Platelet Biochemistry and Morphology after Cryopreservation


Abstract
:1. Introduction
2. Biochemical Changes to Platelets Following Cryopreservation
2.1. At the Level of Single Platelets
2.1.1. Platelet Viability and Recovery
2.1.2. Platelet Morphology
2.1.3. Changes to the Cytoplasmic Membrane
2.1.4. Surface Receptor Expression
2.1.5. Metabolic Changes
2.1.6. Signal Transduction
2.2. Platelet Function
2.2.1. Agonist-Induced Integrin Activation and Aggregation
2.2.2. Platelet Adhesion and Coagulation in Hydrodynamic Flow
2.2.3. Coagulation in Static Conditions
2.2.4. Procoagulant Cryopreserved Platelets
2.3. Clinical Relevance
2.3.1. Recovery/Count Increment and Survival Time
2.3.2. Efficacy
2.3.3. Safety
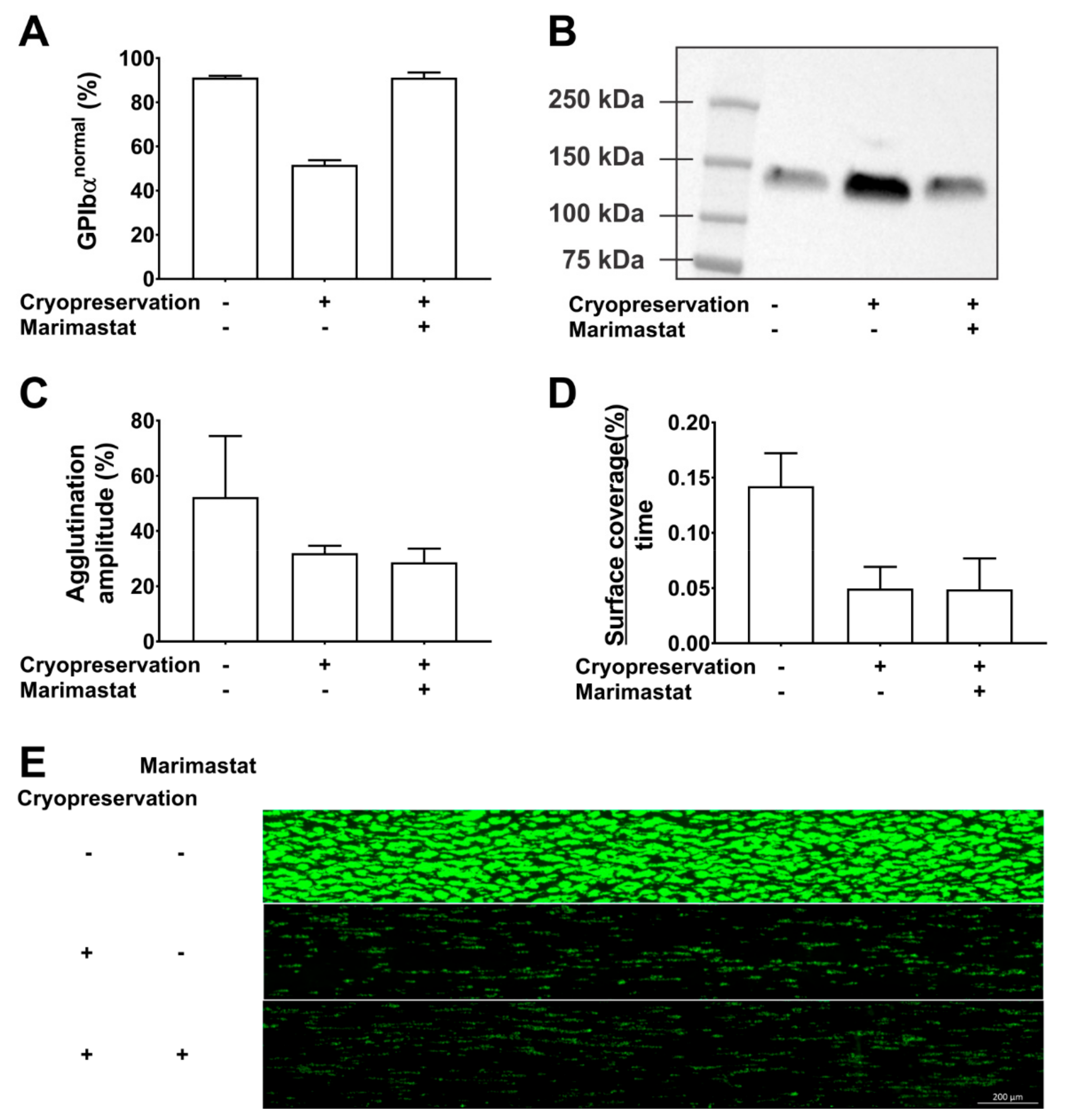
3. Changes to the GPIbα Receptor
4. Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 7-AAD | 7-aminoactinomycin D |
| ADAM17 | A Disintegrin And Metalloproteinase 17 |
| ADP | Adenosine diphosphate |
| β-GlcNAc | β-N-acetylglucosamine |
| (C)CI | (Corrected) count increment |
| CRP-XL | Collagen-related peptide, cross-linked |
| DMSO | Dimethylsulfoxide |
| ECM | Extracellular matrix |
| EV | Extracellular vesicle |
| PAR | Protease-activated receptor |
| PC | Platelet concentrate |
| PE | Phosphatidyl ethanolamine |
| PS | Phosphatidyl serine |
| ROTEM | Rotational thromboelastometry |
| RT | Room temperature |
| TF | Tissue factor |
| TGA | Thrombin generation assay |
| TRAP-6 | Thrombin receptor agonist peptide-6 |
| VWF | von Willebrand factor |
References
- Estcourt, L.J. Why has demand for platelet components increased? A review. Transfus. Med. 2014, 24, 260–268. [Google Scholar] [CrossRef]
- Murphy, S.; Gardner, F.H. Effect of storage temperature on maintenance of platelet viability--deleterious effect of refrigerated storage. N. Engl. J. Med. 1969, 280, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, M. The platelet storage lesion. Transfus. Apher. Sci. 2009, 41, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Feys, H.B.; Van Aelst, B.; Compernolle, V. Biomolecular Consequences of Platelet Pathogen Inactivation Methods. Transfus. Med. Rev. 2019, 33, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Getz, T.M. Physiology of cold-stored platelets. Transfus. Apher. Sci. 2019, 58, 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddoch-Cardenas, K.M.; Bynum, J.A.; Meledeo, M.A.; Nair, P.M.; Wu, X.; Darlington, D.N.; Ramasubramanian, A.K.; Cap, A.P. Cold-stored platelets: A product with function optimized for hemorrhage control. Transfus. Apher. Sci. 2019, 58, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, R.K.; Berndt, M.C. Platelet physiology: In cold blood. Curr Biol 2003, 13, R282–R284. [Google Scholar] [CrossRef] [Green Version]
- Valeri, C.R.; Feingold, H.; Marchionni, L.D. A simple method for freezing human platelets using 6 per cent dimethylsulfoxide and storage at −80 degrees C. Blood 1974, 43, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Cohn, C.S.; Dumont, L.J.; Lozano, M.; Marks, D.C.; Johnson, L.; Ismay, S.; Bondar, N.; T’Sas, F.; Yokoyama, A.P.H.; Kutner, J.M.; et al. Vox Sanguinis International Forum on platelet cryopreservation: Summary. Vox Sang. 2017, 112, 684–688. [Google Scholar] [CrossRef]
- Valeri, C.R.; Ragno, G.; Khuri, S. Freezing human platelets with 6 percent dimethyl sulfoxide with removal of the supernatant solution before freezing and storage at −80 degrees C without postthaw processing. Transfusion 2005, 45, 1890–1898. [Google Scholar] [CrossRef]
- Johnson, L.N.; Winter, K.M.; Reid, S.; Hartkopf-Theis, T.; Marks, D.C. Cryopreservation of buffy-coat-derived platelet concentrates in dimethyl sulfoxide and platelet additive solution. Cryobiology 2011, 62, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Meinke, S.; Wikman, A.; Gryfelt, G.; Hultenby, K.; Uhlin, M.; Hoglund, P.; Sandgren, P. Cryopreservation of buffy coat-derived platelet concentrates photochemically treated with amotosalen and UVA light. Transfusion 2018, 58, 2657–2668. [Google Scholar] [CrossRef] [PubMed]
- Balint, B.; Paunovic, D.; Vucetic, D.; Vojvodic, D.; Petakov, M.; Trkuljic, M.; Stojanovic, N. Controlled-rate versus uncontrolled-rate freezing as predictors for platelet cryopreservation efficacy. Transfusion 2006, 46, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Tan, S.; Wood, B.; Davis, A.; Marks, D.C. Refrigeration and cryopreservation of platelets differentially affect platelet metabolism and function: A comparison with conventional platelet storage conditions. Transfusion 2016, 56, 1807–1818. [Google Scholar] [CrossRef]
- Tynngard, N.; Wikman, A.; Uhlin, M.; Sandgren, P. Haemostatic responsiveness and release of biological response modifiers following cryopreservation of platelets treated with amotosalen and ultraviolet A light. Blood Transfus. 2019, 1–8. [Google Scholar]
- Johnson, L.; Coorey, C.P.; Marks, D.C. The hemostatic activity of cryopreserved platelets is mediated by phosphatidylserine-expressing platelets and platelet microparticles. Transfusion 2014, 54, 1917–1926. [Google Scholar] [CrossRef]
- Six, K.R.; Delabie, W.; Devreese, K.M.J.; Johnson, L.; Marks, D.C.; Dumont, L.J.; Compernolle, V.; Feys, H.B. Comparison between manufacturing sites shows differential adhesion, activation, and GPIbalpha expression of cryopreserved platelets. Transfusion 2018, 58, 2645–2656. [Google Scholar] [CrossRef] [Green Version]
- Dumont, L.J.; Cancelas, J.A.; Dumont, D.F.; Siegel, A.H.; Szczepiorkowski, Z.M.; Rugg, N.; Pratt, P.G.; Worsham, D.N.; Hartman, E.L.; Dunn, S.K.; et al. A randomized controlled trial evaluating recovery and survival of 6% dimethyl sulfoxide-frozen autologous platelets in healthy volunteers. Transfusion 2013, 53, 128–137. [Google Scholar] [CrossRef]
- Tegegn, T.Z.; De Paoli, S.H.; Orecna, M.; Elhelu, O.K.; Woodle, S.A.; Tarandovskiy, I.D.; Ovanesov, M.V.; Simak, J. Characterization of procoagulant extracellular vesicles and platelet membrane disintegration in DMSO-cryopreserved platelets. J. Extracell Vesicles 2016, 5, 30422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deckmyn, H.; Feys, H.B. Assays for quality control of platelets for transfusion. Isbt. Sci. Ser. 2013, 8, 221–224. [Google Scholar] [CrossRef]
- Dijkstra-Tiekstra, M.J.; de Korte, D.; Pietersz, R.N.; Reesink, H.W.; van der Meer, P.F.; Verhoeven, A.J. Comparison of various dimethylsulphoxide-containing solutions for cryopreservation of leucoreduced platelet concentrates. Vox Sang. 2003, 85, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.; Padula, M.P.; Marks, D.C.; Johnson, L. Cryopreserved platelets demonstrate reduced activation responses and impaired signaling after agonist stimulation. Transfusion 2017, 57, 2845–2857. [Google Scholar] [CrossRef] [PubMed]
- Reid, T.J.; LaRussa, V.F.; Esteban, G.; Clear, M.; Davies, L.; Shea, S.; Gorogias, M. Cooling and freezing damage platelet membrane integrity. Cryobiology 1999, 38, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.T.-B.; Burnouf, T. Preparation, quality criteria, and properties of human blood platelet lysate supplements for ex vivo stem cell expansion. New Biotechnol. 2015, 32, 199–211. [Google Scholar] [CrossRef]
- Burnouf, T.; Strunk, D.; Koh, M.B.; Schallmoser, K. Human platelet lysate: Replacing fetal bovine serum as a gold standard for human cell propagation? Biomaterials 2016, 76, 371–387. [Google Scholar] [CrossRef]
- Barnard, M.R.; MacGregor, H.; Ragno, G.; Pivacek, L.E.; Khuri, S.F.; Michelson, A.D.; Valeri, C.R. Fresh, liquid-preserved, and cryopreserved platelets: Adhesive surface receptors and membrane procoagulant activity. Transfusion 1999, 39, 880–888. [Google Scholar] [CrossRef]
- Gerber, B.; Alberio, L.; Rochat, S.; Stenner, F.; Manz, M.G.; Buser, A.; Schanz, U.; Stussi, G. Safety and efficacy of cryopreserved autologous platelet concentrates in HLA-alloimmunized patients with hematologic malignancies. Transfusion 2016, 56, 2426–2437. [Google Scholar] [CrossRef]
- Waters, L.; Padula, M.P.; Marks, D.C.; Johnson, L. Cryopreservation of UVC pathogen-inactivated platelets. Transfusion 2019, 59, 2093–2102. [Google Scholar] [CrossRef]
- Johnson, L.; Reade, M.C.; Hyland, R.A.; Tan, S.; Marks, D.C. In vitro comparison of cryopreserved and liquid platelets: Potential clinical implications. Transfusion 2015, 55, 838–847. [Google Scholar] [CrossRef]
- Kelly, K.; Dumont, L.J. Frozen platelets. Transfus. Apher. Sci. 2019, 58, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Jansen, A.J.; Josefsson, E.C.; Rumjantseva, V.; Liu, Q.P.; Falet, H.; Bergmeier, W.; Cifuni, S.M.; Sackstein, R.; von Andrian, U.H.; Wagner, D.D.; et al. Desialylation accelerates platelet clearance after refrigeration and initiates GPIbalpha metalloproteinase-mediated cleavage in mice. Blood 2012, 119, 1263–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josefsson, E.C.; Gebhard, H.H.; Stossel, T.P.; Hartwig, J.H.; Hoffmeister, K.M. The macrophage alphaMbeta2 integrin alphaM lectin domain mediates the phagocytosis of chilled platelets. J. Biol. Chem. 2005, 280, 18025–18032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, S.; Akbar, H.; Zheng, Y.; Cancelas, J.A. Towards increasing shelf life and haemostatic potency of stored platelet concentrates. Curr. Opin. Hematol. 2018, 25, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xu, B.; Chen, G.; Zhang, Y.; Wang, Q.; Zhao, L.; Zhou, H. Cryopreserved platelets augment the inflammatory response: Role of phosphatidylserine- and P-selectin-mediated platelet phagocytosis in macrophages. Transfusion 2019, 59, 1799–1808. [Google Scholar] [CrossRef]
- Born, G.V.; Cross, M.J. The aggregation of blood platelets. J. Physiol. 1963, 168, 178–195. [Google Scholar] [CrossRef]
- Keuren, J.F.; Cauwenberghs, S.; Heeremans, J.; de Kort, W.; Heemskerk, J.W.; Curvers, J. Platelet ADP response deteriorates in synthetic storage media. Transfusion 2006, 46, 204–212. [Google Scholar] [CrossRef]
- Six, K.R.; Devloo, R.; Compernolle, V.; Feys, H.B. Impact of cold storage on platelets treated with Intercept pathogen inactivation. Transfusion 2019, 59, 2662–2671. [Google Scholar] [CrossRef] [Green Version]
- Gast, A.; Tschopp, T.B.; Baumgartner, H.R. Thrombin plays a key role in late platelet thrombus growth and/or stability. Effect of a specific thrombin inhibitor on thrombogenesis induced by aortic subendothelium exposed to flowing rabbit blood. Arter. Thromb. 1994, 14, 1466–1474. [Google Scholar] [CrossRef] [Green Version]
- Six, K.R.; Devloo, R.; Van Aelst, B.; Vandekerckhove, P.; Feys, H.B.; Compernolle, V. A Microfluidic Flow Chamber Model for Platelet Transfusion and Hemostasis Measures Platelet Deposition and Fibrin Formation in Real-time. J. Vis. Exp. 2017, 120, e55351. [Google Scholar] [CrossRef] [Green Version]
- Van Aelst, B.; Feys, H.B.; Devloo, R.; Vandekerckhove, P.; Compernolle, V. Microfluidic Flow Chambers Using Reconstituted Blood to Model Hemostasis and Platelet Transfusion In Vitro. J. Vis. Exp. 2016. [Google Scholar] [CrossRef]
- Cid, J.; Escolar, G.; Galan, A.; Lopez-Vilchez, I.; Molina, P.; Diaz-Ricart, M.; Lozano, M.; Dumont, L.J. In vitro evaluation of the hemostatic effectiveness of cryopreserved platelets. Transfusion 2016, 56, 580–586. [Google Scholar] [CrossRef] [PubMed]
- The Subcommittee on Control of Anticoagulation of the, S.S.C.o.t.I. Towards a recommendation for the standardization of the measurement of platelet-dependent thrombin generation. J. Thromb. Haem. 2011, 9, 1859–1861. [Google Scholar] [CrossRef] [PubMed]
- Monroe, D.M.; Hoffman, M. What does it take to make the perfect clot? Arter. Thromb. Vasc. Biol. 2006, 26, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tomaiuolo, M.; Matzko, C.N.; Poventud-Fuentes, I.; Weisel, J.W.; Brass, L.F.; Stalker, T.J. Interrelationships between structure and function during the hemostatic response to injury. Proc. Natl. Acad. Sci. USA 2019, 116, 2243–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinauridze, E.I.; Kireev, D.A.; Popenko, N.Y.; Pichugin, A.V.; Panteleev, M.A.; Krymskaya, O.V.; Ataullakhanov, F.I. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb. Haemost. 2007, 97, 425–434. [Google Scholar]
- Slichter, S.J.; Jones, M.; Ransom, J.; Gettinger, I.; Jones, M.K.; Christoffel, T.; Pellham, E.; Bailey, S.L.; Corson, J.; Bolgiano, D. Review of in vivo studies of dimethyl sulfoxide cryopreserved platelets. Transfus. Med. Rev. 2014, 28, 212–225. [Google Scholar] [CrossRef]
- Bohonek, M.; Kutac, D.; Landova, L.; Koranova, M.; Sladkova, E.; Staskova, E.; Voldrich, M.; Tyll, T. The use of cryopreserved platelets in the treatment of polytraumatic patients and patients with massive bleeding. Transfusion 2019, 59, 1474–1478. [Google Scholar] [CrossRef] [Green Version]
- Khuri, S.F.; Healey, N.; MacGregor, H.; Barnard, M.R.; Szymanski, I.O.; Birjiniuk, V.; Michelson, A.D.; Gagnon, D.R.; Valeri, C.R. Comparison of the effects of transfusions of cryopreserved and liquid-preserved platelets on hemostasis and blood loss after cardiopulmonary bypass. J. Thorac. Cardiovasc. Surg. 1999, 117, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Quach, M.E.; Chen, W.; Li, R. Mechanisms of platelet clearance and translation to improve platelet storage. Blood 2018, 131, 1512–1521. [Google Scholar] [CrossRef]
- Cohn, C.S.; Williams, S. Cryopreserved platelets: The thaw begins. Transfusion 2019, 59, 2759–2762. [Google Scholar] [CrossRef] [Green Version]
- Cancelas, J.A. Future of platelet formulations with improved clotting profile: A short review on human safety and efficacy data. Transfusion 2019, 59, 1467–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milford, E.M.; Reade, M.C. Comprehensive review of platelet storage methods for use in the treatment of active hemorrhage. Transfusion 2016, 56, S140–S148. [Google Scholar] [CrossRef] [Green Version]
- Lelkens, C.C.; Koning, J.G.; de Kort, B.; Floot, I.B.; Noorman, F. Experiences with frozen blood products in the Netherlands military. Transfus. Apher. Sci. 2006, 34, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Noorman, F.; van Dongen, T.T.; Plat, M.J.; Badloe, J.F.; Hess, J.R.; Hoencamp, R. Transfusion: −80 degrees C Frozen Blood Products Are Safe and Effective in Military Casualty Care. PLoS ONE 2016, 11, e0168401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyatt, G.H.; Oxman, A.D.; Vist, G.E.; Kunz, R.; Falck-Ytter, Y.; Alonso-Coello, P.; Schünemann, H.J. GRADE: An emerging consensus on rating quality of evidence and strength of recommendations. BMJ 2008, 336, 924–926. [Google Scholar] [CrossRef] [Green Version]
- Reade, M.C.; Marks, D.C.; Bellomo, R.; Deans, R.; Faulke, D.J.; Fraser, J.F.; Gattas, D.J.; Holley, A.D.; Irving, D.O.; Johnson, L.; et al. A randomized, controlled pilot clinical trial of cryopreserved platelets for perioperative surgical bleeding: The CLIP-I trial. Transfusion 2019, 59, 2794–2804. [Google Scholar] [CrossRef]
- Group, T.t.R.A.t.P.S. Leukocyte reduction and ultraviolet B irradiation of platelets to prevent alloimmunization and refractoriness to platelet transfusions. N. Engl. J. Med. 1997, 337, 1861–1869. [Google Scholar]
- Slichter, S.J.; Dumont, L.J.; Cancelas, J.A.; Jones, M.; Gernsheimer, T.B.; Szczepiorkowski, Z.M.; Dunbar, N.M.; Prakash, G.; Medlin, S.; Rugg, N.; et al. Safety and efficacy of cryopreserved platelets in bleeding patients with thrombocytopenia. Transfusion 2018, 58, 2129–2138. [Google Scholar] [CrossRef] [Green Version]
- Bergmeier, W.; Piffath, C.L.; Cheng, G.; Dole, V.S.; Zhang, Y.; von Andrian, U.H.; Wagner, D.D. Tumor necrosis factor-alpha-converting enzyme (ADAM17) mediates GPIbalpha shedding from platelets in vitro and in vivo. Circ. Res. 2004, 95, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Six, K.R.; Lyssens, S.; Devloo, R.; Compernolle, V.; Feys, H.B. The ice recrystallization inhibitor polyvinyl alcohol does not improve platelet cryopreservation. Transfusion 2019, 59, 3029–3031. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| RT Stored Platelets | Cryopreserved Platelets | |
|---|---|---|
| General | ||
| Storage conditions | 22 °C with constant agitation | −80 °C without agitation |
| Shelf-life | 5–7 days | At least 2 years |
| Platelet Characteristics | ||
| Morphology | Disc | Sphere |
| Cytoplasmic membrane | Membrane asymmetry | PS/PE expression Loss of membrane integrity |
| Degranulation | Minimal | Increased |
| Metabolism | Normal | Loss of mitochondrial membrane potential Decreased hypotonic shock response Accelerated glycolysis |
| GPIbα shedding | Minimal | Increased |
| EV formation | Minimal | Increased |
| Platelet Function | ||
| PAC-1 binding upon stimulation with agonists | Normal | Decreased |
| Aggregation response upon stimulation with agonists | Normal | Decreased |
| Adhesion rate to ECM under flow | Normal | Decreased |
| Coagulation rate under flow | Normal | Increased |
| ROTEM | ||
| Clotting time | Normal | Shortened |
| Clot firmness | Normal | Slightly decreased |
| TGA | ||
| Peak thrombin | Normal | Increased |
| Lag time | Normal | Shortened |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Six, K.R.; Compernolle, V.; Feys, H.B. Platelet Biochemistry and Morphology after Cryopreservation. Int. J. Mol. Sci. 2020, 21, 935. https://doi.org/10.3390/ijms21030935
Six KR, Compernolle V, Feys HB. Platelet Biochemistry and Morphology after Cryopreservation. International Journal of Molecular Sciences. 2020; 21(3):935. https://doi.org/10.3390/ijms21030935
Chicago/Turabian StyleSix, Katrijn R., Veerle Compernolle, and Hendrik B. Feys. 2020. "Platelet Biochemistry and Morphology after Cryopreservation" International Journal of Molecular Sciences 21, no. 3: 935. https://doi.org/10.3390/ijms21030935
APA StyleSix, K. R., Compernolle, V., & Feys, H. B. (2020). Platelet Biochemistry and Morphology after Cryopreservation. International Journal of Molecular Sciences, 21(3), 935. https://doi.org/10.3390/ijms21030935