Bisphenol A (BPA)-Induced Changes in the Number of Serotonin-Positive Cells in the Mucosal Layer of Porcine Small Intestine—the Preliminary Studies


Abstract
:1. Introduction
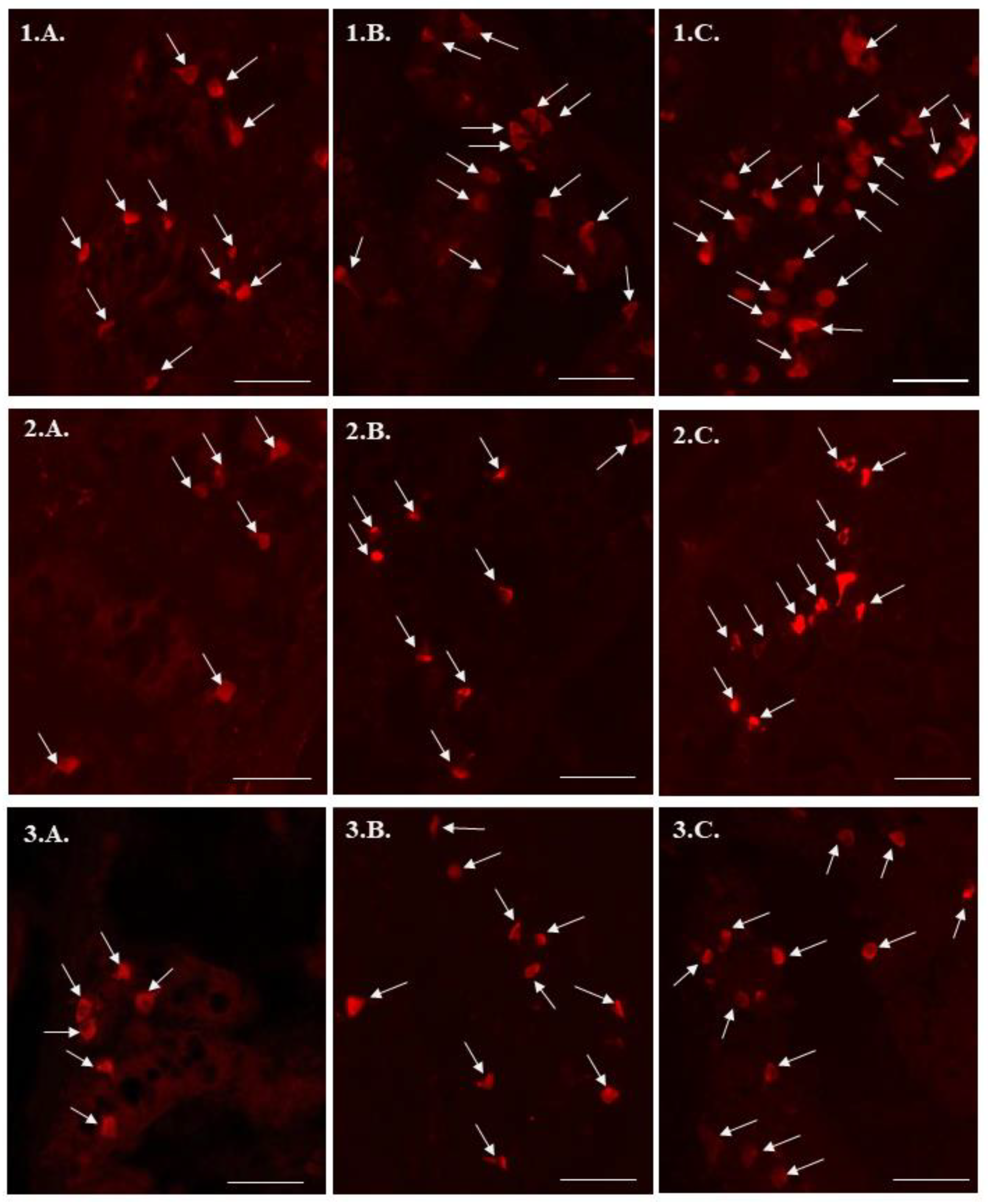
2. Results
3. Discussion
4. Materials and Methods
Funding
Conflicts of Interest
References
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Wazir, U.; Mokbel, K. Bisphenol A: A concise review of literature and a discussion of health and regulatory implications. In Vivo. 2019, 33, 1421–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.; Jiang, C.; Luo, J.; Cui, Y.; Qin, L.; Liu, J. Maternal exposure to bisphenol A may increase the risks of Parkinson’s disease through down-regulation of fetal IGF-1 expression. Med. Hypotheses. 2014, 82, 245–249. [Google Scholar] [CrossRef]
- Thongkorn, S.; Kanlayaprasit, S.; Jindatip, D.; Tencomnao, T.; Hu, V.W.; Sarachana, T. Sex differences in the effects of prenatal bisphenol A exposure on genes associated with autism spectrum disorder in the hippocampus. Sci. Rep. 2019, 9, 3038. [Google Scholar] [CrossRef] [PubMed]
- Ambreen, S.; Akhtar, T.; Hameed, N.; Ashfaq, I.; Sheikh, N. In vivo evaluation of histopathological alterations and trace metals estimation of the small intestine in bisphenol A-intoxicated rats. Can. J.Gastroenterol. Hepatol. 2019, 9, 9292316. [Google Scholar] [CrossRef]
- Zhao, Z.; Qu, W.; Wang, K.; Chen, S.; Zhang, L.; Wu, D.; Chen, Z. Bisphenol A inhibits mucin 2 secretion in intestinal goblet cells through mitochondrial dysfunction and oxidative stress. Biomed. Pharmacother. 2019, 111, 901–908. [Google Scholar] [CrossRef]
- Houdeau, E.; Moriez, R.; Leveque, M.; Salvador-Cartier, C.; Waget, A.; Leng, L.; Bueno, L.; Bucala, R.; Fioramonti, J. Sex steroid regulation of macrophage migration inhibitory factor in normal and inflamed colon in the female rat. Gastroenterology 2007, 132, 982–993. [Google Scholar] [CrossRef]
- Braniste, V.; Audebert, M.; Zalko, D.; Houdeau, E. Bisphenol A in the gut: Another break in the wall? In Multi-System Endocrine Disruption, 2nd ed.; Bourguignon, J.P., Jégou, B., Kerdelhué, B., Toppari, J., Christen, Y., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2011; pp. 127–144. ISBN 978-3-642-22774-5. [Google Scholar]
- Szymanska, K.; Makowska, K.; Gonkowski, S. The influence of high and low doses of bisphenol A (BPA) on the enteric nervous system of the porcine ileum. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Lesurtel, M.; Soll, C.; Graf, R.; Clavien, P.A. Role of serotonin in the hepato-gastrointestinal tract: An old molecule for new perspectives. Cell Mol. Life. Sci. 2008, 65, 940–952. [Google Scholar] [CrossRef]
- Banskota, S.; Ghia, J.E.; Khan, W.I. Serotonin in the gut: Blessing or a curse. Biochimie 2019, 161, 56–64. [Google Scholar] [CrossRef]
- Coates, M.D.; Tekin, I.; Vrana, K.E.; Mawe, G.M. Review article: The many potential roles of intestinal serotonin (5-hydroxytryptamine, 5-HT) signaling in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2017, 46, 569–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.; Miyazaki, A.; Saif, L.J. Immunohistochemical detection of the vomiting-inducing monoamine neurotransmitter serotonin and enterochromaffin cells in the intestines of conventional or gnotobiotic (Gn) pigs infected with porcine epidemic diarrhea virus (PEDV) and serum cytokine responses of Gn pigs to acute PEDV infection. Res. Vet. Sci. 2018, 119, 99–108. [Google Scholar] [PubMed]
- Atkinson, W.; Lockhart, S.; Whorwell, P.J.; Keevil, B.; Houghton, L.A. Altered 5-hydroxytryptamine signaling in patients with constipation- and diarrhea-predominant irritable bowel syndrome. Gastroenterology. 2006, 130, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Matsuzawa, D.; Ishii, D.; Tomizawa, H.; Sajiki, J.; Shimizu, E. Perinatal exposure to bisphenol A enhances contextual fear memory and affects the serotoninergic system in juvenile female mice. Horm. Behav. 2013, 63, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Ogi, H.; Fushiki, S.; Itoh, K. Prenatal and lactational bisphenol A exposure does not alter serotonergic neurons morphologically in the murine dorsal raphe nucleus. Brain. Dev. 2017, 39, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef]
- Almeida, S.; Raposo, A.; Almeida-Gonzales, M.; Carrascosa, C. Bisphenol A: Food exposure and impact on human health. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1503–1517. [Google Scholar] [CrossRef] [Green Version]
- Rytel, L. The influence of bisphenol A (BPA) on neuregulin 1-like immunoreactive nerve fibers in the wall of porcine uterus. Int. J. Mol. Sci. 2018, 19, E2962. [Google Scholar] [CrossRef]
- Qu, W.; Zhao, Z.; Chen, S.; Zhang, L.; Wu, D.; Chen, Z. Bisphenol A suppresses proliferation and induces apoptosis in colonic epithelial cells through mitochondrial and MAPK/AKT pathways. Life Sci. 2018, 208, 167–174. [Google Scholar] [CrossRef]
- Feng, L.; Chen, S.; Zhang, L.; Qu, W.; Chen, Z. Bisphenol A increases intestinal permeability through disrupting intestinal barrier function in mice. Environ. Pollut. 2019, 254, 112960. [Google Scholar] [CrossRef]
- Lu, X.; Li, M.; Wu, C.; Zhou, C.; Zhang, J.; Zhu, Q.; Shen, T. Bisphenol A promotes macrophage proinflammatory subtype polarization via upregulation of IRF5 expression in vitro. Toxicol. In Vitro 2019, 60, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Daly, D.M.; Adam, I.J.; Kitsanta, P.; Hill, C.J.; Wild, J.; Shorthouse, A.; Grundy, D.; Jiang, W. Interplay between mast cells, enterochromaffin cells, and sensory signaling in the aging human bowel. Neurogastroenterol. Motil. 2016, 28, 1465–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, K.; Tarafder, P.; Paul, G. Bisphenol A inhibits duodenal movement ex vivo of rat through nitric oxide-mediated soluble guanylyl cyclase and α-adrenergic signaling pathways. J. Appl. Toxicol. 2016, 36, 131–139. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Part of the Intestine | C Group | Exp I G | Exp II G |
|---|---|---|---|
| Duodenum | |||
| Animal 1 | 13.4 | 15.26 | 22.76 |
| Animal 2 | 13.96 | 16.34 | 21.72 |
| Animal 3 | 11.6 | 18.42 | 20.33 |
| Animal 4 | 12.26 | 16.94 | 17.34 |
| Animal 5 | 11.16 | 14.34 | 18.92 |
| Average ± SEM | 12.48 ± 0.53 | 16.26 ± 0.70 * | 20.21 ± 0.10 * |
| Jejunum | |||
| Animal 1 | 7.04 | 9.76 | 13.88 |
| Animal 2 | 6.96 | 10.34 | 12.96 |
| Animal 3 | 7.54 | 10.18 | 12.56 |
| Animal 4 | 7.64 | 10.52 | 13.64 |
| Animal 5 | 8.15 | 9.6 | 14.24 |
| Average ± SEM | 7.47 ± 0.22 | 10.08 ± 0.17 * | 13.46 ± 0.31 * |
| Ileum | |||
| Animal 1 | 8.42 | 13 | 13.92 |
| Animal 2 | 8.38 | 12.14 | 15.58 |
| Animal 3 | 9.64 | 11.16 | 13.06 |
| Animal 4 | 7.78 | 10.42 | 14.36 |
| Animal 5 | 8.72 | 13.84 | 16.52 |
| Average ± SEM | 8.59 ± 0.30 | 12.11 ± 0.61 * | 14.69 ± 0.61 * |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonkowski, S. Bisphenol A (BPA)-Induced Changes in the Number of Serotonin-Positive Cells in the Mucosal Layer of Porcine Small Intestine—the Preliminary Studies. Int. J. Mol. Sci. 2020, 21, 1079. https://doi.org/10.3390/ijms21031079
Gonkowski S. Bisphenol A (BPA)-Induced Changes in the Number of Serotonin-Positive Cells in the Mucosal Layer of Porcine Small Intestine—the Preliminary Studies. International Journal of Molecular Sciences. 2020; 21(3):1079. https://doi.org/10.3390/ijms21031079
Chicago/Turabian StyleGonkowski, Slawomir. 2020. "Bisphenol A (BPA)-Induced Changes in the Number of Serotonin-Positive Cells in the Mucosal Layer of Porcine Small Intestine—the Preliminary Studies" International Journal of Molecular Sciences 21, no. 3: 1079. https://doi.org/10.3390/ijms21031079
APA StyleGonkowski, S. (2020). Bisphenol A (BPA)-Induced Changes in the Number of Serotonin-Positive Cells in the Mucosal Layer of Porcine Small Intestine—the Preliminary Studies. International Journal of Molecular Sciences, 21(3), 1079. https://doi.org/10.3390/ijms21031079