Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
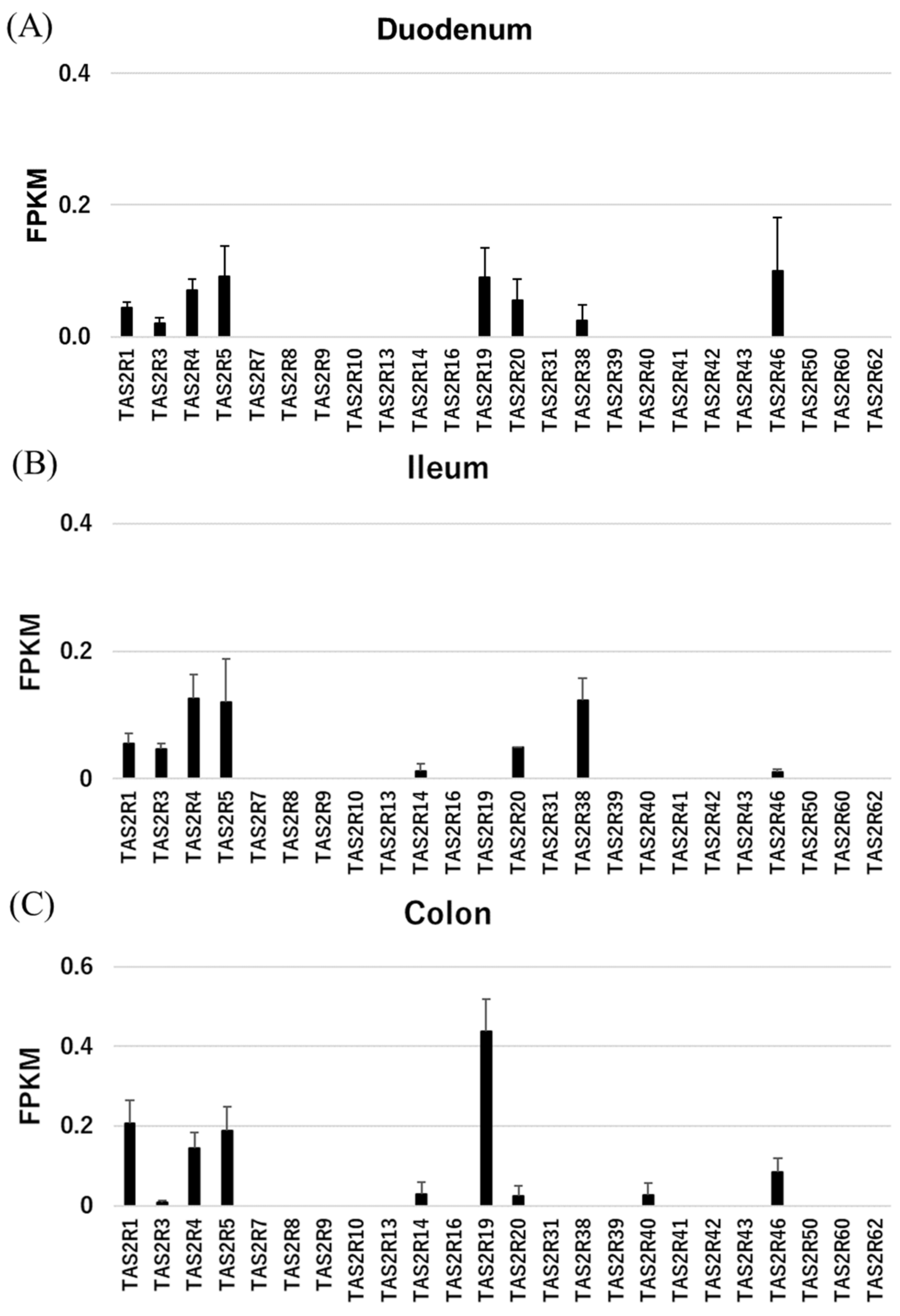
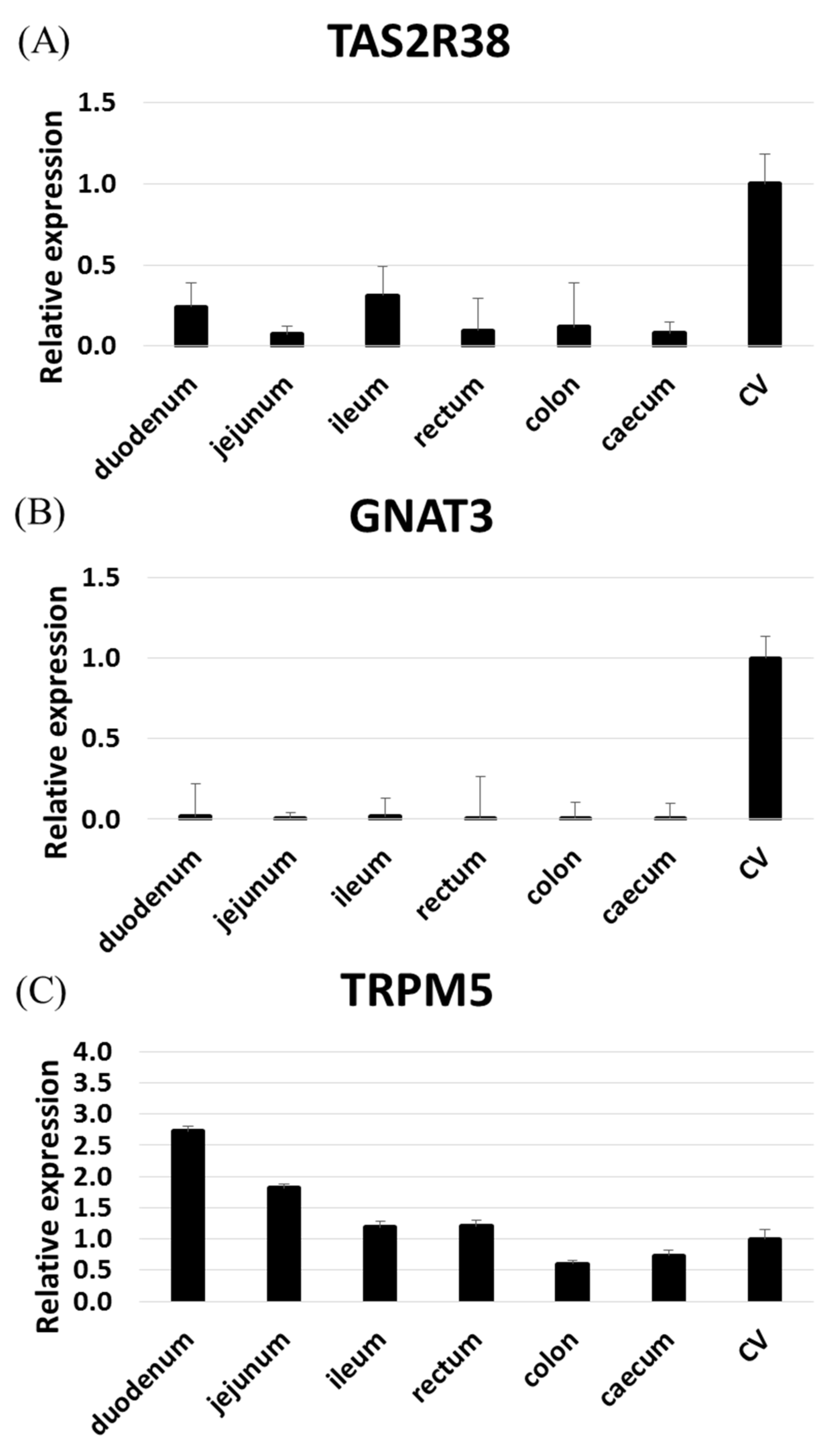
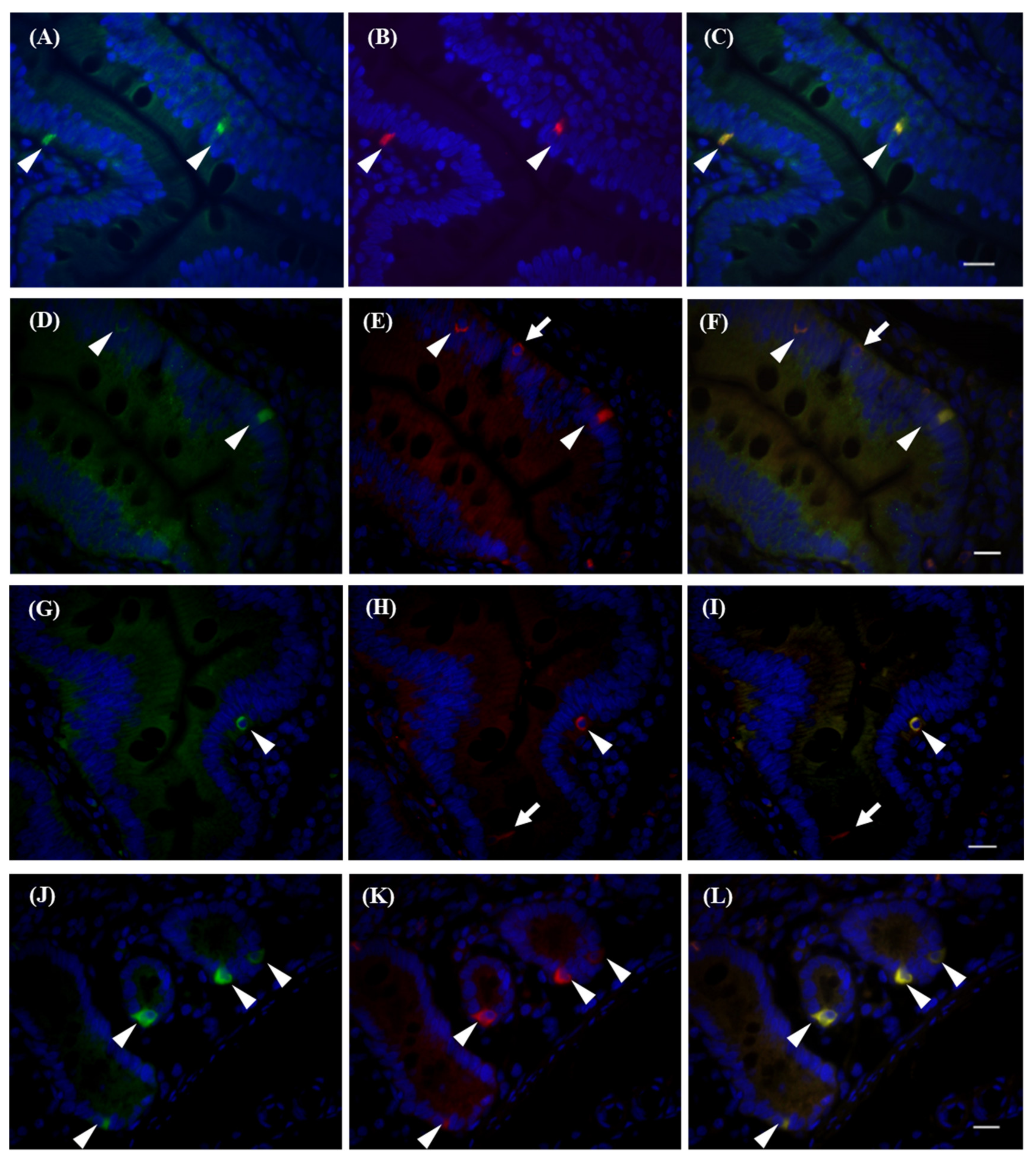
2. Results
2.1. RNAseq and RT-PCR
2.2. RNAscope
2.3. Immunohistochemistry
3. Discussion
3.1. Expression Profiles
3.2. TAS2R-Positive Cells
4. Materials and Methods
4.1. Materials
4.2. RNAseq and RT-PCR
4.3. RNAscope
4.4. Immunohistochemistry
4.5. Immunocytochemistry and Calcium Imaging
Abbreviation
| T2R | Bitter taste receptor protein |
| TAS2R | Bitter taste receptor gene in primates |
| Tas2R | Bitter taste receptor gene in rodents |
| CV | Circumvallate papillae |
| PTC | Phenylthiocarbamide |
| DAPI | 4’,6-Diamidino-2-phenylindole |
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Behrens, M.; Meyerhof, W. Gustatory and extragustatory functions of mammalian taste receptors. Physiol. Behav. 2011, 105, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Abe, M.; Asakura, T.; Imai, H.; Abe, K. Expression analysis of taste signal transduction molecules in the fungiform and circumvallate papillae of the rhesus macaque, Macaca mulatta. PloS ONE 2012, 7, e45426. [Google Scholar] [CrossRef] [PubMed]
- Gonda, S.; Matsumura, S.; Saito, S.; Go, Y.; Imai, H. Expression of taste signal transduction molecules in the caecum of common marmosets. Biol. Lett. 2013, 9, 20130409. [Google Scholar] [CrossRef] [PubMed]
- Bufe, B.; Breslin, P.A.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashido, S.N.; Hayakawa, T.; Matsui, A.; Go, Y.; Ishimaru, Y.; Misaka, T.; Abe, K.; Hirai, H.; Satta, Y.; Imai, H. Rapid expansion of phenylthiocarbamide non-tasters among Japanese macaques. PloS ONE 2015, 10, e0132016. [Google Scholar] [CrossRef] [Green Version]
- Lossow, K.; Hubner, S.; Roudnitzky, N.; Slack, J.P.; Pollastro, F.; Behrens, M.; Meyerhof, W. Comprehensive analysis of mouse bitter taste receptors reveals different molecular receptive ranges for orthologous receptors in mice and humans. J. Biol. Chem. 2016, 291, 15358–15377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prandi, S.; Voigt, A.; Meyerhof, W.; Behrens, M. Expression profiling of Tas2r genes reveals a complex pattern along the mouse GI tract and the presence of Tas2r131 in a subset of intestinal Paneth cells. Cell. Mol. Life Sci. 2018, 75, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Vegezzi, G.; Anselmi, L.; Huynh, J.; Barocelli, E.; Rozengurt, E.; Raybould, H.; Sternini, C. Diet-induced regulation of bitter taste receptor subtypes in the mouse gastrointestinal tract. PloS ONE 2014, 9, e107732. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Huynh, J.; Mazzoni, M.; Gupta, A.; Bonora, E.; Clavenzani, P.; Chang, L.; Mayer, E.A.; De Giorgio, R.; Sternini, C. Expression of the bitter taste receptor, T2R38, in enteroendocrine cells of the colonic mucosa of overweight/obese vs. lean subjects. PloS ONE 2016, 11, e0147468. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, M.; Saito, H.; Isono, K.; Goto, T.; Shirakawa, H.; Shoji, N.; Satoh-Kuriwada, S.; Sasano, T.; Okada, R.; Kudoh, K.; et al. Genotyping analysis of bitter-taste receptor genes TAS2R38 and TAS2R46 in Japanese patients with gastrointestinal cancers. J. Nutr. Sci. Vitaminol. 2017, 63, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prandi, S.; Bromke, M.; Hubner, S.; Voigt, A.; Boehm, U.; Meyerhof, W.; Behrens, M. A subset of mouse colonic goblet cells expresses the bitter taste receptor Tas2r131. PloS ONE 2013, 8, e82820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.C.; Chen, Z.H.; Xue, J.B.; Zhao, D.X.; Lu, C.; Li, Y.H.; Li, S.M.; Du, Y.W.; Liu, Q.; Wang, P.; et al. Infection by the parasitic helminth Trichinella spiralis activates a Tas2r-mediated signaling pathway in intestinal tuft cells. Proc. Natl. Acad. Sci. USA 2019, 116, 5564–5569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, T.; Kotani, T.; Setiawan, J.; Nishigaito, Y.; Sawada, N.; Imada, S.; Saito, Y.; Murata, Y.; Matozaki, T. Role of lysophosphatidic acid in proliferation and differentiation of intestinal epithelial cells. PloS ONE 2019, 14, e0215255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch, A.J.; Soucek, L.; Junttila, M.R.; Swigart, L.B.; Evan, G.I. Acute overexpression of Myc in intestinal epithelium recapitulates some but not all the changes elicited by Wnt/beta-catenin pathway activation. Mol. Cell Biol. 2009, 29, 5306–5315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, T.; Ugawa, S.; Yamamura, H.; Imaizumi, Y.; Shimada, S. Functional interaction between T2R taste receptors and G-protein alpha subunits expressed in taste receptor cells. J. Neurosci. 2003, 23, 7376–7380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggupilli, A.; Singh, N.; Upadhyaya, J.; Sikarwar, A.S.; Arakawa, M.; Dakshinamurti, S.; Bhullar, R.P.; Duan, K.; Chelikani, P. Analysis of the expression of human bitter taste receptors in extraoral tissues. Mol. Cell. Biochem. 2017, 426, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Middelhoff, M.; Westphalen, C.B.; Hayakawa, Y.; Yan, K.S.; Gershon, M.D.; Wang, T.C.; Quante, M. Dclk1-expressing tuft cells: Critical modulators of the intestinal niche? Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G285–G299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034.1–0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, M.; Born, S.; Redel, U.; Voigt, N.; Schuh, V.; Raguse, J.D.; Meyerhof, W. Immunohistochemical detection of TAS2R38 protein in human taste cells. PloS ONE. 2012, 7, e40304. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imai, H.; Hakukawa, M.; Hayashi, M.; Iwatsuki, K.; Masuda, K. Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 902. https://doi.org/10.3390/ijms21030902
Imai H, Hakukawa M, Hayashi M, Iwatsuki K, Masuda K. Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates. International Journal of Molecular Sciences. 2020; 21(3):902. https://doi.org/10.3390/ijms21030902
Chicago/Turabian StyleImai, Hiroo, Miho Hakukawa, Misa Hayashi, Ken Iwatsuki, and Katsuyoshi Masuda. 2020. "Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates" International Journal of Molecular Sciences 21, no. 3: 902. https://doi.org/10.3390/ijms21030902
APA StyleImai, H., Hakukawa, M., Hayashi, M., Iwatsuki, K., & Masuda, K. (2020). Expression of Bitter Taste Receptors in the Intestinal Cells of Non-Human Primates. International Journal of Molecular Sciences, 21(3), 902. https://doi.org/10.3390/ijms21030902