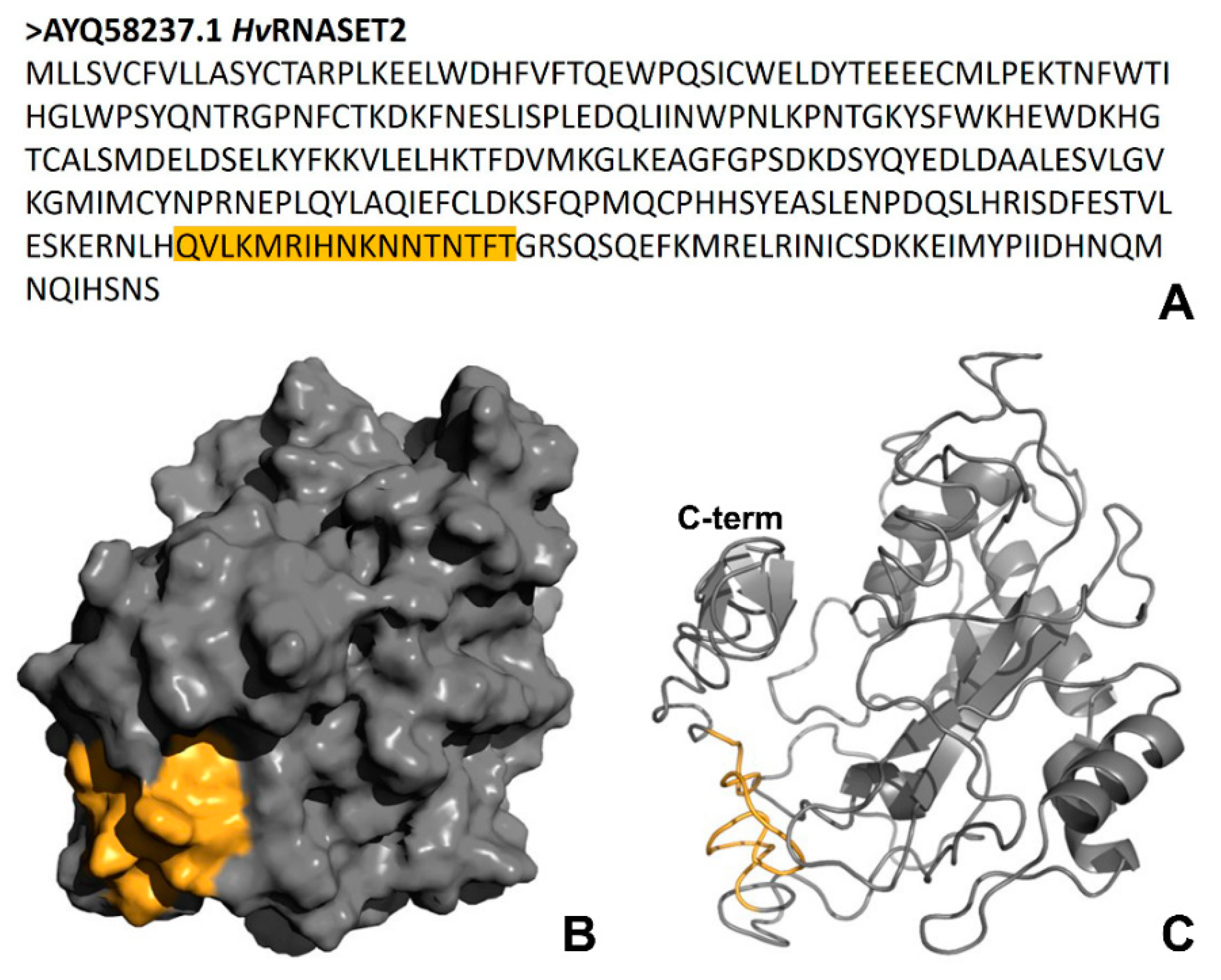
Of note, bioinformatic analyses carried out with the AMPA software, which predicts the presence of putative antimicrobial peptides in a protein of interest [
13], showed the existence of an amino acid motif with potential antibacterial properties in the
HvRNASET2 C-terminal region. By evaluating specific parameters, it was observed that this domain possesses both an elevated Boman index (3.04 kcal/mol) and a positive total net charge (+3.25). Interestingly, the Boman index indicates the probability for a peptide to directly bind to biological membranes, while the presence of a positive charge suggests a mechanism of interaction similar to that observed for many AMPs. Indeed, antimicrobial peptides are cationic and amphipathic molecules, which interact with the negative charges of the external bacterial walls in order to trigger membrane disruption and the activation of intracellular processes that lead to the death of microorganisms [
18,
24,
25]. Both electrostatic and hydrophobic interactions induce bacterial membrane destabilization, lead to the disruption of the bilayer integrity or affect the membrane fluidity [
26,
27]. Moreover, the peptide could enter the cytoplasmic environment to interact with other potential bacterial targets. Although different models have been identified for cationic AMPs (barrel-stave, carpet, toroidal-tole and translocation) [
25], the
HvRNASET2 mechanism of action could appear very similar. However, further experiments will be performed to better investigate the
HvRNASET2 mechanism of action and its ability to affect membranes. Since both Gram-positive and Gram-negative bacteria share the same cytoplasmic membrane,
HvRNASET2 could act against microorganisms regardless of their cell wall organization. Furthermore, Gram-negative bacteria display an additional potential target that is the outer membrane. Of note, other ribonucleases have been classified as antimicrobial proteins or enzymes, constituting a key first line of host defense against invading pathogens. The human RNase 3 (ECP), specifically secreted from the secondary granules of eosinophils, is released during inflammatory processes. ECP shows an antibacterial role independent of its catalytic activity, mediated by a cationic peptide in the N-terminal region that is able to induce both Gram-positive and Gram-negative bacteria agglutination and to inhibit cell growth [
28].
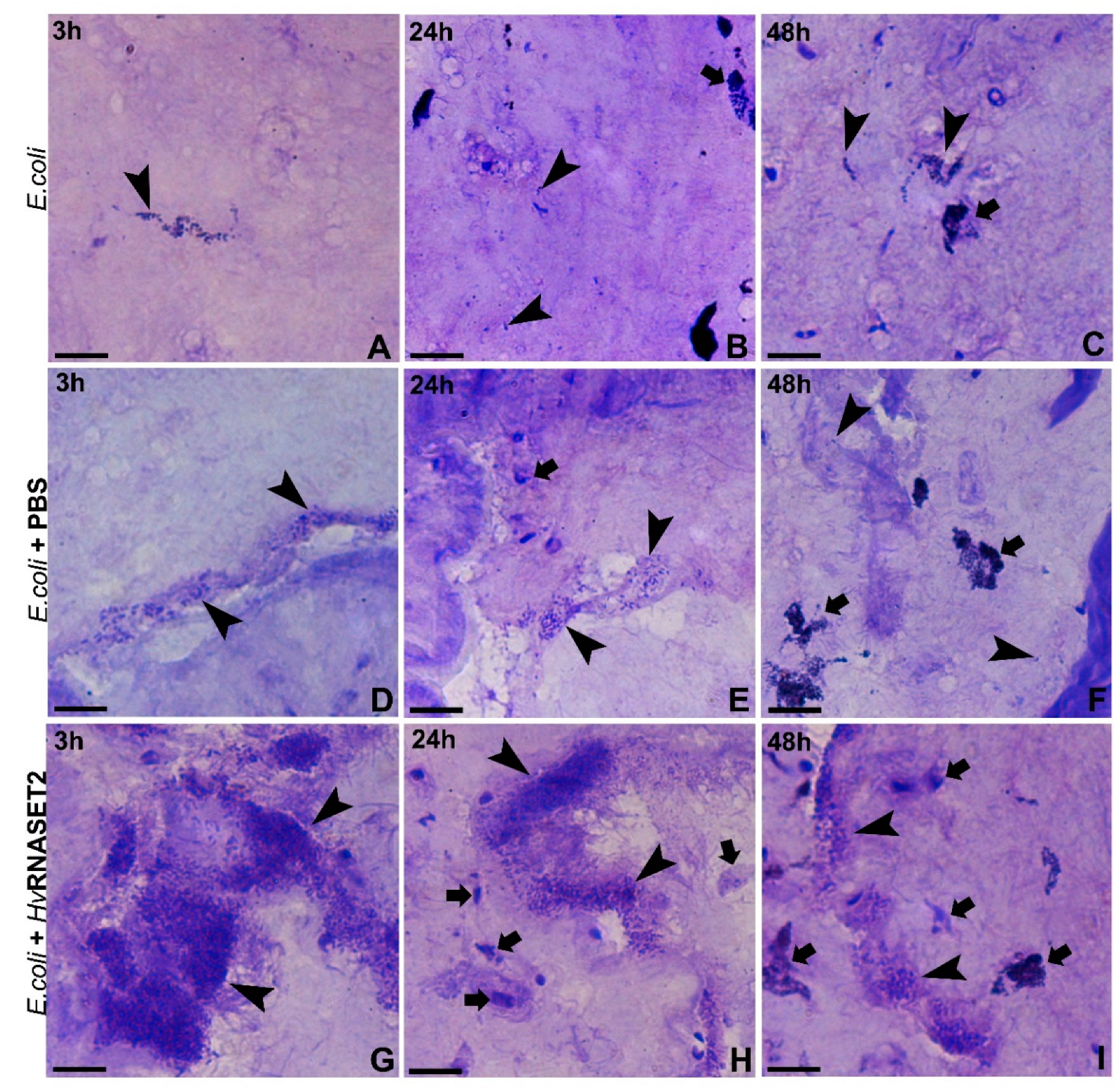
Here, by means of optical and electron microscopy, a marked agglutinating effect on bacterial cells has been demonstrated for
HvRNASET2 as well. As previously reported in
S. aureus [
10], morphological studies performed on
E. coli show that these microorganisms displayed a severe agglutination following
HvRNASET2 incubation compared to control samples. In particular, the aggregation process was clearly detectable by both TEM and SEM analyses, whereby not only were numerous cell clumps observed, but microbial membranes appeared significantl damaged, with the cytoplasmatic content released in the surrounding environment. Particularly after 24 and 48 h from
HvRNASET2 incubation, the leech recombinant enzyme destabilized outer membranes and affected their integrity. These effects are likely due to direct interaction with bacterial components, as confirmed by both immunogold at TEM and LPS binding assays. Of note, the presence of gold particles on the cell external surfaces at TEM indicates a direct interaction of HvRNASET2 with the bacterial cell wall. In parallel, the increase in the Body-P cadaverine fluorescent signal suggests the direct binding of leech T2 enzyme to LPS. It is tempting to speculate that such interaction could be mediated by the observed C-terminal peptide within
HvRNASET2, which might confer a bactericidal role to the enzyme. The administration of
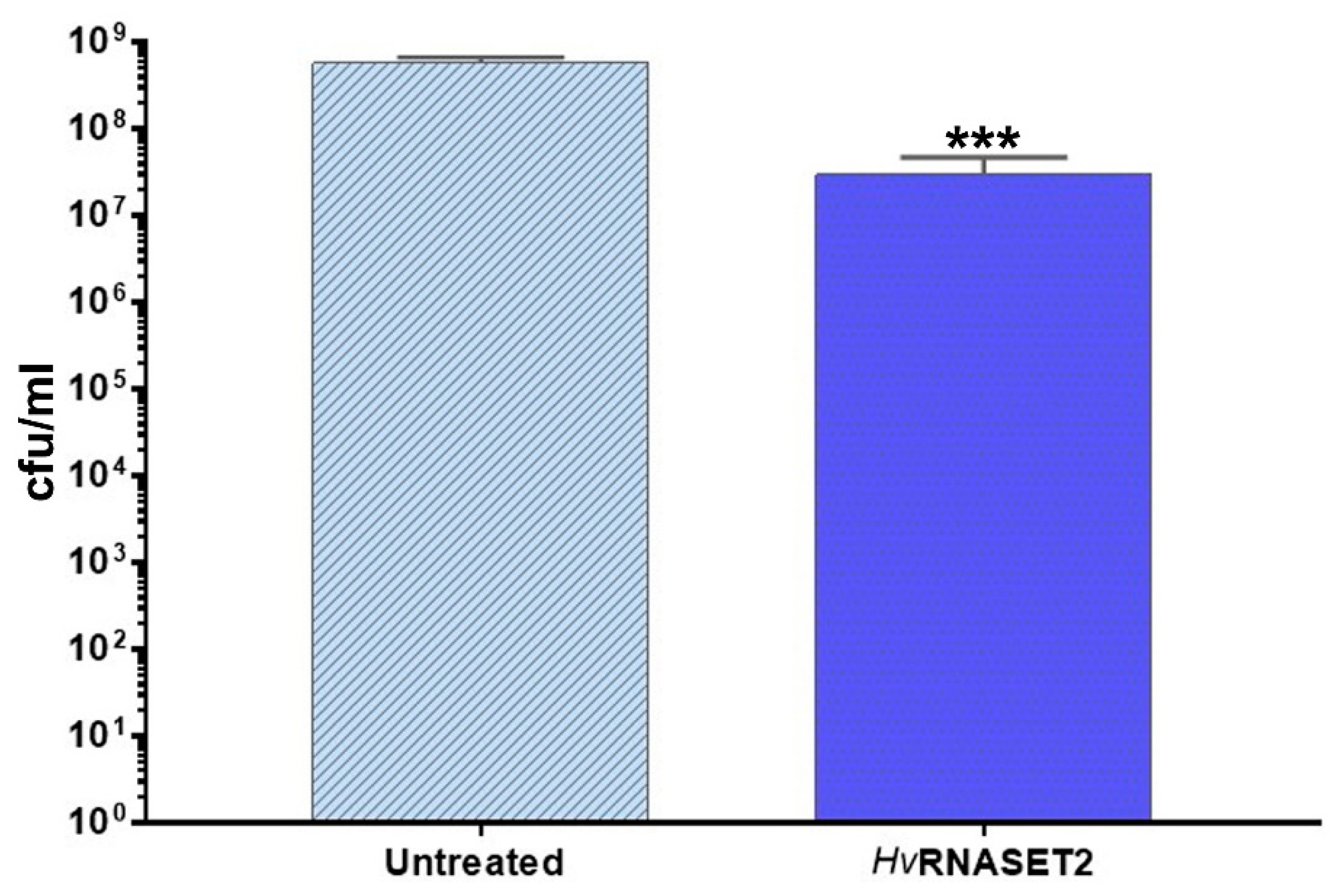
HvRNASET2 to
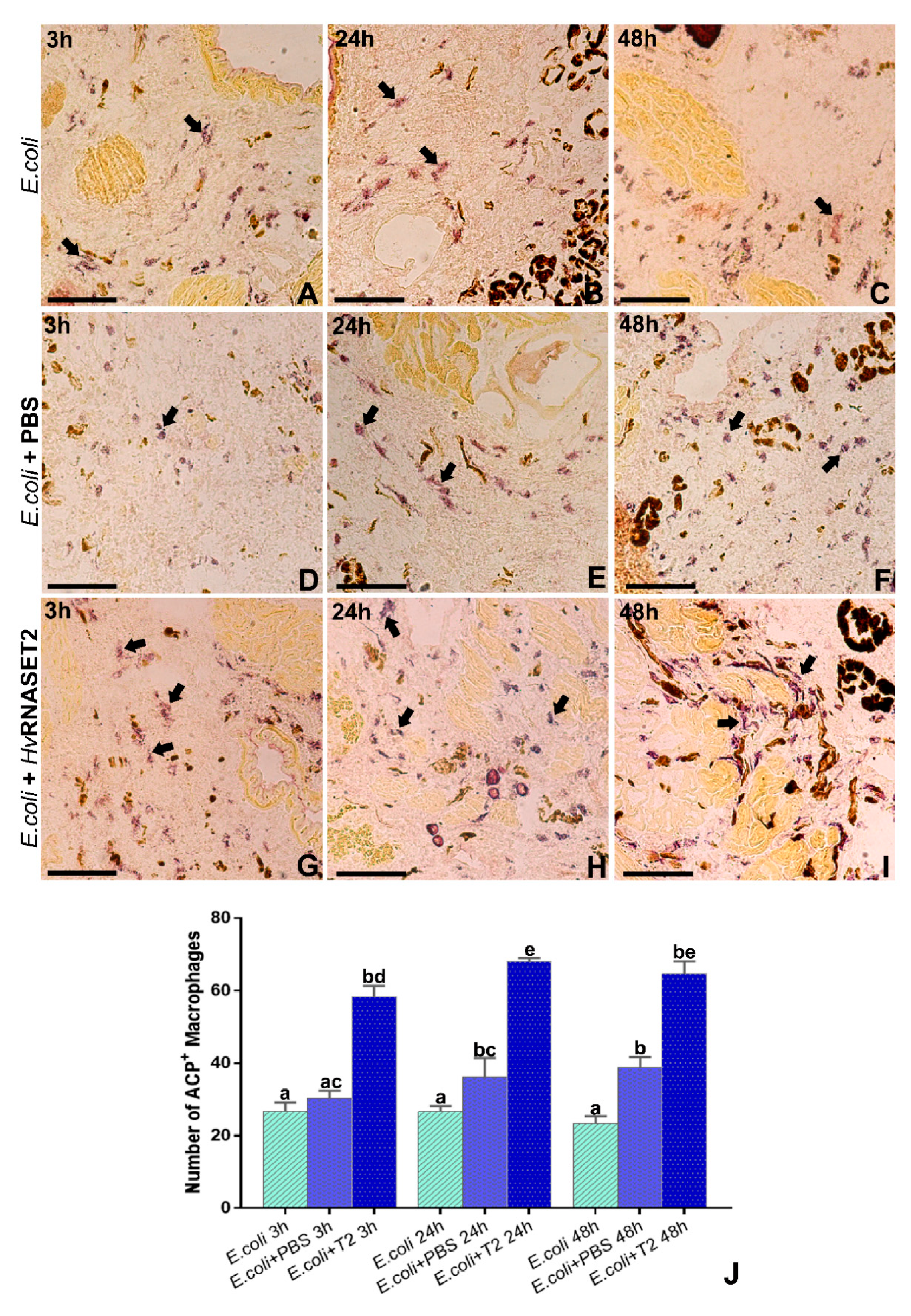
E. coli impaired in a significant manner the detection of viable cells. The observed decrease could be ascribable to a bactericidal effect and/or to an agglutination effect, as already observed in vivo experiments. Indeed, in silico analysis highlighted a putative antimicrobial stretch in RNASET2 that could affect cellular integrity. Moreover, an agglutination effect that can not be ruled out in the in vitro setup could underestimate the cellular concentration. However, regardless of the mechanism(s) involved, a statistically significant decrease of viable cells was observed upon HvRNASET2 treatment.
Finally, in vivo experiments, in which bacterial cells were directly injected in the leech body wall, pointed out cell agglutination as a prerequisite for enhancing the host’s innate immune response. As already hypothesized [
9], HvRNASET2 is specifically expressed and secreted by leech granulocytes during bacterial infections, playing a double role of recruiting host macrophages and at the same time interacting with bacterial pathogens. Indeed, the adhesion induced by rHvRNASET2 on
E. coli living cells promotes the activation of phagocytic macrophages, as observed with ACP assay. Interestingly, this specific mechanism is similar to that observed in several C-type lectins detected in shrimps of the genus Penaeus. Indeed, the PmLec lectin of Penaeus mondon not only agglutinates
E. coli but also binds to LPS in order to opsonize microbes and promote phagocytic events [
29]. Moreover, in the larvae of the insect Manduca sexta, immunolectin-2 shows a high affinity for LPS and triggers the formation of bacterial cell aggregates, also inducing phenol oxidase defensive reaction [
30]. Several cases of invertebrate immune molecules are reported in the literature that share the same effects, suggesting a common defensive mechanism being maintained throughout evolution in different organisms.
The discovery and the analysis of novel antimicrobial compounds will increase our knowledge of intrinsic host defenses and immune system. Identifying new alternative approaches to the expensive and poorly effective ones currently used is considered a key goal to improve human health. Exploiting the key features of the antimicrobial peptides such as efficiency and selectivity, broad range of targets, potentially low toxicity and accumulation in tissues, pharmaceutical industries aim to develop them as commercially available drugs, and several appropriate clinical trials are currently being conducted. Starting from our previous assumptions and experimental evidence [
9,
10,
12], the present research aimed to shed light on leech
HvRNASET2, which displays several features that make it a potentially valuable tool to develop novel antibacterial molecules, also due to its wide-range ability to act on different types of microbial pathogens.
4. Materials and Methods
4.1. Bioinformatic Prediction of the Leech HvRNASET2 Characteristics
4.2. Three-Dimensional Modelling of HvRNASET2
The three-dimensional structure of
HvRNASET2 was predicted with the I-TASSER (Iterative Threading ASSEmbly Refinement) server (
https://zhanglab.ccmb.med.umich.edu/I-TASSER/) [
33] based on the sequence homology of different structural templates present in the Protein Data Bank (PBD) library (
https://www.rcsb.org/) [
34]. The resulting PDB file was visualized and analyzed with PyMOL software (
https://pymol.org/2/) in order to characterize the secondary structure of the enzyme.
4.3. HvRNASET2 Alignment and Conservation Analysis
Possible amino acid similarities and the relative conserved domains related to the T2 ribonuclease family were determined by the structural alignment between
HvRNASET2 and other members of the T2 RNase family by using Clustal Omega (
https://www.ebi.ac.uk/Tools/msa/clustalo/) [
35]. Two different multiple alignments were performed with the following T2 RNase enzymes: human RNASET2 (AIC50165.1);
Aspergillus niger ACTIBIND (AAZ22530.1);
Hydra vulgaris (XP_002164769.1);
Crassostrea gigas (XP_011413920.2);
Temnothorax curvispinosus (XP_024871065.1). The graphical output showed in the figure was viewed with the Jalview program (
http://www.jalview.org/) [
36] and manually created.
4.4. Antimicrobial Peptide Identification
4.5. Light and Transmission Electron Microscopy
E. coli bacterial culture was grown at 37 °C overnight until reaching an OD600 of 0.6. Bacterial cells were then resuspended with the recombinant rHvRNASET2 (10 µM) for 3, 24 and 48 h at 20 °C in order to conduct in vitro experiments. As control, microorganisms were treated with PBS solution (PBS: 138 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4 and 1.5 mM KH2PO4, pH 7.4) at the same times. After treatments, bacteria were centrifuged for 10 min at 13.000 rpm and fixed for 2 h in 0.1 M cacodylate buffer at pH 7.4, containing 2% glutaraldehyde. Several washings were performed in the same buffer, and then cells were postfixed for 1 h with 1% osmium tetroxide in cacodylate buffer, pH 7.4. Subsequently, samples were embedded in an Epon-Araldite 812 mixture (Sigma-Aldrich, Milan, Italy) after serial ethanol dehydration (70%, 90%, 100%). Sections were obtained with a Reichert Ultracut S ultratome (Leica, Wien, Austria). Semi-thin sections (0.7 μm) were colored by conventional methods, crystal violet and basic fuchsin and observed under a light microscope Nikon Eclipse Ni (Nikon, Tokyo, Japan). Data were recorded with a DS-5M-L1 digital camera system (Nikon). Ultrathin sections (80 nm) were collected on copper grids (300 mesh, Sigma-Aldrich, Milan, Italy), counterstained by uranyl acetate and lead citrate, and observed with a Jeol 1010 EX transmission electron microscope TEM (Jeol, Tokyo, Japan). Data were recorded with a MORADA digital camera system (Olympus, Tokyo, Japan).
4.6. Scanning Electron Microscopy
Reaching an OD600 of 0.6, E. coli bacteria were incubated as previously described. After treatments, cells were centrifuged for 10 min at 13,000 rpm, and bacterial pellets were fixed with Karnovsky fixative (2% paraformaldehyde and 2.5% glutaraldehyde in 0.1 M cacodylate Buffer, pH 7.2) for 30 min at 4 °C. Samples were washed in 0.1 M cacodylate buffer (pH 7.2) and post-fixed in a solution of 1% osmium tetroxide and potassium ferrocyanide for 1 h. After several washings in PBS (pH 7.2) and dehydration with an increasing scale of ethanol, 20 µL of bacterial pellet resuspended in 100% ethanol were dried onto glass slides and finally subjected to critical point drying with hexamethyldisilazane. Images were acquired using SEM-FEG XL-30 microscope (Philips, Eindhoven, The Netherlands).
4.7. Immunogold Staining at TEM
Bacterial samples were fixed for 2 h at 4 °C with 4% paraformaldehyde and 0.5% glutaraldehyde in PBS, dehydrated in ethanol series and embedded in an Epon-Araldite 812 mixture (Sigma-Aldrich) as above. Ultrathin sections (80 nm) were collected on gold grids (300 mesh, Sigma-Aldrich). After etching with NaOH 3% in absolute ethanol [
37], slides were incubated for 30 min in blocking solution containing PBS, 1% bovine serum album (BSA) and 0.1% Tween and then with the polyclonal primary antibody rabbit anti-human RNASET2 [
38] diluted at 1:20 in blocking solution. After several washings with PBS, the primary antibody was visualized by immunostaining, with the secondary goat anti-rabbit IgG(H+L)-gold conjugate antibody (GE Healthcare, Amersham, UK; particle size, 10 nm) diluted at 1:100 in blocking solution for 1 h. In control experiments, the primary antibody was omitted, and sections were treated with BSA containing PBS and incubated only with the secondary antibodies. Sections were counterstained with uranyl acetate in water and observed withTEM, and data were recorded with a digital camera system as previously described.
4.8. LPS Binding Assay
The binding with the Gram-negative antigen LPS was evaluated by a fluorescent displacement assay using the fluorescent probe Bodipy cadaverine (Thermo Fisher Scientific, Waltham, MA, USA). The dissociation constant was measured by adding small volumes of concentrated stock of both Polymixin B (Thermo Fisher Scientific, Waltham, MA, USA), used as positive control, and HvRNASET2 to a solution of LPS (10 µM/mL) and fluorescent Bodipy TR cadaverine (10 µM) in 150 mM sodium phosphate buffer, pH 7.5, at 25 °C. Fluorescent Bodipy TR cadaverine normally binds to LPS, and the addition of a compound with a higher affinity for the bacterial antigen induces the detachment between Bodipy TR cadaverine and LPS and the consequent increase of the fluorescent signal. The fluorescence measurements were carried out with a 1 mL cell in a Jasco FP-750 spectrofluorometer (Jasco, Cremella, Italy) using an excitation wavelength of 580 nm. The change in emission at 620 nm was plotted as a function of ligand concentration.
4.9. Antimicrobial In Vitro Assay
E. coli BL21(DE3) was grown in LB broth at 37 °C under agitation. Upon overnight growth, cultures were diluted in fresh medium to reach a cellular concentration of ~105 cfu/mL. A sample was treated with rHvRNASET2 at a final concentration of 10 µM, and an untreated sample was included as control. Upon 24 h incubation, bacterial viability was checked by plate count technique and expressed as colony-forming units per mL (CFU/mL). A volume (0.01 mL) of undiluted or serially diluted samples was plated onto LB agar plates and incubated for 24 h at 37 °C. Experiments were independently repeated at least three times.
4.10. In Vivo Experiments: Animals and Treatments
Adult leeches (Hirudo verbana, Annelida, Hirudinea, from Ricarimpex, Eysines, France), measuring 10 cm were kept in lightly salted water (NaCl 1.5 g/mL), in aerated tanks and kept in an incubator at 20 °C. Before injection and/or dissection, leeches were anesthetized with a 10% ethanol solution, and all treatments were performed at the 80th superficial metamere from the oral sucker. Animals were randomly divided into separate experimental groups (three individuals for each time point) and submitted to various protocols and treatments, as described below:
Group 1: animals were injected with 50 μL of E. coli BL21(DE3) (108 CFU/mL), grown overnight in LB liquid soil (LB Broth, Sigma Aldrich, Milan, Italy). Bacteria, at an OD600 of 0.6, were injected after 3, 24 and 48 h of incubation, and leech tissues were collected 3 h after stimulation.
Group 2: animals were injected with 50 μL of E. coli BL21(DE3) (108 CFU/mL), grown overnight in LB liquid soil, resuspended in a 10 μM recombinant HvRNASET2 enzyme solution. Bacteria, at an OD600 of 0.6, were injected after 3, 24 and 48 h of incubation, and leech tissues were collected after 3 h from stimulation.
Group 3: animals were injected with 50 μL of E. coli BL21(DE3) (108 CFU/mL), grown overnight in LB liquid soil, resuspended in sterilized phosphate buffer saline (PBS). Bacteria, at an OD600 of 0.6, were injected after 3, 24 and 48 h of incubation, and leech tissues were collected after 3 h of stimulation.
4.11. Light Microscopy and Acid Phosphatase Assay (ACP)
Leech tissue samples, dissected from differently treated leech body walls, were embedded in Polyfreeze tissue freezing medium (OCT, Polysciences, Eppelheim, Germany) and immediately frozen in liquid nitrogen. Cryosections (7 µm) were obtained with a cryotome (Leica CM1850, Wetzlar, Germany), collected on gelatinous slides and counterstained with crystal violet and basic fuchsin for morphological analysis. All samples were mounted with Cityfluor (Cityfluor Ltd., London, UK) and examined with a Nikon Eclipse Ni (Nikon, Tokyo, Japan) light microscope. Data were recorded with a Nikon digital sight DS-SM (Nikon).
For ACP assay, cryosections were rehydrated with PBS and incubated with sodium acetate-acetic acid 0.1 M buffer for 5 min, followed by incubation with reaction mixture (sodium acetate-acetic acid 0.1 M buffer, 0.01% naphtol ASBI phosphate, 2% NN-dimethylformamide, 0.06% Fast RedViolet LB and MnCl2 0.5 nM) for 90 min at 37 °C. After several washings in PBS, slides were mounted with PBS/glycerol and observed with a Nikon Eclipse Ni (Nikon) as above.
4.12. Statistical Analyses
Statistical analyses were conducted on AMP and ACP assays. All the experiments were organized in triplicate, and data represented the mean values ± SD. In detail, for both experiments, either the total number of bacterial colonies or positive phagocytic macrophages were counted by hand. For the ACP assay, activated cells were assessed by analyzing five different slides (random fields of 45,000 μm2 for each slide) for each experiment, using the ImageJ software package. Subsequently, statistical analyses were executed using Statistica 7.0 software (StatSoft Inc., Tulsa, OK, USA), and differences were calculated respectively by unpaired t-test and one-way ANOVA followed by Tukey’s post-hoc test. p < 0.05 was considered statistically significant. The related graphs were created with GraphPad Prism 7 (GraphPad Software, La Jolla, CA, USA).


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






