Effects of Elevated Root-Zone CO2 on Root Morphology and Nitrogen Metabolism Revealed by Physiological and Transcriptome Analysis in Oriental Melon Seedling Roots

 , ,
, ,
Abstract
:1. Introduction
2. Results
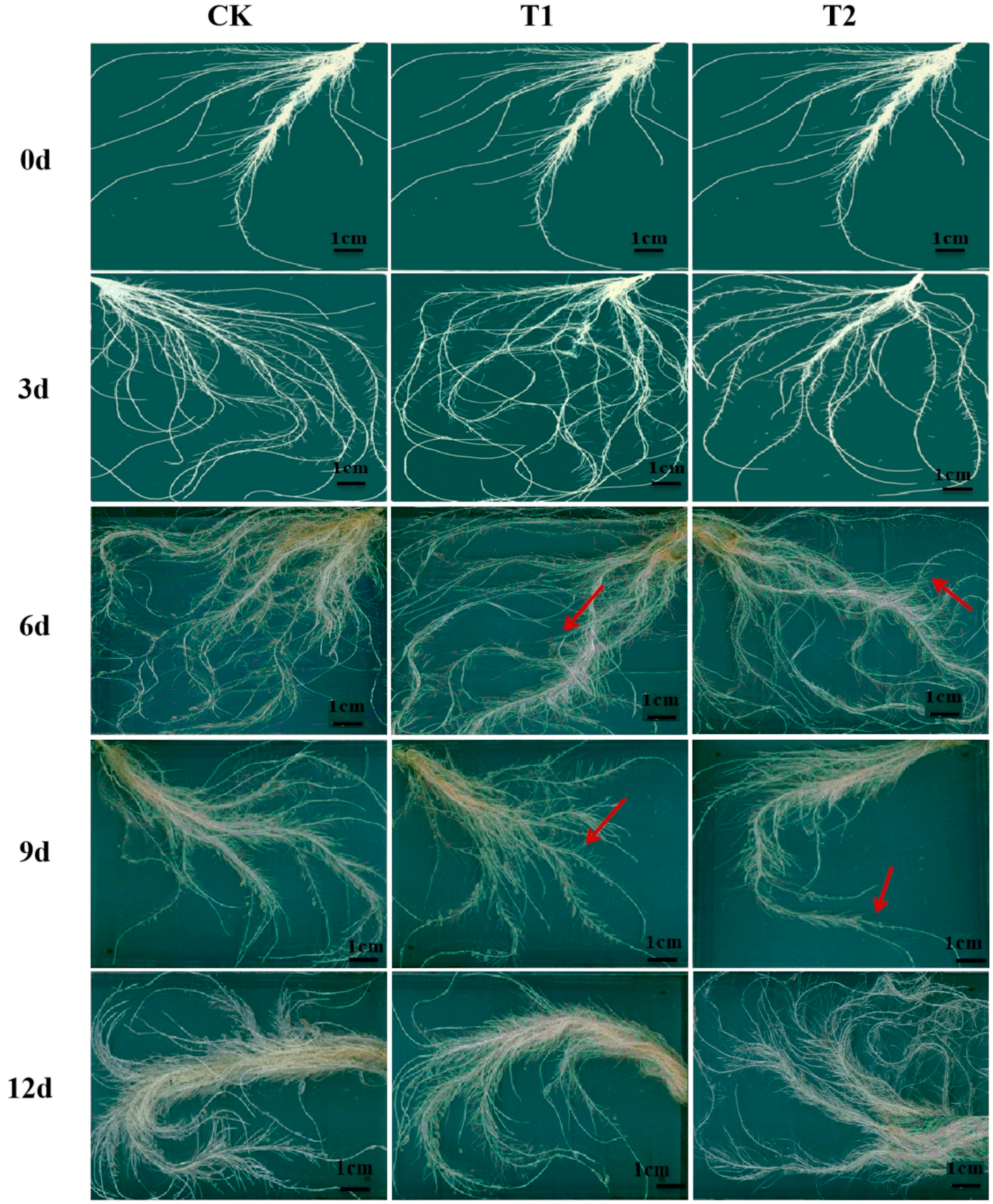
2.1. Effects of Elevated Root-Zone CO2 on Root Morphology
2.2. Effects of Elevated Root-Zone CO2 on Root Vigor, Root Volume, and Plant Biomass
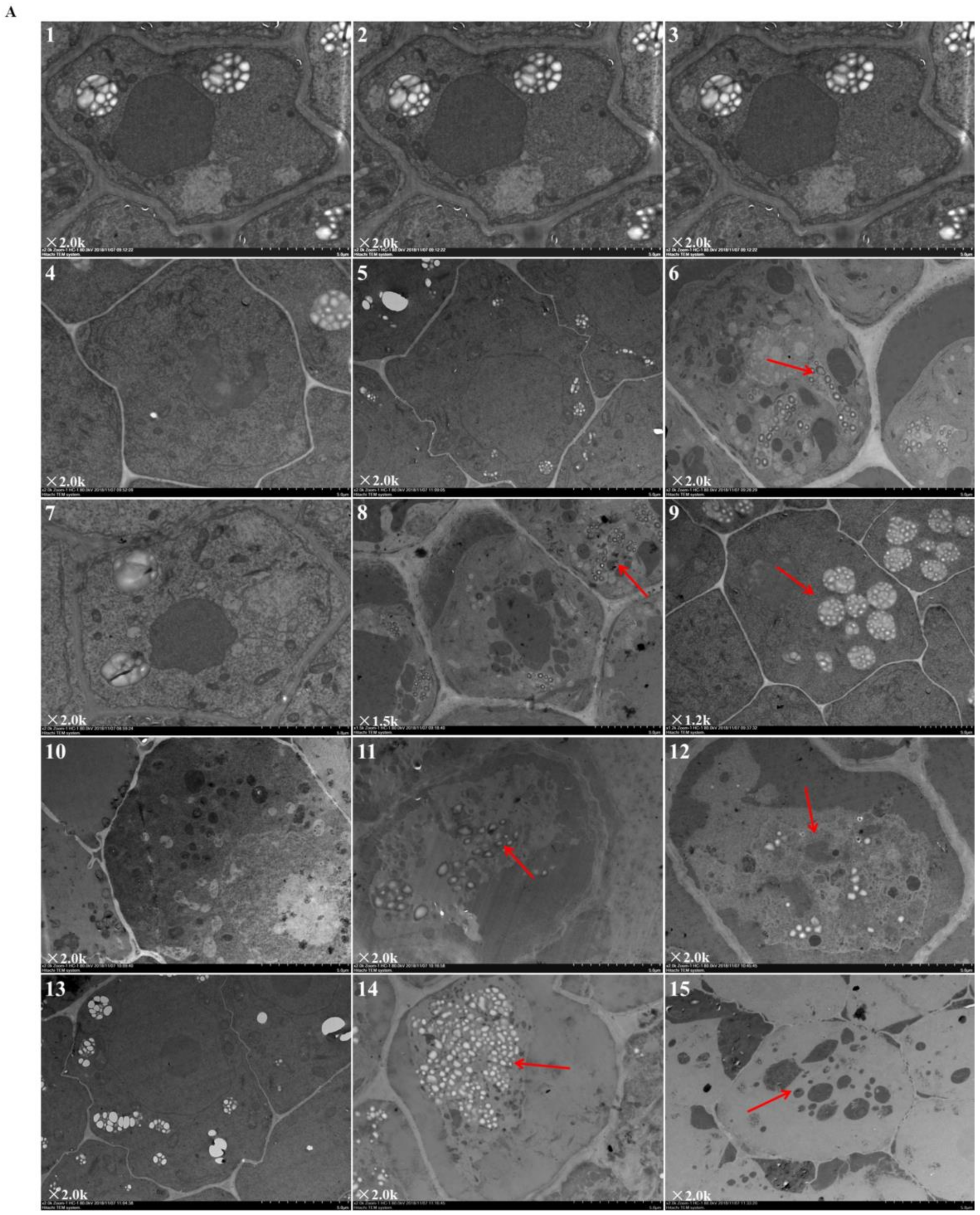
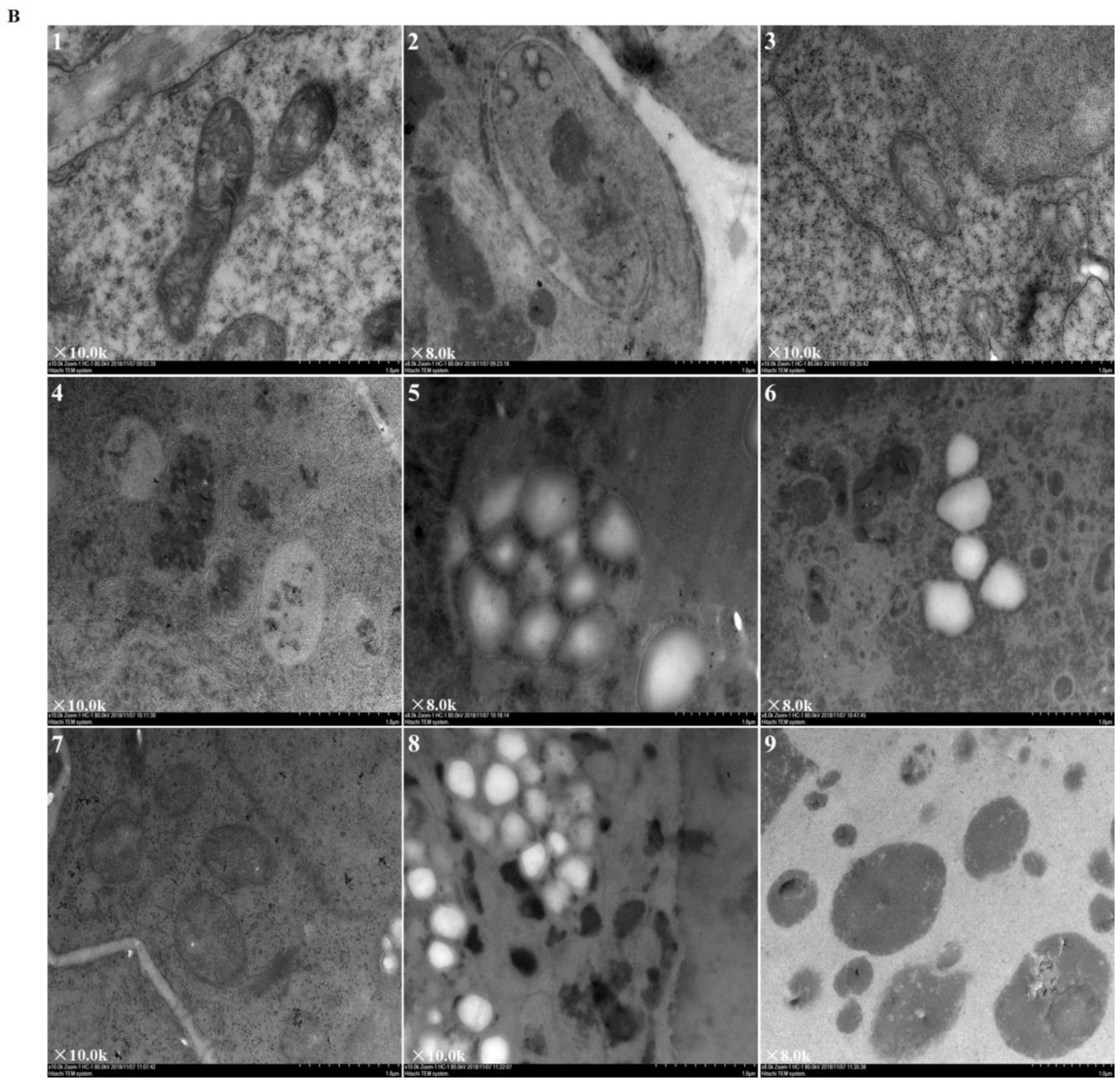
2.3. Effects of Elevated Root-Zone CO2 on Root Tip Cell Ultrastructure
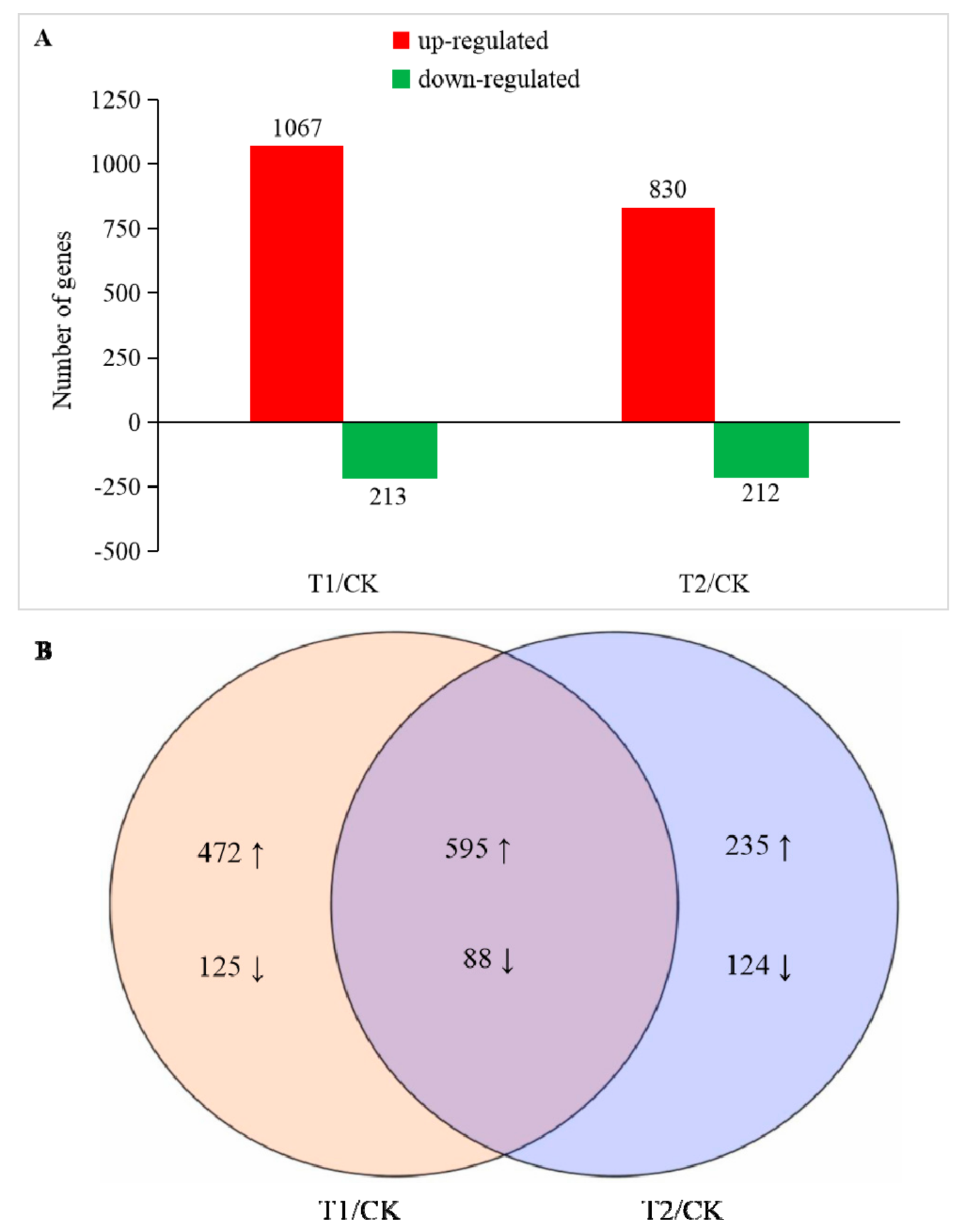
2.4. Identification of Differentially Expressed Genes (DEGs)
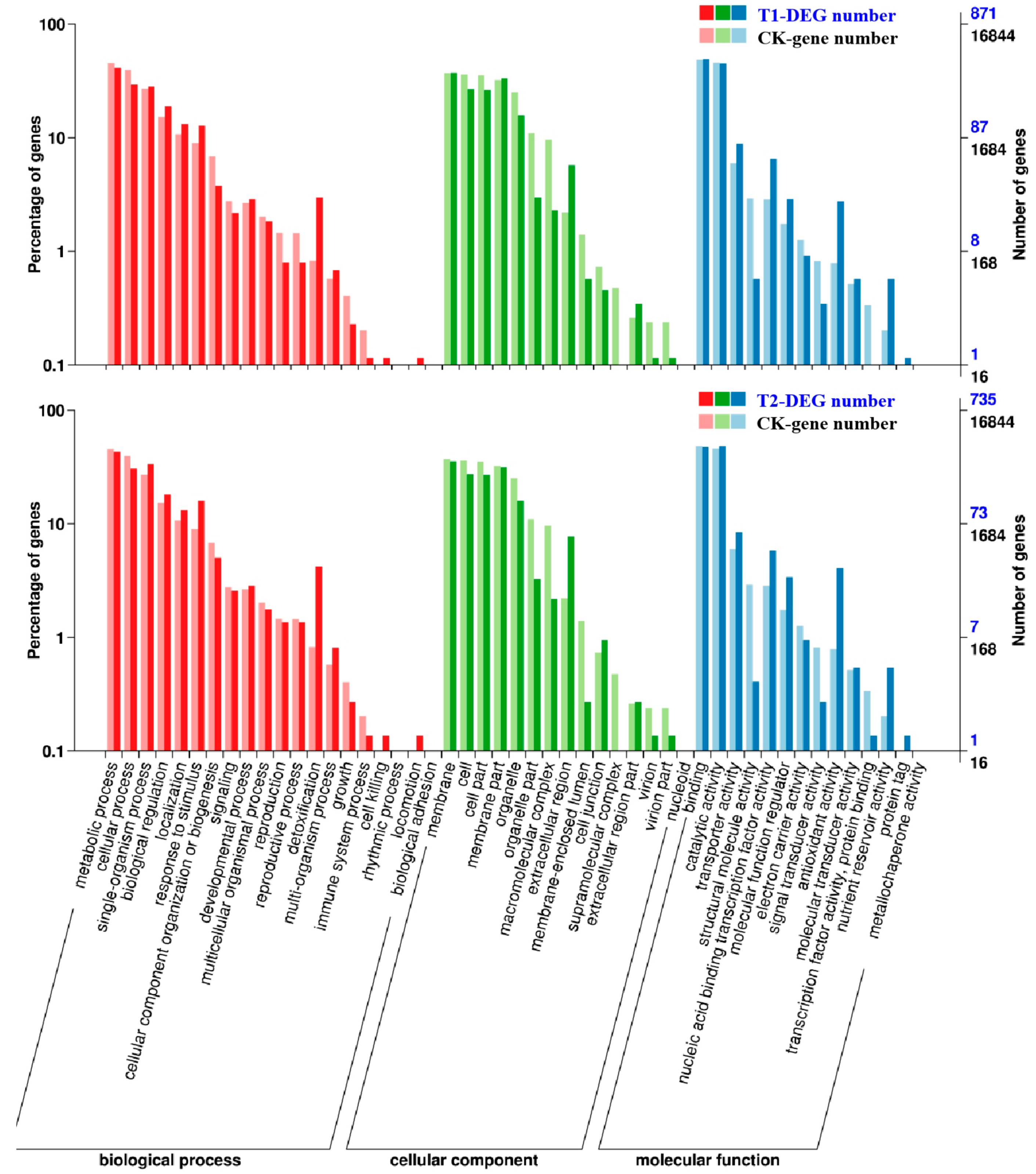
2.5. Functional Classification and Enrichment Analysis on Gene Ontology (GO)
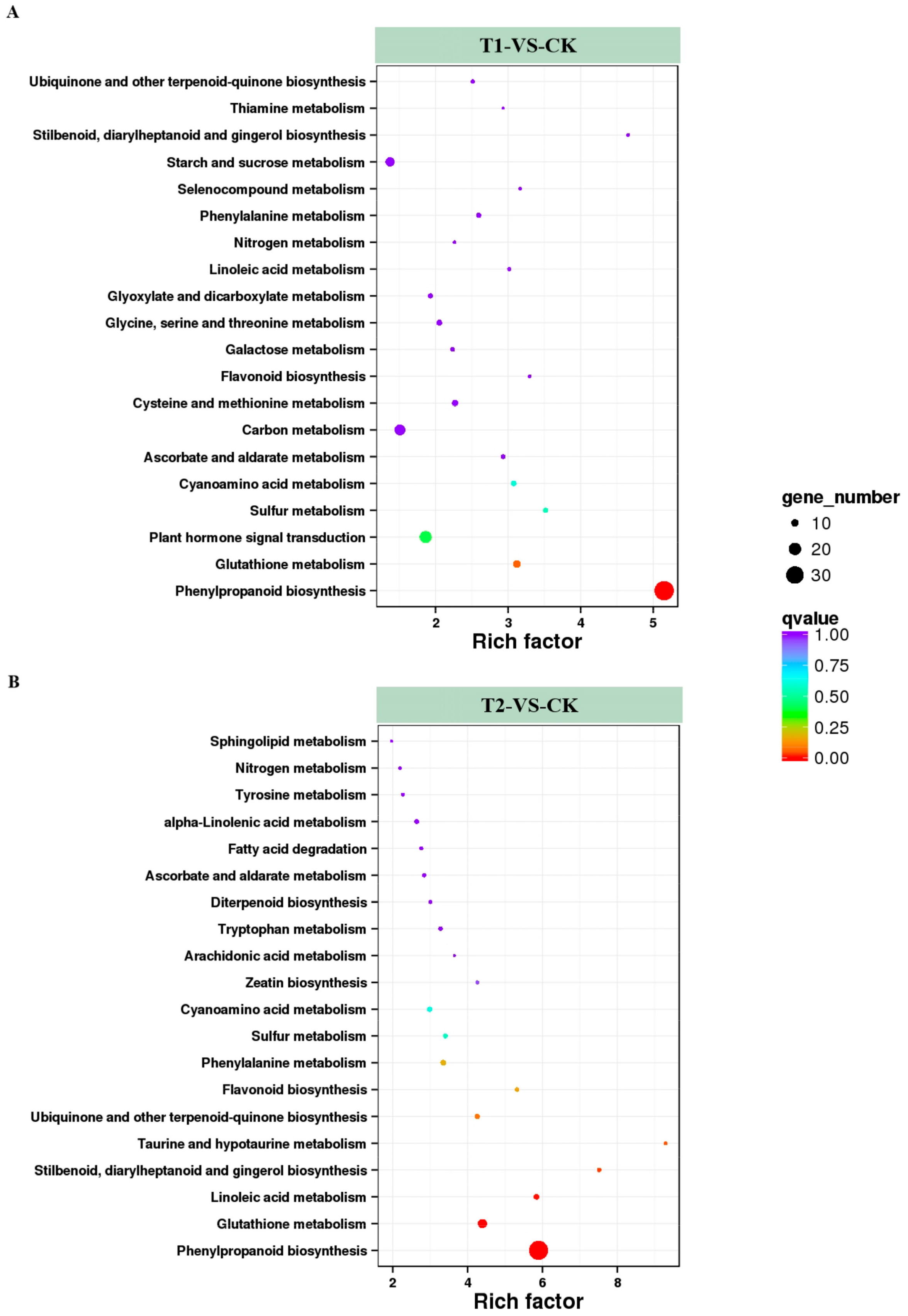
2.6. KEGG (Kyoto Encyclopedia of Genes and Genomes) Analysis of DEGs
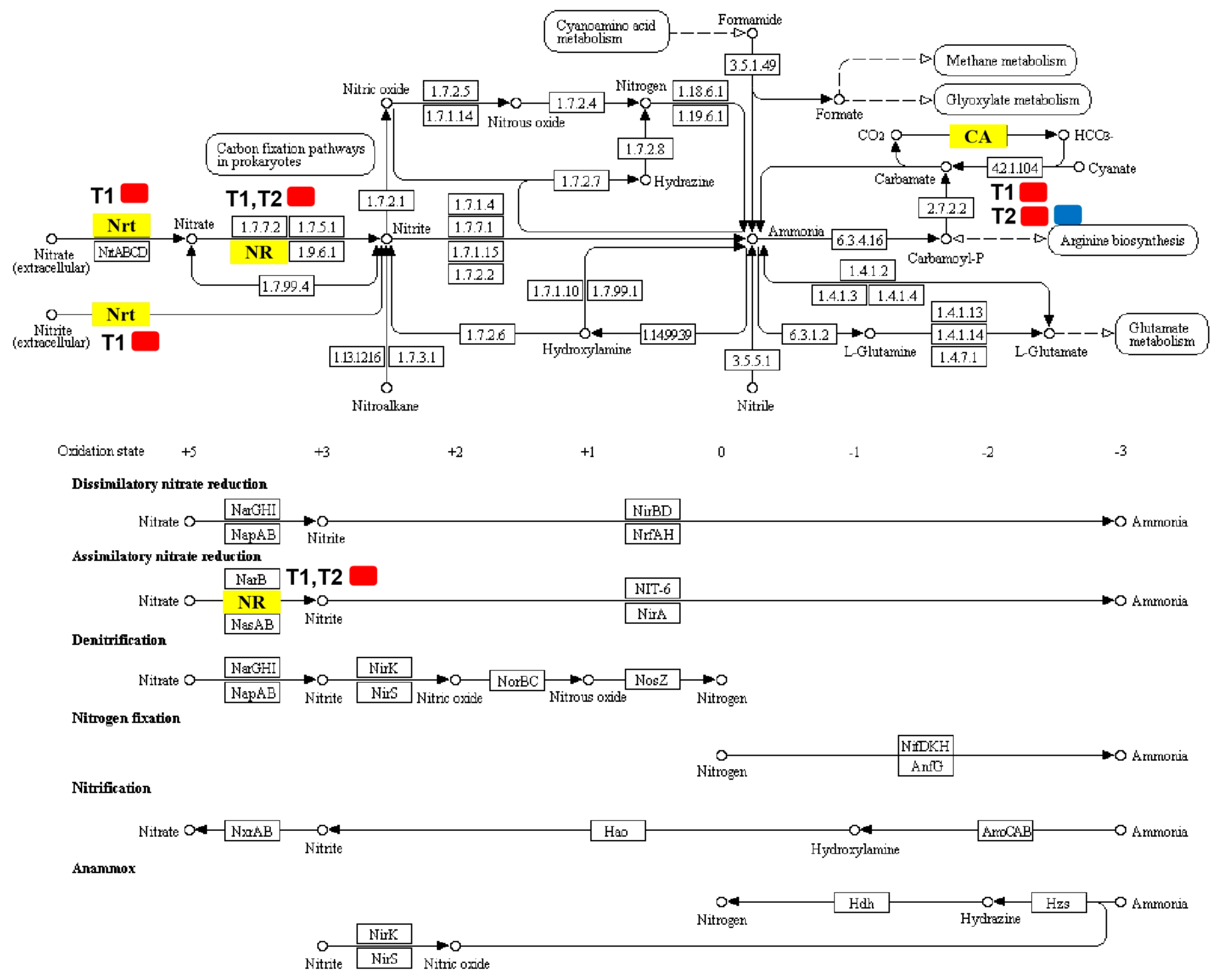
2.7. DEGs Involved in Nitrogen Metabolism
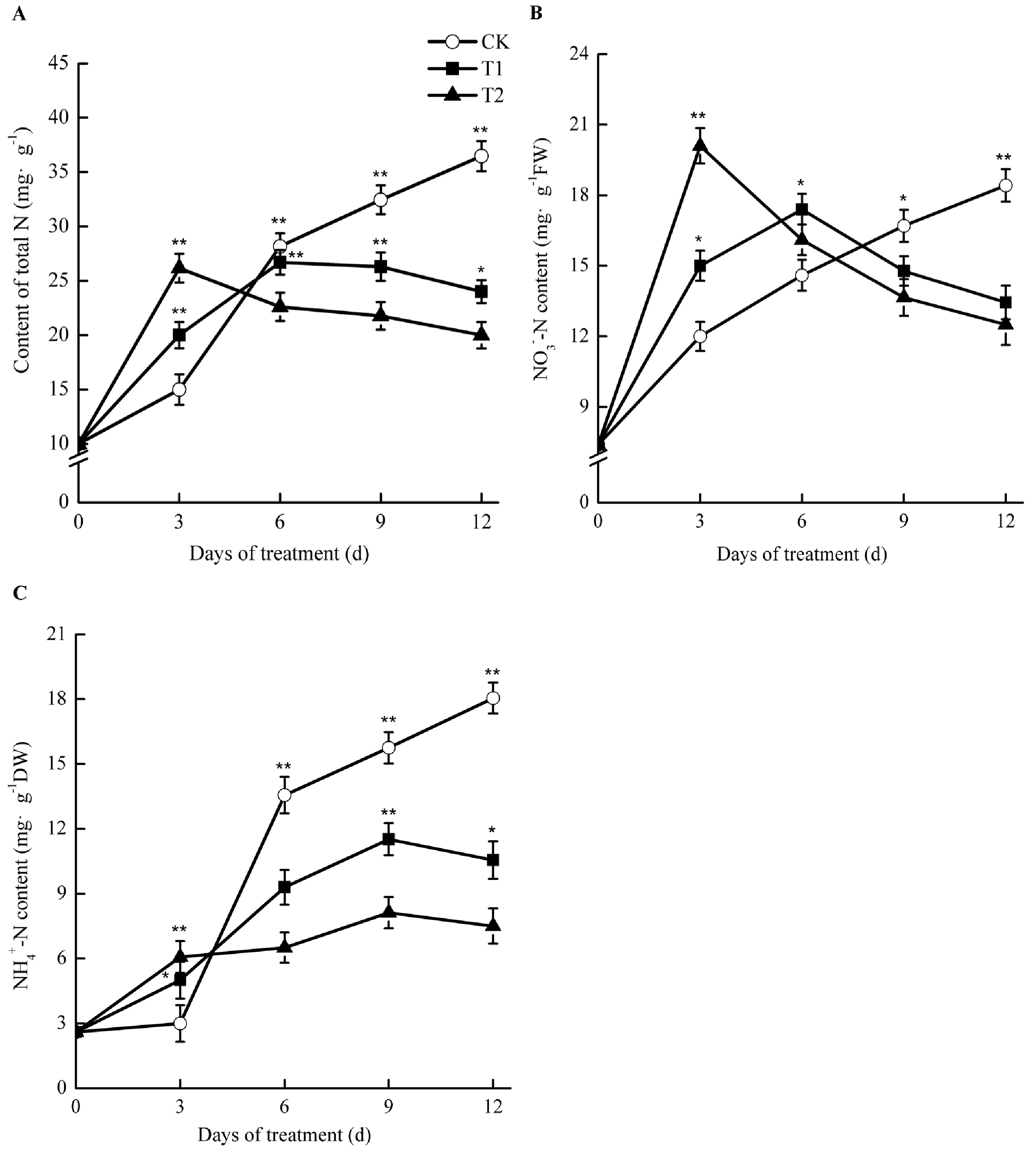
2.8. Effects of Elevated Root-Zone CO2 on Nitrogen Concentration
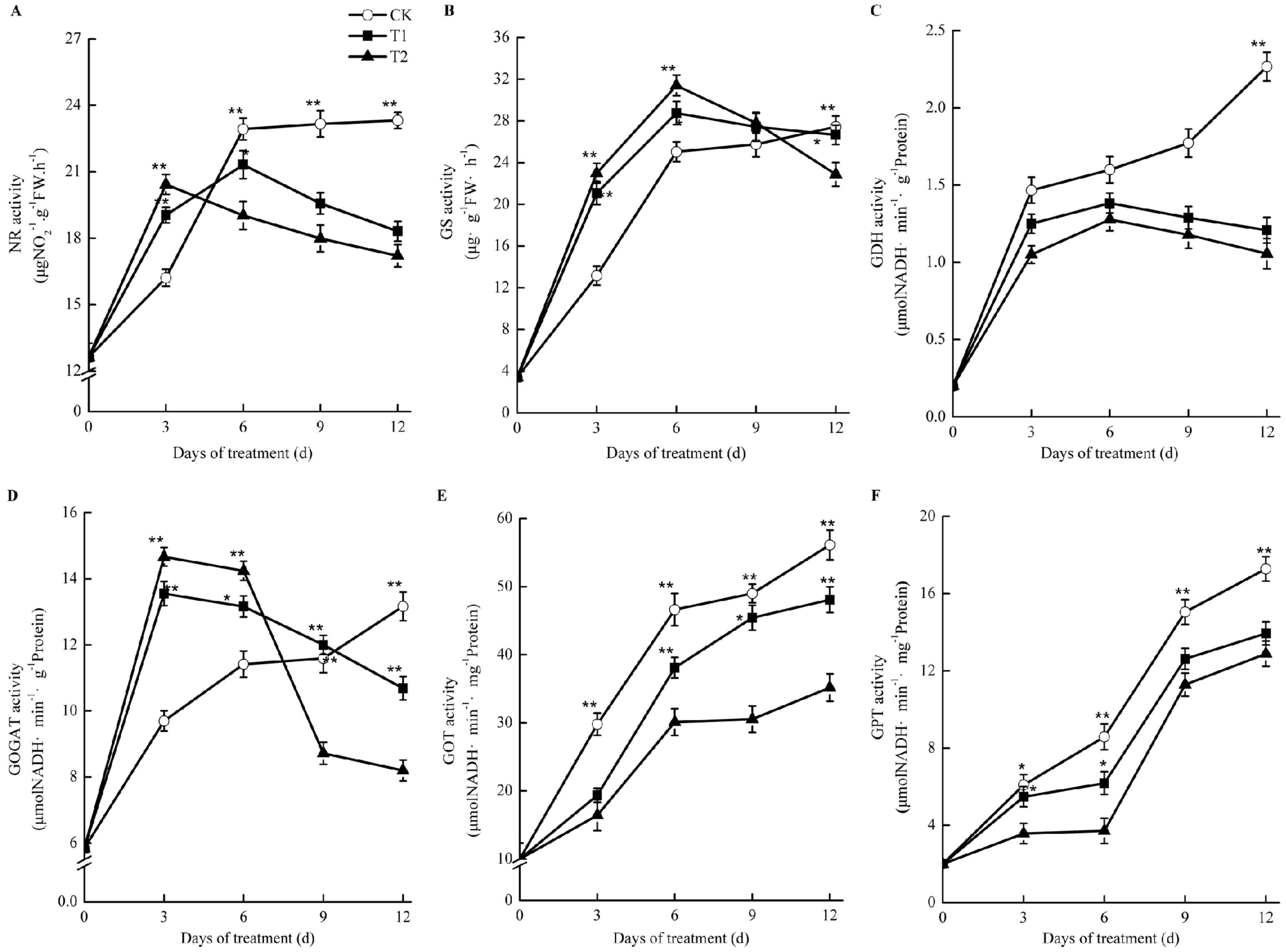
2.9. Effects of Elevated Root-Zone CO2 on Enzyme Activities Related to Nitrogen Metabolism
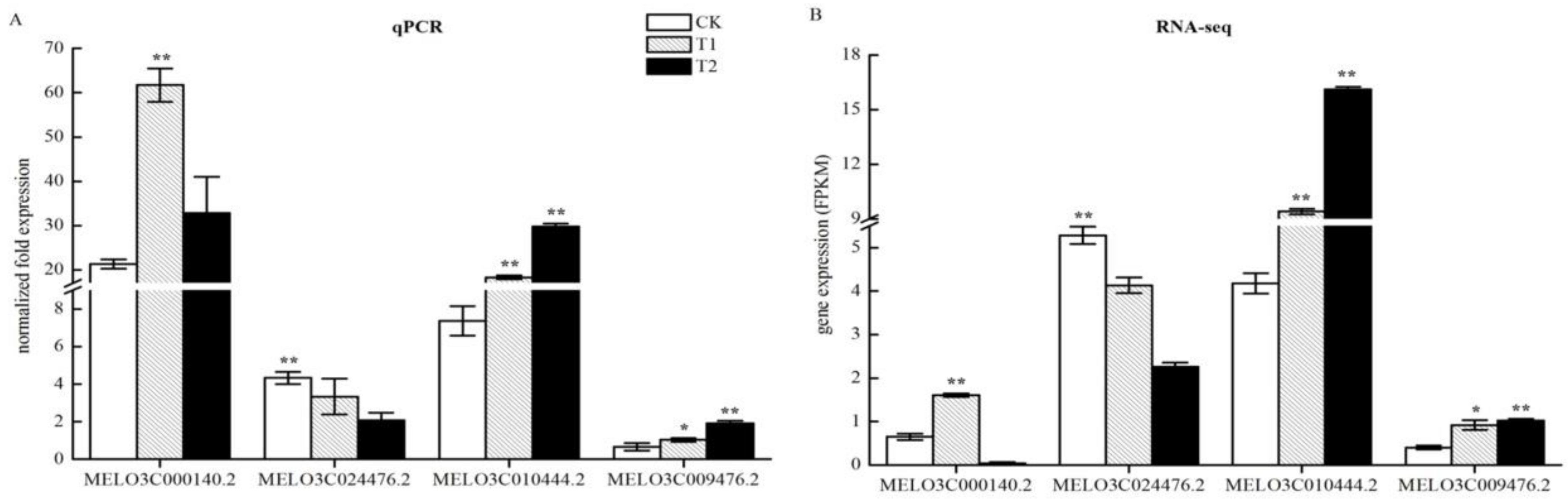
2.10. QPCR Verification of the Accuracy of Changes in Expression of DEGs Related to Nitrogen Metabolism
3. Discussion
3.1. Elevated Root-Zone CO2 Affected the Root Growth and Morphological Structure of Melon Seedlings
3.2. Elevated Root-Zone CO2 Affected the Accumulation and Metabolism of Nitrogen in Melon Seedlings Roots
4. Materials and Methods
4.1. Plant Material and Growth Conditions
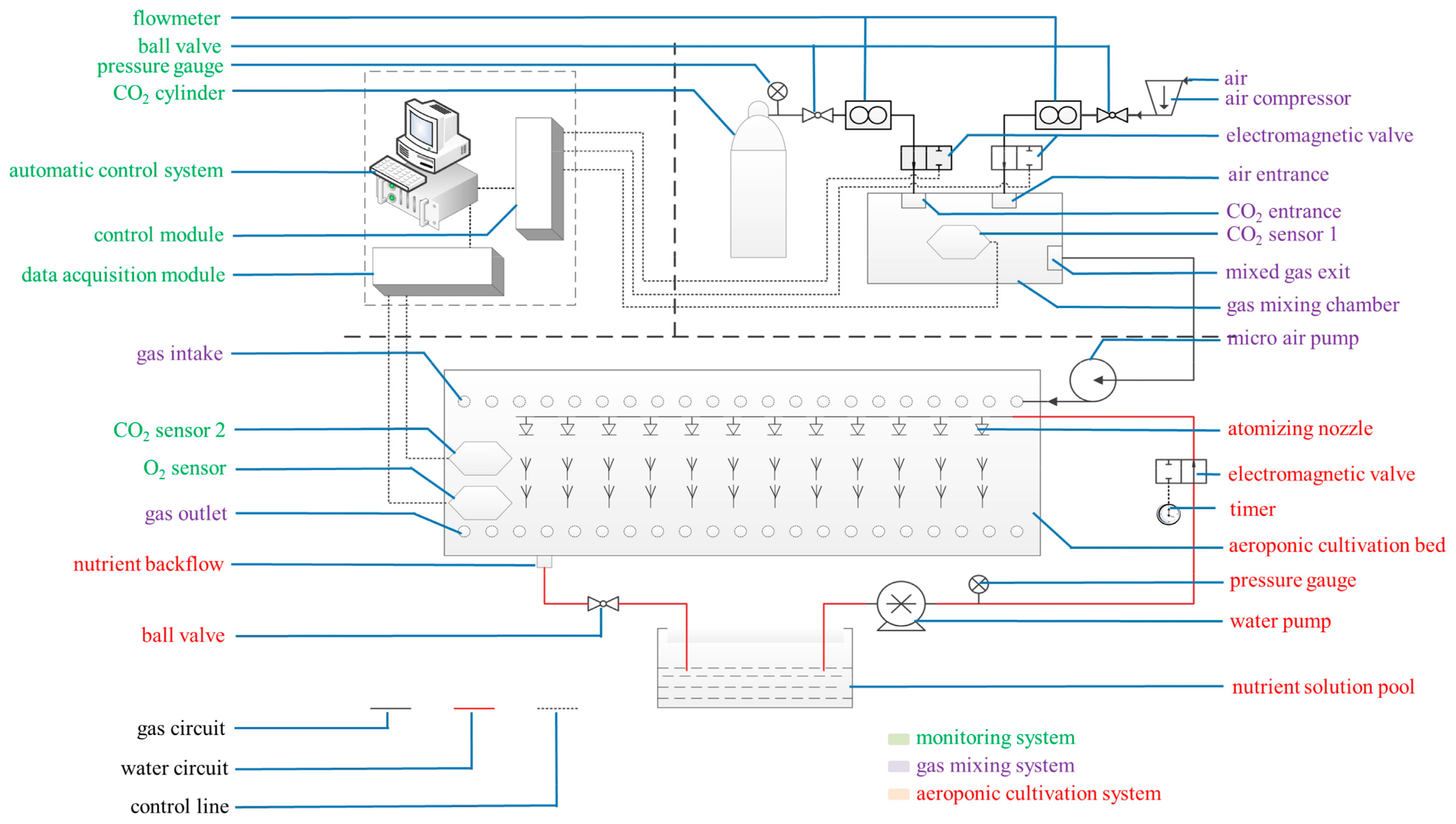
4.2. Root-Zone CO2 Concentration Treatments
4.3. Measurements of the Plant Growth Index
4.4. Analysis of Root Morphology
4.5. Observation of Root Tip Cell Ultrastructure
4.6. RNA Isolation, cDNA Library Construction, and Illumina Sequencing for Transcriptome Analysis
4.7. Identification of Differentially Expressed Genes (DEGs) and Functional Annotations
4.8. Determinations of NO3−, NH4+, and Total N Concentrations
4.9. Determination of Nitrate Reductase (NR), Glutamine Synthetase (GS), Glutamate Synthase (GOGAT), Glutamate Dehydrogenase (GDH), Glutamic Acid Oxaloacetate Transaminase (GOT), and Glutamic-Pyruvic Transaminase (GPT) Activities
4.10. Validation of Gene Expression Using qPCR
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Greenway, H.; Armstrong, W.; Colmer, T.D. Conditions leading to high CO2 (>5 kPa) in waterlogged–flooded soils and possible effects on root growth and metabolism. Ann. Bot. 2006, 98, 9–32. [Google Scholar] [CrossRef] [Green Version]
- Teskey, R.O.; Saveyn, A.; Steppe, K.; McGuire, M.A. Origin, fate and significance of CO2 in tree stems. New Phytol. 2008, 177, 17–32. [Google Scholar]
- Pumpanen, J.S.; Heinonsalo, J.; Rasilo, T.; Hurme, K.R.; Ilvesniemi, H. Carbon balance and allocation of assimilated CO2 in scots pine, norway spruce, and Silver birch seedlings determined with gas exchange measurements and 14C pulse labeling. Trees 2009, 23, 611–621. [Google Scholar] [CrossRef]
- Cramer, M.D.; Richards, M.B. The effect of rhizosphere dissolved inorganic carbon on gas exchange characteristics and growth rates of tomato seedlings. J. Exp. Bot. 1999, 50, 79–87. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, X.; Li, Y.E.; Wan, Y.F. The impacts of introduced CO2 flux on maize/alfalfa and soil. Int. J. Greenh. Gas Control 2014, 23, 86–97. [Google Scholar] [CrossRef]
- Lake, J.A.; Steven, M.D.; Smith, K.L.; Lomax, B.H. Plant responses to elevated CO2 levels in soils: Distinct CO2 and O2-depletion effects. Int. J. Greenh. Gas Control 2017, 64, 333–339. [Google Scholar] [CrossRef]
- Munir, R.; Konnerup, D.; Khan, H.A.; Siddique, K.H.M.; Colmer, T.D. Sensitivity of chickpea and faba bean to root-zone hypoxia, elevated ethylene, and carbon dioxide. Plant Cell Environ. 2019, 42, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Yoo, G.; Moonis, M.; Kim, Y.; Chen, X. Impact assessment of high soil CO2 on plant growth and soil environment: A greenhouse study. Peer J. 2019, 7, 6311–6328. [Google Scholar] [CrossRef]
- Van der Merwe, C.A.; Cramer, M.D. Effect of enriched rhizosphere carbon dioxide on nitrate and ammonium uptake in hydroponically grown tomato plants. Plant Soil 2000, 221, 5–11. [Google Scholar] [CrossRef]
- Viktor, A.; Cramer, M.D. Variation in root-zone CO2 concentration modifies isotopic fractionation of carbon and nitrogen in tomato seedlings. New Phytol. 2003, 157, 45–54. [Google Scholar] [CrossRef]
- He, J.; Austin, P.T.; Nichols, M.A.; Lee, S.K. Elevated root-zone CO2 protects lettuce plants from midday depression of photosynthesis. Environ. Exp. Bot. 2007, 61, 94–101. [Google Scholar] [CrossRef]
- He, J.; Austin, R.T.; Nichols, M.A.; Lee, S.K. Effect of root-zone (RZ) CO2, on productivity and photosynthesis in aeroponically grown lettuce plants. Acta Hortic. 2004, 648, 39–45. [Google Scholar] [CrossRef]
- He, J.; Austin, P.T.; Lee, S.K. Effects of elevated root zone CO2 and air temperature on photosynthetic gas exchange, nitrate uptake, and total reduced nitrogen content in aeroponically grown lettuce plants. J. Exp. Bot. 2010, 61, 3959–3969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boru, G.; Vantoai, T.; Alves, J.; Hua, D.; Knee, M. Response of soybean to oxygen deficiency and elevated root-zone carbon dioxide concentration. Ann. Bot. 2003, 91, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.D.; Savidov, N.A.; Lips, S.H. The influence of enriched rhizosphere CO2 on N uptake and metabolism in wild-type and NR-deficient barley plants. Physiol. Plant 1996, 97, 47–54. [Google Scholar] [CrossRef]
- Nichols, M.A.; Woolley, D.J.; Christie, C.B. Effect of oxygen and carbon dioxide concentration in the root zone on the growth of vegetables. Acta Hortic. 2002, 578, 119–122. [Google Scholar] [CrossRef]
- Cramer, M.D.; Shane, M.W.; Lambers, H. Physiological changes in white lupin associated with variation in root-zone CO2 concentration and cluster-root P mobilization. Plant Cell Environ. 2005, 28, 1203–1217. [Google Scholar] [CrossRef]
- Wang, C.; Fan, L.; Gao, H.; Wu, X.; Li, J.; Lv, G.; Gong, B. Polyamine biosynthesis and degradation are modulated by exogenous gamma-aminobutyric acid in root-zone hypoxia-stressed melon roots. Plant Physiol. Biochem. 2014, 82, 17–26. [Google Scholar] [CrossRef]
- Fan, L.; Wu, X.; Tian, Z.; Jia, K.; Pan, Y.; Li, J.; Gao, H. Comparative proteomic analysis of gamma-aminobutyric acid responses in hypoxia-treated and untreated melon roots. Phytochemistry 2015, 116, 28–37. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. Stringtie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.K.H.; McCouch, S. Getting to the roots of it: Genetic and hormonal control of root architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungk, A. Root hairs and the acquisition of plant nutrients from soil. J. Plant Nutr. Soil Sci. 2001, 164, 121–129. [Google Scholar] [CrossRef]
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant root growth, architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar] [CrossRef]
- He, J.; Qin, L.; Lee, S.K. Root morphology, plant growth, nitrate accumulation and nitrogen metabolism of temperate lettuce grown in the tropics with elevated root-zone CO2 at different root-zone temperatures. Am. J. Plant Sci. 2016, 7, 1821–1833. [Google Scholar] [CrossRef] [Green Version]
- Berveiller, D.; Damesin, C. Carbon assimilation by tree stems: Potential involvement of phosphoenolpyruvate carboxylase. Trees 2008, 22, 149–157. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How tree roots respond to drought. Front Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guern, J.; Felle, H.; Mathieu, Y.; Kurkdjian, A. Regulation of intracellular pH in plant cells. Int. Rev. Cytol. 1991, 127, 111–173. [Google Scholar]
- Heber, U.; Wagner, U.; Kaiser, W.; Neimanis, S.; Bailey, K.; Walker, D. Fast cytoplasmic pH regulation in acid-stressed leaves. Plant Cell Physiol. 1994, 35, 479–488. [Google Scholar]
- Davies, M.J. The oxidative environment and protein damage. Biochim. Biophys. Acta. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Naser, L.; Kourosh, V.; Bahman, K.; Reza, A. Soluble sugars and proline accumulation play a role as effective indices for drought tolerance screening in Persian walnut (Juglans regia l.) during germination. Fruits 2010, 65, 97–112. [Google Scholar] [CrossRef] [Green Version]
- Nikolopoulou, A.-E.; Nektarios, P.A.; Aivalakis, G.; Volterrani, M.; Chronopoulos, I. Effects of rootzone CO2 and O2 levels on seed germination and stolon growth of cynodon dactylon. Acta Agric. Scand. Sect. B Soil Plant Sci. 2012, 62, 53–61. [Google Scholar]
- Stitt, M.; Müller, C.; Matt, P.; Gibon, Y.; Carillo, P.; Morcuende, R.; Scheible, W.-R.; Krapp, A. Steps towards an integrated view of nitrogen metabolism. J. Exp. Bot. 2002, 53, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Crawford, N.M.; Coruzzi, G.M.; Tsay, Y.-F. Nitrate signaling: Adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Filleur, S.; Gan, Y.; Forde, B.G. Signaling mechanisms integrating root and shoot responses to changes in the nitrogen supply. Photosynth. Res. 2005, 83, 239–250. [Google Scholar] [CrossRef]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Functions of mineral nutrients: Macronutrients. In The Mineral Nutrition of Higher Plants; Marschner, H., Ed.; Academic Press: London, UK, 1995; pp. 229–255. [Google Scholar]
- De La Peña, T.C.; Frugier, F.; McKhann, H.I.; Bauer, P.; Brown, S.; Kondorosi, A.; Crespi, M. A carbonic anhydrase gene is induced in the nodule primordium and its cell-specific expression is controlled by the presence of rhizobium during development. Plant J. 1997, 11, 407–420. [Google Scholar] [CrossRef]
- Van der Westhuizen, M.M.; Cramer, M.D. The influence of elevated rhizosphere dissolved inorganic carbon concentrations on respiratory O2 and CO2 flux in tomato roots. J. Exp. Bot. 1998, 49, 1977–1985. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Cheng, W. Photosynthesis controls of rhizosphere respiration and organic matter decomposition. Soil Biol. Biochem. 2001, 33, 1915–1925. [Google Scholar] [CrossRef]
- Huber, S.C.; MacKintosh, C.; Kaiser, W.M. Metabolic enzymes as targets for 14-3-3 proteins. Plant Mol. Biol. 2002, 50, 1053–1063. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, B.; Harris, N.S.; Deyholos, M.K. Comparative proteomic analysis of NaCl stress-responsive proteins in arabidopsis roots. J. Exp. Bot. 2007, 58, 3591–3607. [Google Scholar] [CrossRef] [Green Version]
- Fredes, I.; Moreno, S.; Díaz, F.P.; Gutiérrez, R.A. Nitrate signaling and the control of arabidopsis growth and development. Curr. Opin. Plant Biol. 2019, 47, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Osugi, A.; Kojima, M.; Takebayashi, Y.; Ueda, N.; Kiba, T.; Sakakibara, H. Systemic transport of trans-zeatin and its precursor have differing roles in arabidopsis shoots. Nat. Plants 2017, 3, 17112. [Google Scholar] [CrossRef] [PubMed]
- Landrein, B.; Formosa-Jordan, P.; Malivert, A.; Schuster, C.; Melnyk, C.W.; Yang, W.; Turnbull, C.; Meyerowitz, E.M.; Locke, J.C.W.; Jönsson, H. Nitrate modulates stem cell dynamics in arabidopsis shoot meristems through cytokinins. Proc. Natl. Acad. Sci. USA 2018, 115, 1382–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Pan, R.; Tan, L.; Zhang, Z.; Guo, M. Pleiotropic roles of O-mannosyltransferase MoPmt4 in development and pathogenicity of Magnaporthe oryzae. Curr. Genet. 2019, 65, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Knievel, D.P. Procedure for estimating ratio of live to dead root dry matter in root core samples. Crop Sci. 1973, 13, 124–126. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Tan, L.P.; He, J.; Lee, S.K. Effects of root-zone temperature on the root development and nutrient uptake of lactuca sativa L. “Panama” grown in an aeroponic system in the tropics. J. Plant Nutr. 2002, 25, 297–314. [Google Scholar] [CrossRef]
- Bräutigam, A.; Gagneul, D.; Weber, A.P.M. High-throughput colorimetric method for the parallel assay of glyoxylic acid and ammonium in a single extract. Anal. Biochem. 2007, 362, 151–153. [Google Scholar] [CrossRef]
- Kaiser, W.M.; Huber, S.C. Correlation between apparent activation state of nitrate reductase (NR), NR hysteresis and degradation of NR protein. J. Exp. Bot. 1997, 48, 1367–1374. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, Q.; Ding, L.; Sun, Y. Effect of cadmium toxicity on nitrogen metabolism in leaves of solanum nigrum l. as a newly found cadmium hyperaccumulator. J. Hazard Mater. 2008, 154, 818–825. [Google Scholar] [CrossRef]
- Lin, C.C.; Kao, C.H. Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl. Plant Growth Regul. 1996, 18, 233–238. [Google Scholar] [CrossRef]
- Kaur, B.; Asthir, B.; Bains, N.S. Apical stem culturing to enhance cell sap assimilates towards grain sucrose and glutamine metabolism in wheat. Cereal Res. Commun. 2015, 43, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root Morphology Index | Days of Treatment (day) | Treatment | ||
|---|---|---|---|---|
| CK | T1 | T2 | ||
| The main root length (cm) | 0 | 30.00 ± 3.47 a | 30.00 ± 3.47 a | 30.00 ± 3.47 a |
| 3 | 40.33 ± 2.09 b | 52.10 ± 2.17 a | 54.67 ± 2.27 a | |
| 6 | 65.25 ± 4.33 a | 61.07 ± 2.31 a | 59.67 ± 2.23 a | |
| 9 | 90.83 ± 1.19 a | 83.17 ± 1.44 b | 71.85 ± 1.67 c | |
| 12 | 117.23 ± 2.21 a | 103.8 ± 0.92 b | 89.17 ± 0.98 c | |
| Total root length (cm) | 0 | 72.53 ± 5.52 a | 72.53 ± 5.52 a | 72.53 ± 5.52 a |
| 3 | 138.23 ± 6.52 b | 153.19 ± 5.34 a | 165.85 ± 5.25 a | |
| 6 | 228.64 ± 6.26 a | 233.06 ± 5.85 a | 186.56 ± 5.35 b | |
| 9 | 257.07 ± 5.28 a | 245.21 ± 6.13 b | 207.31 ± 5.98 c | |
| 12 | 321.61 ± 5.69 a | 299.65 ± 6.61 b | 269.09 ± 5.27 c | |
| Root surface area (cm2) | 0 | 25.35 ± 0.12 a | 25.35 ± 0.12 a | 25.35 ± 0.12 a |
| 3 | 30.36 ± 2.45 b | 35.63 ± 2.20 a | 37.94 ± 3.30 a | |
| 6 | 51.46 ± 6.96 a | 56.00 ± 6.70 a | 50.44 ± 4.63 a | |
| 9 | 128.75 ± 7.33 a | 102.55 ± 7.01 b | 61.74 ± 5.18 c | |
| 12 | 153.61 ± 7.83 a | 123.69 ± 7.43 b | 80.98 ± 5.54 c | |
| Total root tip numbers | 0 | 81.00 ± 2.21 a | 81.00 ± 2.21 a | 81.00 ± 2.21 a |
| 3 | 125.50 ± 5.30 b | 156.80 ± 8.93 a | 152.70 ± 11.80 a | |
| 6 | 202.00 ± 7.86 c | 233.00 ± 10.08 b | 236.00 ± 8.62 a | |
| 9 | 254.00 ± 6.36 a | 245.00 ± 9.86 b | 243.00 ± 8.09 b | |
| 12 | 330.00 ± 15.72 a | 247.00 ± 11.28 b | 212.00 ± 9.05 c | |
| Number of root tip 0–0.5 mm in diameter | 0 | 22.83 ± 2.99 a | 22.83 ± 2.99 a | 22.83 ± 2.99 a |
| 3 | 34.23 ± 2.99 b | 36.52 ± 3.39 b | 45.99 ± 2.75 a | |
| 6 | 86.77 ± 2.22 a | 72.65 ± 2.46 b | 51.37 ± 2.13 c | |
| 9 | 98.49 ± 3.10 a | 88.93 ± 5.17 b | 77.01 ± 2.45 b | |
| 12 | 118.95 ± 5.70 a | 98.58 ± 5.10 b | 85.68 ± 2.60 c | |
| Number of root tip 0.5–2 mm in diameter | 0 | 38.29 ± 3.85 a | 38.29 ± 3.85 a | 38.29 ± 3.85 a |
| 3 | 65.30 ± 3.99 a | 45.89 ± 3.85 b | 47.82 ± 4.73 b | |
| 6 | 115.99 ± 5.37 a | 105.18 ± 3.33 a | 102.78 ± 2.54 a | |
| 9 | 125.87 ± 4.10 b | 122.58 ± 5.17 b | 146.29 ± 4.12 a | |
| 12 | 150.17 ± 4.70 b | 168.22 ± 4.10 b | 210.88 ± 4.60 a | |
| Growth Index | Days of Treatment (day) | Treatment | ||
|---|---|---|---|---|
| CK | T1 | T2 | ||
| Root vigor (μg·g−1·h−1) | 0 | 9.47 ± 0.87 a | 9.47 ± 0.88 a | 9.47 ± 0.89 a |
| 3 | 11.49 ± 0.70 b | 13.01 ± 0.74 a | 14.59 ± 0.82 a | |
| 6 | 12.56 ± 0.67 b | 15.52 ± 0.66 a | 16.57 ± 0.57 a | |
| 9 | 16.81 ± 0.69 a | 16.13 ± 0.54 a | 17.47 ± 0.70 a | |
| 12 | 22.79 ± 0.83 a | 17.85 ± 0.76 b | 18.39 ± 0.59 b | |
| Root volume (cm3) | 0 | 2.00 ± 0.06 a | 2.00 ± 0.06 a | 2.00 ± 0.06 a |
| 3 | 3.33 ± 0.38 b | 4.00 ± 0.38 a | 5.67 ± 0.01 a | |
| 6 | 9.67 ± 1.73 b | 10.00 ± 0.01 a | 8.67 ± 0.02 c | |
| 9 | 15.67 ± 1.15 a | 13.00 ± 1.01 b | 11.67 ± 1.05 b | |
| 12 | 36.67 ± 1.67 a | 33.33 ± 1.72 b | 28.33 ± 0.96 c | |
| Shoot dry weight (g) | 0 | 0.52 ± 0.07 a | 0.52 ± 0.07 a | 0.52 ± 0.07 a |
| 3 | 0.71 ± 0.00 a | 0.67 ± 0.00 a | 0.58 ± 0.07 b | |
| 6 | 1.43 ± 0.08 a | 1.38 ± 0.01 a | 1.11 ± 0.09 b | |
| 9 | 2.20 ± 0.06 a | 1.97 ± 0.25 a | 1.53 ± 0.16 b | |
| 12 | 4.30 ± 0.26 a | 3.20 ± 0.39 b | 2.27 ± 0.21 c | |
| Root dry weight (g) | 0 | 0.12 ± 0.03 a | 0.12 ± 0.03 a | 0.12 ± 0.03 a |
| 3 | 0.21 ± 0.00 a | 0.20 ± 0.00 a | 0.21 ± 0.02 a | |
| 6 | 0.36 ± 0.04 b | 0.42 ± 0.04 a | 0.39 ± 0.08 a | |
| 9 | 0.60 ± 0.05 a | 0.41 ± 0.10 b | 0.33 ± 0.07 c | |
| 12 | 1.07 ± 0.07 a | 0.82 ± 0.08 b | 0.54 ± 0.08 c | |
| Different Expressed Genes | T1/CK | |||
|---|---|---|---|---|
| Gene ID | Gene Name | FDR | Log2FoldChange | Regulation |
| MELO3C000140.2 | High-affinity nitrate transporter 2.2 | 0.007581 | 1.219896 | Up |
| MELO3C010444.2 | Nitrate reductase | 5.21 ± 10−9 | 1.085544 | Up |
| MELO3C009476.2 | Alpha carbonic anhydrase | 0.030669 | 1.104078 | Up |
| Different Expressed Genes | T2/CK | |||
| Gene ID | Gene Name | FDR | Log2FoldChange | Regulation |
| MELO3C010444.2 | Nitrate reductase | 4.40 ± 10−31 | 1.903265 | Up |
| MELO3C009476.2 | Alpha carbonic anhydrase | 0.009895 | 1.286820 | Up |
| MELO3C024476.2 | Alpha carbonic anhydrase 7-like | 9.99 ± 10−5 | −1.256327 | Down |
| Gene ID | Sequence (5′-3′) | |
|---|---|---|
| actin | F | AAGGCAAACAGGGAGAAGATGA |
| R | AGCAAGGTCGAGACGTAGGATA | |
| MELO3C000140.2 | F | GATGGCGAATCTAGTGGCTAG |
| R | GTCCACAACCTTCCTCTCATC | |
| MELO3C010444.2 | F | TGGTGTATGCGAATAGAACGG |
| R | CACACTATATTCCCACCCTTCC | |
| MELO3C009476.2 | F | CTTCATCTAGTCCATCAGGCAG |
| R | GTGCTTTGTGTCTAGGTCTCC | |
| MELO3C024476.2 | F | GAAGATAAGGACAGTGTCAAGG |
| R | GAGTTGGCCGATAGAGACCG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Yin, Z.; Yin, Y.; Xu, C.; Wang, W.; Liu, Y.; Li, T. Effects of Elevated Root-Zone CO2 on Root Morphology and Nitrogen Metabolism Revealed by Physiological and Transcriptome Analysis in Oriental Melon Seedling Roots. Int. J. Mol. Sci. 2020, 21, 803. https://doi.org/10.3390/ijms21030803
Chen X, Yin Z, Yin Y, Xu C, Wang W, Liu Y, Li T. Effects of Elevated Root-Zone CO2 on Root Morphology and Nitrogen Metabolism Revealed by Physiological and Transcriptome Analysis in Oriental Melon Seedling Roots. International Journal of Molecular Sciences. 2020; 21(3):803. https://doi.org/10.3390/ijms21030803
Chicago/Turabian StyleChen, Xinyu, Zepeng Yin, Yang Yin, Chuanqiang Xu, Wanxin Wang, Yiling Liu, and Tianlai Li. 2020. "Effects of Elevated Root-Zone CO2 on Root Morphology and Nitrogen Metabolism Revealed by Physiological and Transcriptome Analysis in Oriental Melon Seedling Roots" International Journal of Molecular Sciences 21, no. 3: 803. https://doi.org/10.3390/ijms21030803
APA StyleChen, X., Yin, Z., Yin, Y., Xu, C., Wang, W., Liu, Y., & Li, T. (2020). Effects of Elevated Root-Zone CO2 on Root Morphology and Nitrogen Metabolism Revealed by Physiological and Transcriptome Analysis in Oriental Melon Seedling Roots. International Journal of Molecular Sciences, 21(3), 803. https://doi.org/10.3390/ijms21030803