The Biofunctional Effects of Mesima as a Radiosensitizer for Hepatocellular Carcinoma

Abstract
:1. Introduction
2. Results
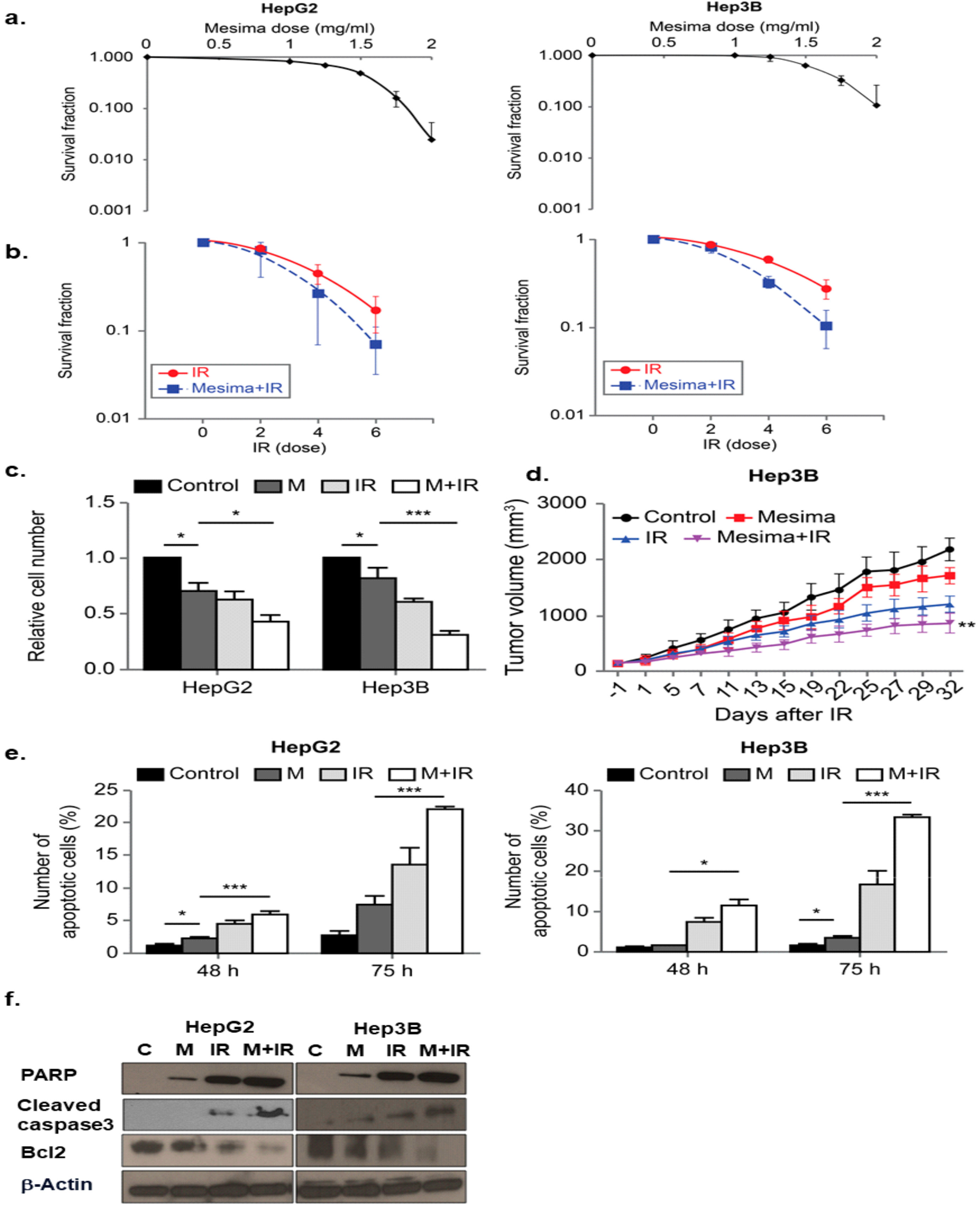
2.1. Mesima Sensitized HCC to Radiation In Vitro and In Vivo
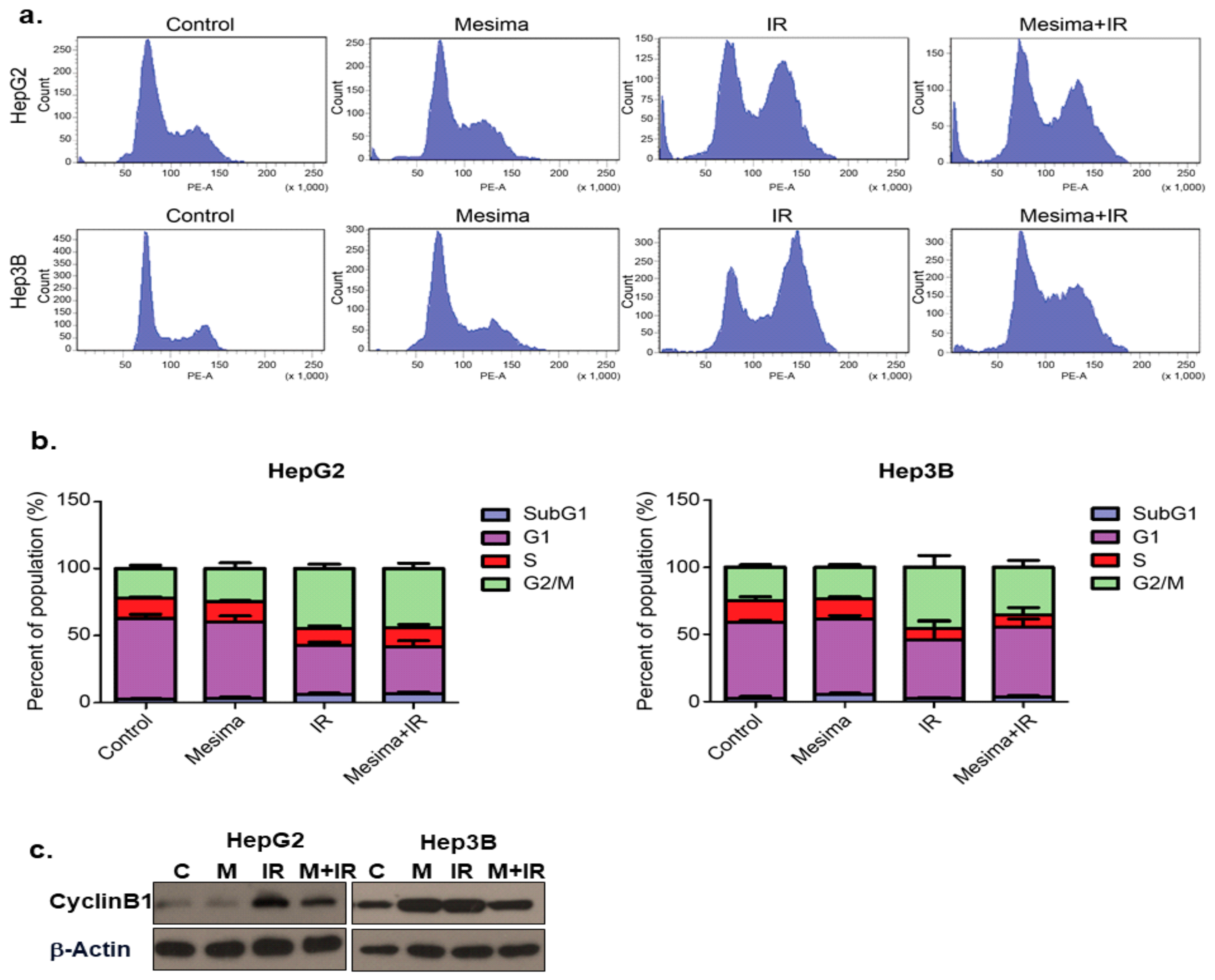
2.2. Effects of Mesima and Radiation on the Cell Cycle
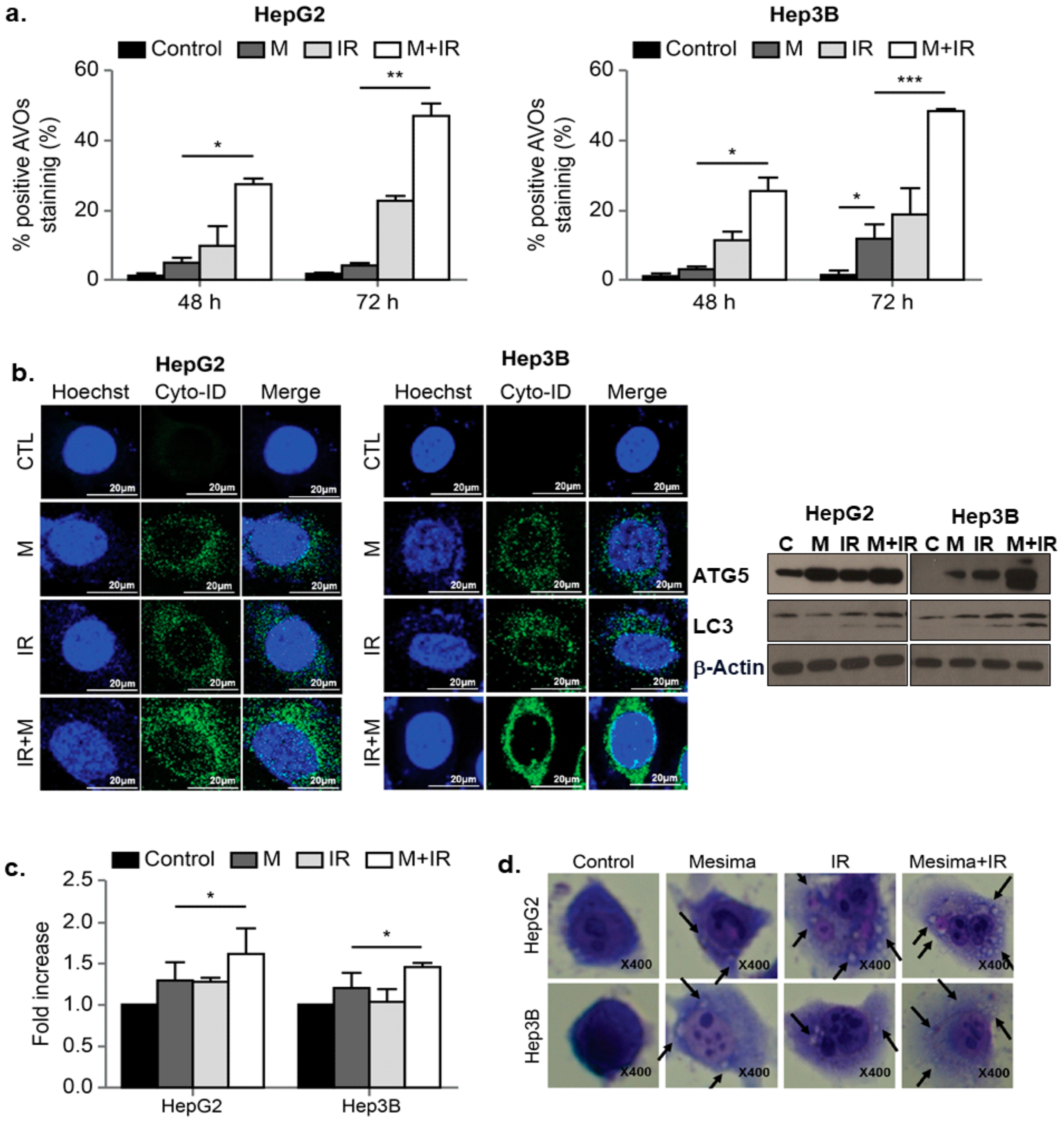
2.3. Effects of Mesima and Radiation on Autophagic Cell Death
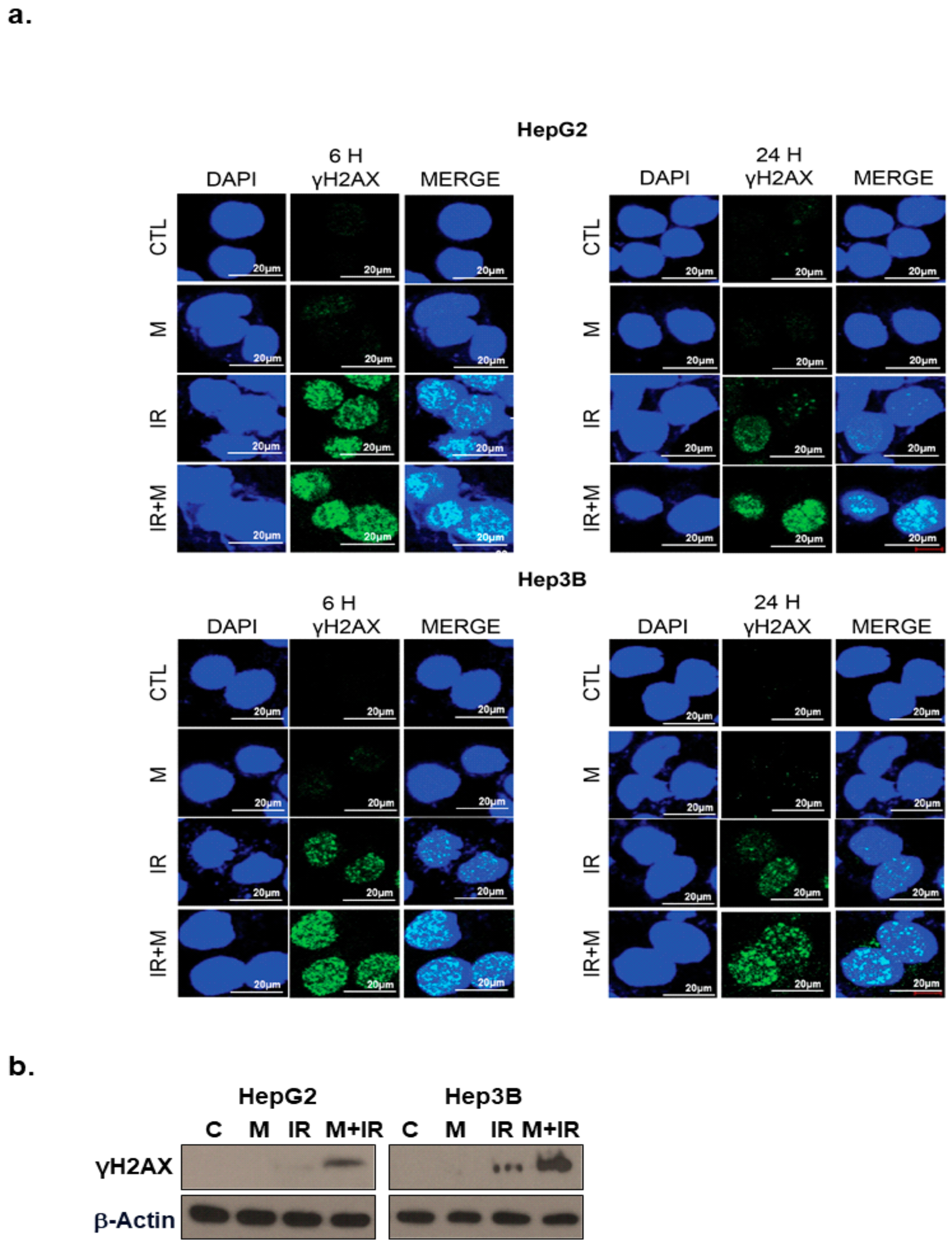
2.4. Effects of Mesima and Radiation on DNA Damage
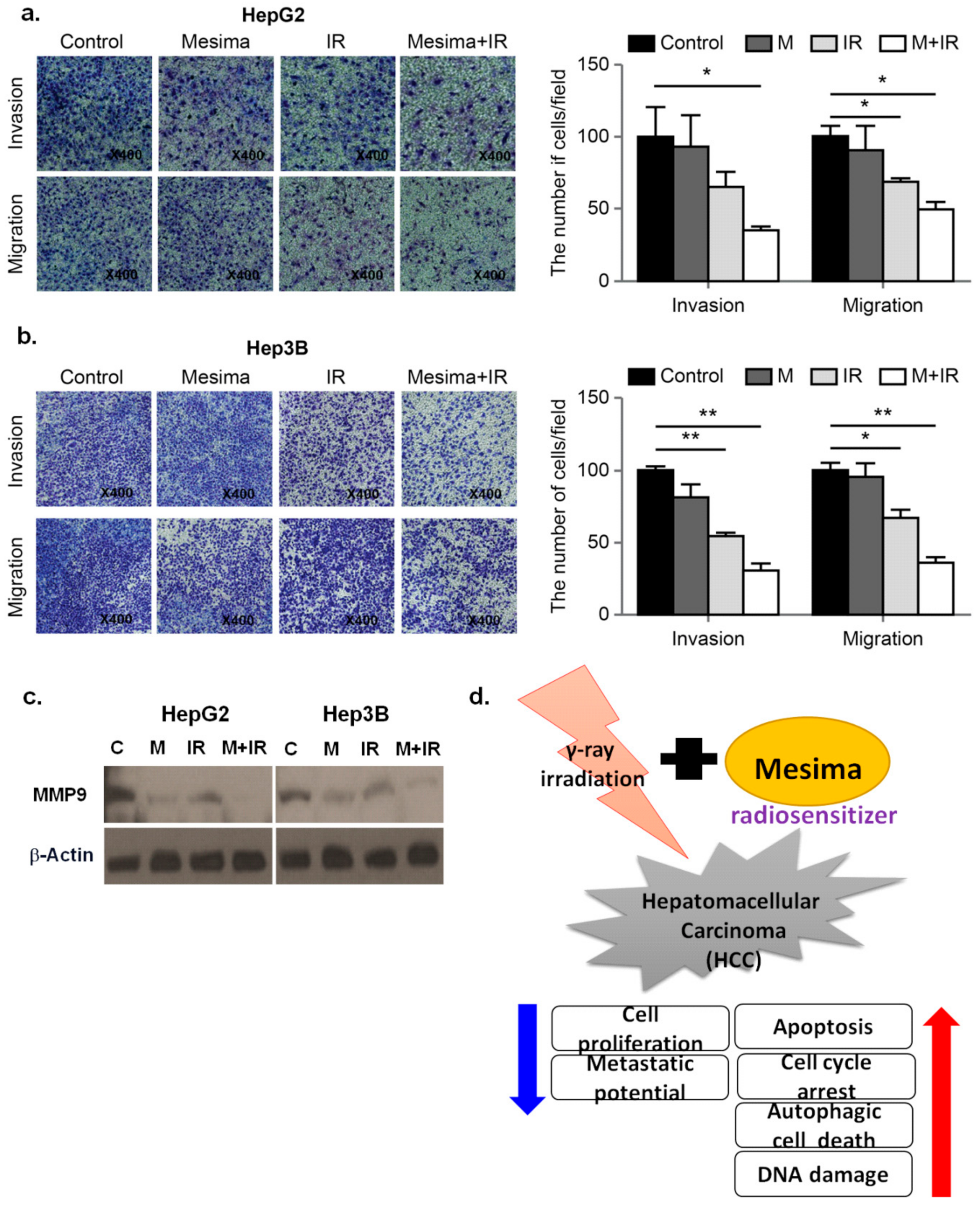
2.5. Combination Therapy Significantly Inhibited Tumor Cell Motility and Invasion
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Irradiation
4.3. Western Blotting
4.4. Antibodies and Chemicals
4.5. Colony-Forming Assay
4.6. Xenograft Tumors in Nude Mice
4.7. Detection of Apoptotic Cells by Annexin V Staining
4.8. Analysis of Cell Cycle Progression
4.9. Quantification of AVOs by Acridine Orange (AO) Staining
4.10. Immunofluorescence
4.11. Morphology
4.12. Transwell Chamber Assay
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Parkin, D.M. Global cancer statistics in the year 2000. Lancet Oncol. 2001, 2, 533–543. [Google Scholar] [CrossRef]
- El–Serag, H.B.; Rudolph, K.L. Hepatocellular carcinoma: Epidemiology and molecular carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar] [CrossRef] [PubMed]
- Bruix, J.; Sherman, M.J.H. Management of hepatocellular carcinoma. Hepatology 2005, 42, 1208–1236. [Google Scholar] [CrossRef] [PubMed]
- Sliva, D. Medicinal mushroom Phellinus linteus as an alternative cancer therapy. Exp. Ther. Med. 2010, 1, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-S.; Cho, S.-M.; Lee, J.; Kim, H.-M.; Han, S.-B.; Ko, K.-S.; Yoo, I.-D. B-lymphocyte-stimulating polysaccharide from mushroom Phellinus linteus. Chem. Pharm. Bull. 1995, 43, 2105–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.B.; Lee, C.W.; Jeon, Y.J.; Hong, N.D.; Yoo, I.D.; Yang, K.-H.; Kim, H.M. The inhibitory effect of polysaccharides isolated from Phellinus linteus on tumor growth and metastasis. Immunopharmacology 1999, 41, 157–164. [Google Scholar] [CrossRef]
- Borchers, A.T.; Stern, J.S.; Hackman, R.M.; Keen, C.L.; Gershwin, M.E. Medicine, Mushrooms, tumors, and immunity. Proc. Soc. Exp. Biol. Med. 1999, 221, 281–293. [Google Scholar]
- Chihara, G.; Maeda, Y.; Hamuro, J.; Sasaki, T.; Fukuoka, F. Inhibition of mouse sarcoma 180 by polysaccharides from Lentinus edodes (Berk.) sing. Nature 1969, 222, 687. [Google Scholar] [CrossRef]
- Wasser, S.P. Biotechnology, Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Guo, J.; Zhu, T.; Collins, L.; Xiao, Z.X.J.; Kim, S.H.; Chen, C.Y. Modulation of lung cancer growth arrest and apoptosis by Phellinus Linteus. Mol. Carcinog. 2007, 46, 144–154. [Google Scholar] [CrossRef]
- Collins, L.; Zhu, T.; Guo, J.; Xiao, Z.; Chen, C.Y. Phellinus linteus sensitises apoptosis induced by doxorubicin in prostate cancer. Br. J. Cancer 2006, 95, 282. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-S.; Li, G.; Kim, J.-S.; Jing, K.; Kim, T.-D.; Kim, J.-P.; Seo, S.-B.; Yoo, J.-K.; Park, H.-D.; Hwang, B.-D. Protein-bound polysaccharide from Phellinus linteus inhibits tumor growth, invasion, and angiogenesis and alters Wnt/β-catenin in SW480 human colon cancer cells. BMC Cancer 2011, 11, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Kim, D.-H.; Kim, T.-D.; Park, B.-J.; Park, H.-D.; Park, J.-I.; Na, M.-K.; Kim, H.-C.; Hong, N.-D.; Lim, K. Protein-bound polysaccharide from Phellinus linteus induces G2/M phase arrest and apoptosis in SW480 human colon cancer cells. Cancer Lett. 2004, 216, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Lee, H.-J.; Lim, E.-S.; Ahn, K.-S.; Shim, B.-S.; Kim, H.-M.; Gong, S.-J.; Kim, D.-K.; Kim, S.-H. Cambodian Phellinus linteus inhibits experimental metastasis of melanoma cells in mice via regulation of urokinase type plasminogen activator. Biol. Pharm. Bull. 2005, 28, 27–31. [Google Scholar] [CrossRef] [Green Version]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Los, M.; Mozoluk, M.; Ferrari, D.; Stepczynska, A.; Stroh, C.; Renz, A.; Herceg, Z.; Wang, Z.-Q.; Schulze-Osthoff, K. Activation and caspase-mediated inhibition of PARP: A molecular switch between fibroblast necrosis and apoptosis in death receptor signaling. Mol. Biol. Cell 2002, 13, 978–988. [Google Scholar] [CrossRef] [Green Version]
- Leber, M.F.; Efferth, T. Molecular principles of cancer invasion and metastasis. Int. J. Oncol. 2009, 34, 881–895. [Google Scholar]
- Gullett, N.P.; Amin, A.R.; Bayraktar, S.; Pezzuto, J.M.; Shin, D.M.; Khuri, F.R.; Aggarwal, B.B.; Surh, Y.-J.; Kucuk, O. Cancer prevention with natural compounds. Semin. Oncol. 2010, 37, 258–281. [Google Scholar] [CrossRef]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; Van Bree, C.J.N. Clonogenic assay of cells in vitro. Nature Protoc. 2006, 1, 2315. [Google Scholar] [CrossRef]
- Pawlik, T.M.; Keyomarsi, K. Role of cell cycle in mediating sensitivity to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 928–942. [Google Scholar] [CrossRef]
- Jackson, S.P. Sensing and repairing DNA double-strand breaks. Carcinogenesis 2002, 23, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Psaila, B.; Lyden, D. The metastatic niche: Adapting the foreign soil. Nat. Rev. Cancer 2009, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Song, H.S.; Yoo, S.H.; Yoon, M. Tumor treating fields inhibit glioblastoma cell migration, invasion and angiogenesis. Oncotarget 2016, 8, 65125. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Treatment | α (Gy-1) | β (Gy-1) |
|---|---|---|---|
| HepG2 | γ-ray | 0.249 ± 0.756 | 0.172 ± 0.219 |
| γ-ray + mesima | 0.328 ± 0.777 | 0.248 ± 0.226 | |
| Hep3B | γ-ray | 0.185 ± 0.748 | 0.124 ± 0.216 |
| γ-ray + mesima | 0.265 ± 0.767 | 0.206 ± 0.223 |
| Cell Type | Radiation | D50 without Mesima | D50 with Mesima |
|---|---|---|---|
| HepG2 | γ-ray | 1.42 Gy | 1.14 Gy |
| Hep3B | γ-ray | 1.73 Gy | 1.30 Gy |
| Cell Type | With Mesima (%) | Without Mesima (%) |
|---|---|---|
| HepG2 | 35 ± 11 | 62 ± 10 |
| Hep3B | 63 ± 3 | 82 ± 9 |
| Cell Type | Radiation | REF Value | Dose Reduction (%) |
|---|---|---|---|
| HepG2 | γ-ray | 1.24 Gy | 80.3 |
| Hep3B | γ-ray | 1.33 Gy | 75.1 |
| Vs. Control | % of TGI | Vs. Radiation | % of TGI |
|---|---|---|---|
| Mesima | 21.6 | Mesima + radiation | 28.3 |
| Radiation | 44.9 | ||
| Mesima + radiation | 60.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.K.; Oh, J.Y.; Yoo, J.K.; Lim, S.H.; Kim, E.H. The Biofunctional Effects of Mesima as a Radiosensitizer for Hepatocellular Carcinoma. Int. J. Mol. Sci. 2020, 21, 871. https://doi.org/10.3390/ijms21030871
Jeong YK, Oh JY, Yoo JK, Lim SH, Kim EH. The Biofunctional Effects of Mesima as a Radiosensitizer for Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2020; 21(3):871. https://doi.org/10.3390/ijms21030871
Chicago/Turabian StyleJeong, Youn Kyoung, Ju Yeon Oh, Jae Kuk Yoo, Sun Ha Lim, and Eun Ho Kim. 2020. "The Biofunctional Effects of Mesima as a Radiosensitizer for Hepatocellular Carcinoma" International Journal of Molecular Sciences 21, no. 3: 871. https://doi.org/10.3390/ijms21030871
APA StyleJeong, Y. K., Oh, J. Y., Yoo, J. K., Lim, S. H., & Kim, E. H. (2020). The Biofunctional Effects of Mesima as a Radiosensitizer for Hepatocellular Carcinoma. International Journal of Molecular Sciences, 21(3), 871. https://doi.org/10.3390/ijms21030871