AAV-Mediated Gene Delivery to 3D Retinal Organoids Derived from Human Induced Pluripotent Stem Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
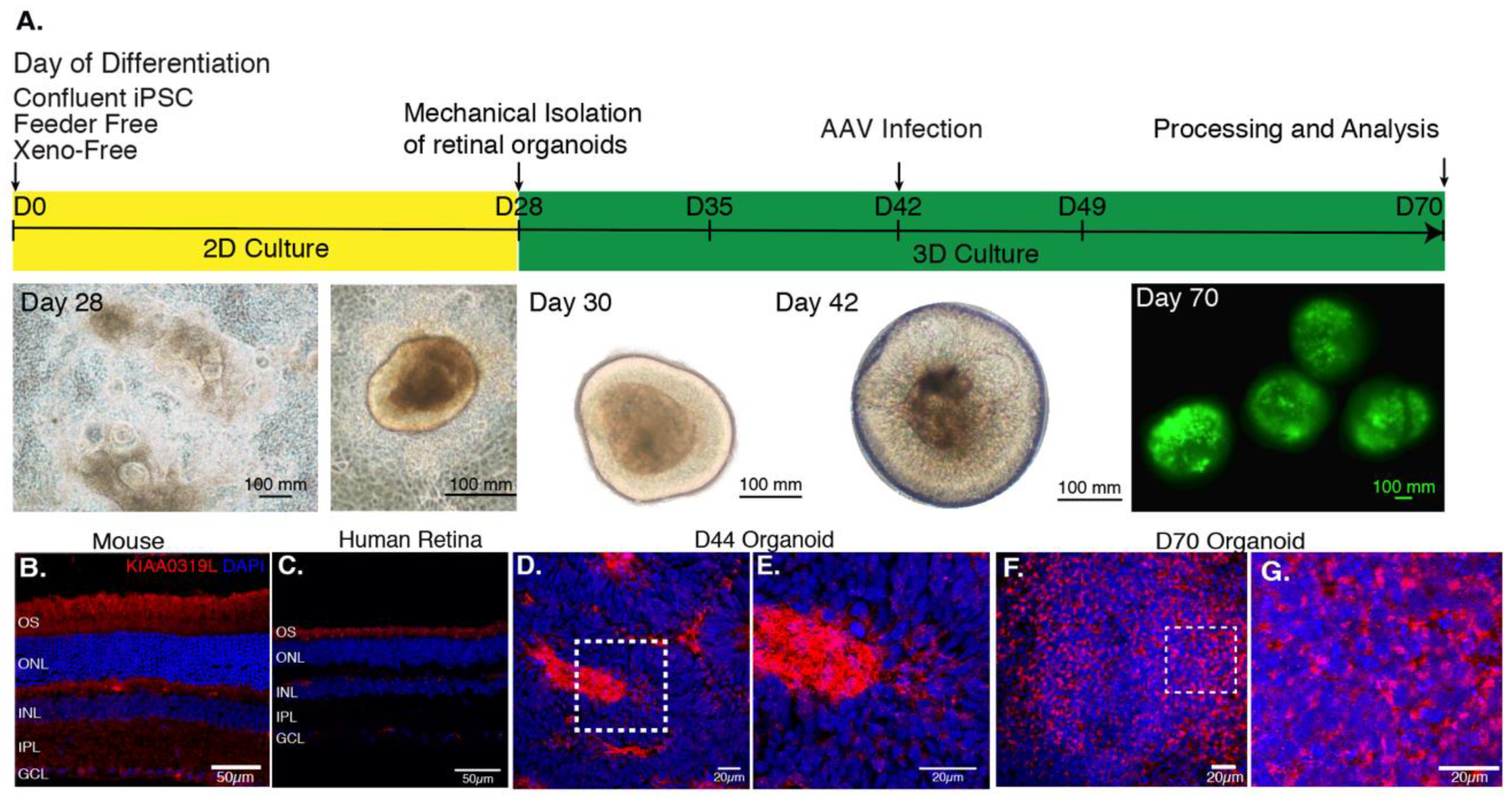
2.1. Presence of the Universal AAV Receptor AAVR in 3D Retinal Organoids
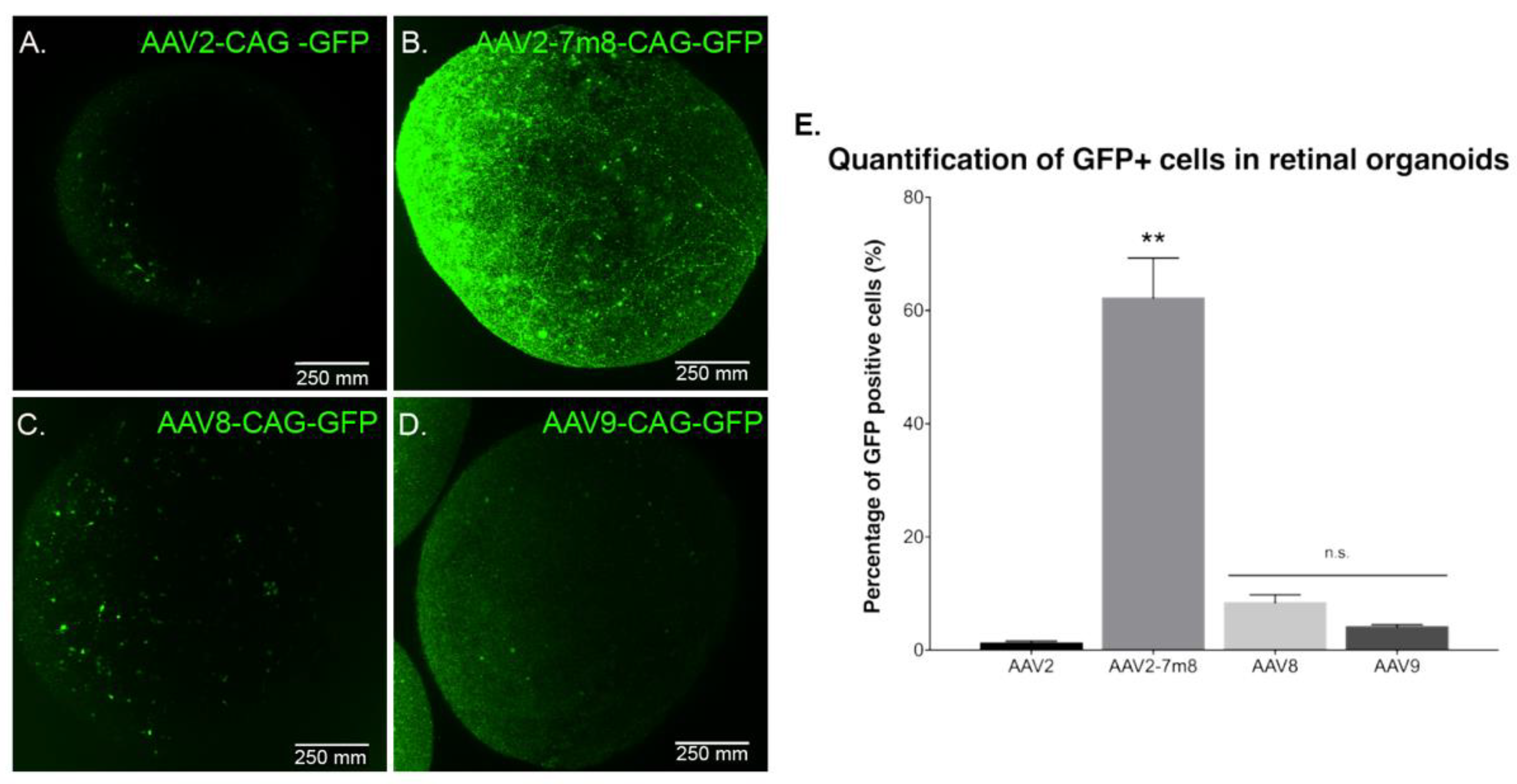
2.2. Tropism of Different rAAVs towards Human Retinal Organoids
2.3. AAV Transduction Efficiency Correlates with the Presence of Cell-Surface Receptors
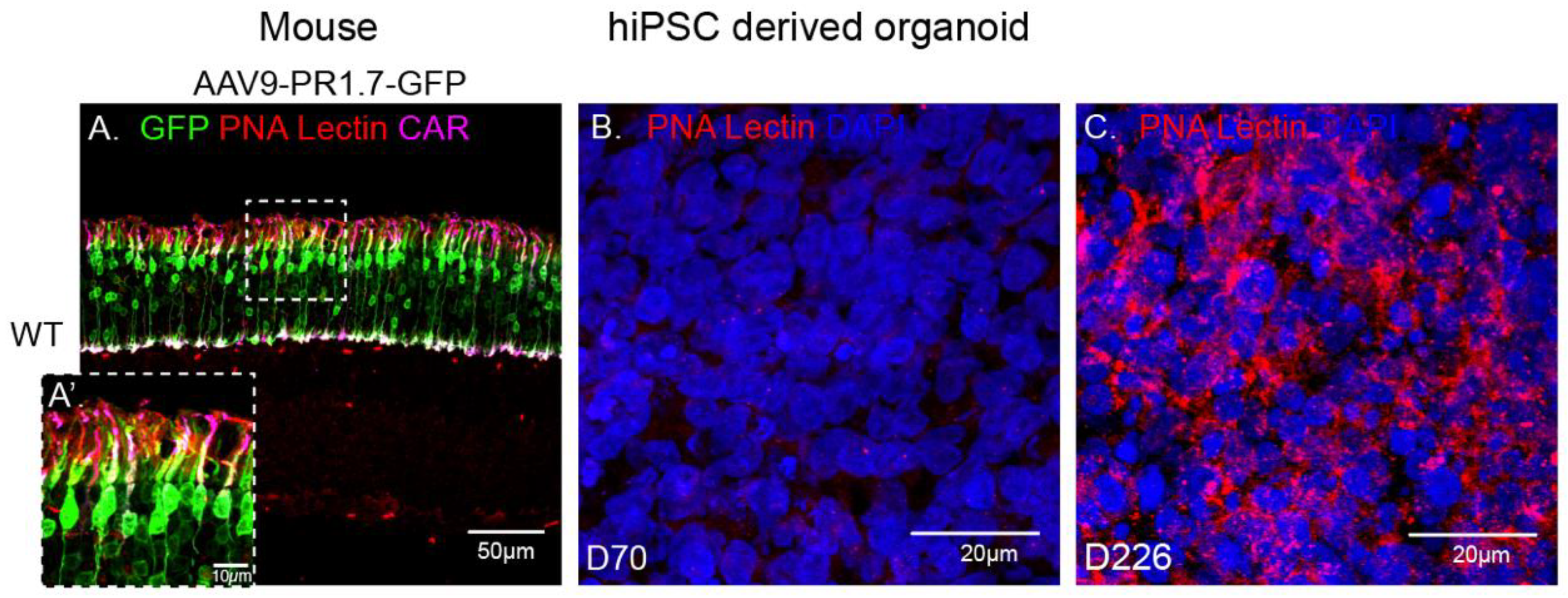
2.4. rAAV9 and Cone Transduction Efficiency
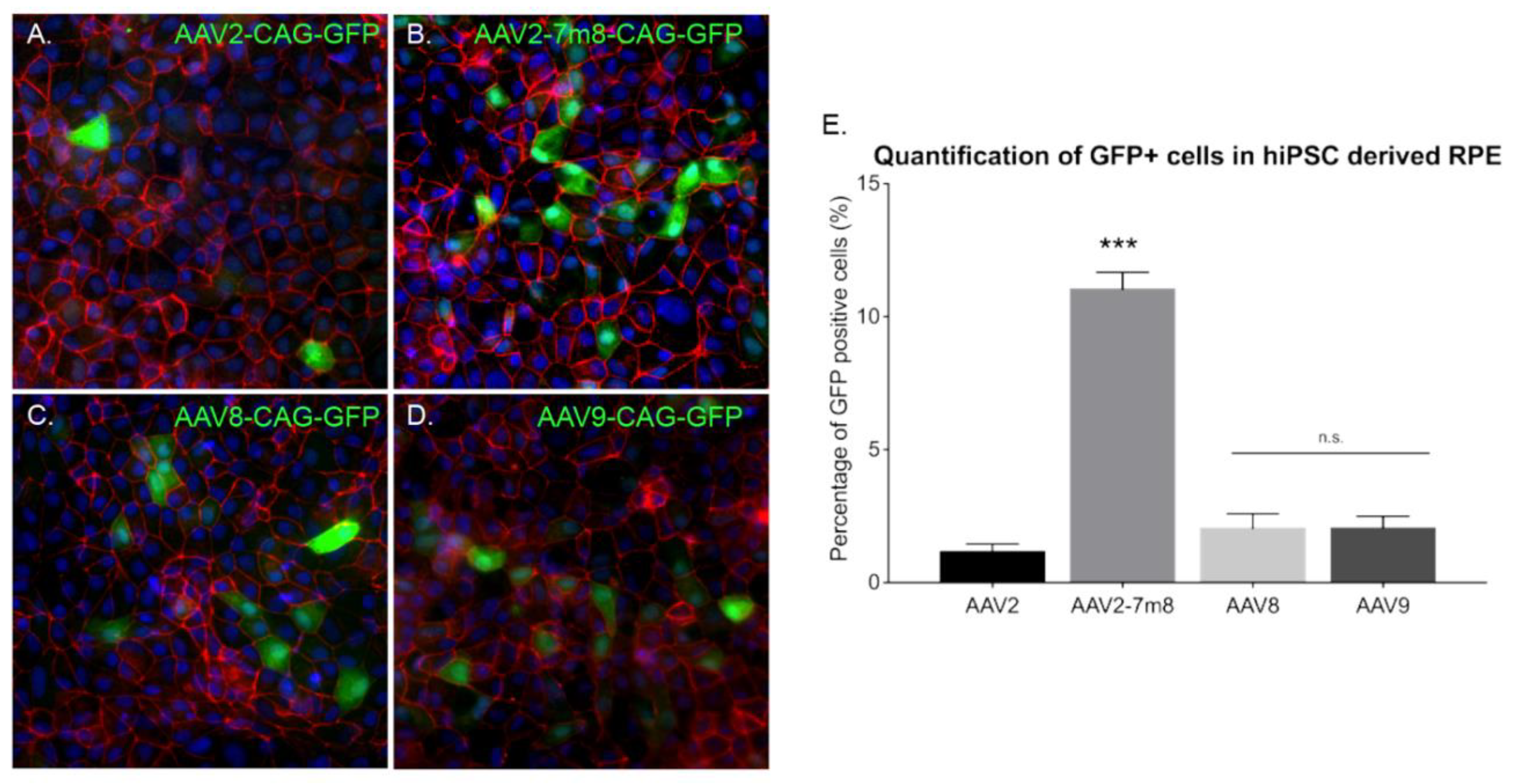
2.5. AAV Mediated Transduction of hiPSC-Derived RPE
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Maintenance of hiPSC Culture
4.3. Differentiation of Human iPS Cells into Retinal Organoids
4.4. Generation of RPE from Human iPS Cells (hi-RPE Cells)
4.5. AAV Vector Production
4.6. Infection of Retinal Organoids with AAV Capsids
4.7. Infection of hiPSC-Derived RPE with AAV Capsids
4.8. Subretinal Injections
4.9. Tissue Preparation
4.10. Cryosectionning and Immunohistochemistry
4.11. Image Acquisition
4.12. RNA Isolation and RT-PCR
4.13. Cell Counts on RPE Cells
4.14. FACS Analysis
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAV | Adeno-Associated Virus |
| AAVR | Adeno-Associated Virus Receptor |
| CAG | CMV early enhancer/chicken β actin |
| ESC | Embryonic Stem Cells |
| FGFR1 | Fibroblast Growth Factor Receptors 1 |
| GFP | Green Fluorescent Protein |
| hiPSC | Human induced Pluripotent Stem Cells |
| ILM | Inner Limiting Membrane |
| PDGFR | Platelet-Derived Growth Factor Receptors |
| PNA | Peanut Agglutinin |
| PR | Photoreceptors |
| RT | Room Temperature |
| RT-PCR | Reverse Transcription Polymerase Chain Reaction |
| RPE | Retinal Pigmented Epithelium |
References
- Meyer, J.S.; Shearer, R.L.; Capowski, E.E.; Wright, L.S.; Wallace, K.A.; McMillan, E.L.; Zhang, S.-C.C.; Gamm, D.M. Modeling early retinal development with human embryonic and induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2009, 106, 16698–16703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Ando, S.; Takata, N.; Kawada, M.; Muguruma, K.; Sekiguchi, K.; Saito, K.; Yonemura, S.; Eiraku, M.; Sasai, Y. Self-formation of optic cups and storable stratified neural retina from human ESCs. Cell Stem Cell 2012, 10, 771–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichman, S.; Terray, A.; Slembrouck, A.; Nanteau, C.; Orieux, G.; Habeler, W.; Nandrot, E.F.; Sahel, J.-A.J.-A.J.-A.; Monville, C.; Goureau, O. From confluent human iPS cells to self-forming neural retina and retinal pigmented epithelium. Proc. Natl. Acad. Sci. USA 2014, 111, 8518–8523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Gutierrez, C.; Xue, T.; Hampton, C.; Natalia, M.; Cao, L.-H.; Peters, A.; Park, T.S.; Zambidis, E.T.; Meyer, J.S.; et al. Generation of three-dimensional retinal tissue with functional photoreceptors from human iPSCs. Nat. Commun. 2014, 5, 4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vugler, A.; Carr, A.J.; Lawrence, J.; Chen, L.L.; Burrell, K.; Wright, A.; Lundh, P.; Semo, M.; Ahmado, A.; Gias, C.; et al. Elucidating the phenomenon of HESC-derived RPE: anatomy of cell genesis, expansion and retinal transplantation. Exp. Neurol. 2008, 214, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.J.; Vugler, A.A.; Hikita, S.T.; Lawrence, J.M.; Gias, C.; Chen, L.L.; Buchholz, D.E.; Ahmado, A.; Semo, M.; Smart, M.J.; et al. Protective effects of human iPS-derived retinal pigment epithelium cell transplantation in the retinal dystrophic rat. PLoS ONE 2009, 4, e8152. [Google Scholar] [CrossRef]
- M’Barek, K.B.; Habeler, W.; Plancheron, A.; Jarraya, M.; Regent, F.; Terray, A.; Yang, Y.; Chatrousse, L.; Domingues, S.; Masson, Y.; et al. Human ESC–derived retinal epithelial cell sheets potentiate rescue of photoreceptor cell loss in rats with retinal degeneration. Sci. Transl. Med. 2017, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Planul, A.; Dalkara, D. Vectors and Gene Delivery to the Retina. Annu. Rev. Vis. Sci. 2017, 3, 121–140. [Google Scholar] [CrossRef]
- Hastie, E.; Samulski, R.J. AAV at 50: A golden anniversary of discovery, research, and gene therapy success, a personal perspective. Hum. Gene Ther. 2015, 26, A7. [Google Scholar] [CrossRef] [Green Version]
- Cearley, C.N.; Wolfe, J.H. Transduction characteristics of adeno-associated virus vectors expressing cap serotypes 7, 8, 9, and Rh10 in the mouse brain. Mol. Ther. 2006, 13, 528–537. [Google Scholar] [CrossRef]
- Trapani, I.; Puppo, A.; Auricchio, A. Vector platforms for gene therapy of inherited retinopathies. Prog. Retin. Eye Res. 2014, 43, 108–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garita-Hernandez, M.; Guibbal, L.; Toualbi, L.; Routet, F.; Chaffiol, A.; Winckler, C.; Harinquet, M.; Robert, C.; Fouquet, S.; Bellow, S.; et al. Optogenetic Light Sensors in Human Retinal Organoids. Front. Neurosci. 2018, 12, 789. [Google Scholar] [CrossRef] [PubMed]
- Garita-Hernandez, M.; Lampič, M.; Chaffiol, A.; Guibbal, L.; Routet, F.; Santos-Ferreira, T.; Gasparini, S.; Borsch, O.; Gagliardi, G.; Reichman, S.; et al. Restoration of visual function by transplantation of optogenetically engineered photoreceptors. Nat. Commun. 2019, 10, 4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberghe, L.H.; Bell, P.; Maguire, A.M.; Cearley, C.N.; Xiao, R.; Calcedo, R.; Wang, L.; Castle, M.J.; Maguire, A.C.; Grant, R.; et al. Dosage thresholds for AAV2 and AAV8 photoreceptor gene therapy in monkey. Sci. Transl. Med. 2011, 3, 88ra54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, G.-J.; Budzynski, E.; Sonnentag, P.; Nork, T.M.; Sheibani, N.; Gurel, Z.; Boye, S.L.; Peterson, J.J.; Boye, S.E.; Hauswirth, W.W.; et al. Cone-Specific Promoters for Gene Therapy of Achromatopsia and Other Retinal Diseases. Hum. Gene Ther. 2016, 27, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Dalkara, D.; Byrne, L.C.; Klimczak, R.R.; Visel, M.; Yin, L.; Merigan, W.H.; Flannery, J.G.; Schaffer, D.V. In vivo-directed evolution of a new adeno-associated virus for therapeutic outer retinal gene delivery from the vitreous. Sci. Transl. Med. 2013, 5, 189ra76. [Google Scholar] [CrossRef] [PubMed]
- Khabou, H.; Desrosiers, M.; Winckler, C.; Fouquet, S.; Auregan, G.; Bemelmans, A.-P.; Sahel, J.-A.; Dalkara, D. Insight into the mechanisms of enhanced retinal transduction by the engineered AAV2 capsid variant -7m8. Biotechnol. Bioeng. 2016, 113, 2712–2724. [Google Scholar] [CrossRef] [Green Version]
- Welby, E.; Lakowski, J.; Di Foggia, V.; Budinger, D.; Gonzalez-Cordero, A.; Lun, A.T.L.; Epstein, M.; Patel, A.; Cuevas, E.; Kruczek, K.; et al. Isolation and Comparative Transcriptome Analysis of Human Fetal and iPSC-Derived Cone Photoreceptor Cells. Stem Cell Rep. 2017, 9, 1898–1915. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Cordero, A.; Goh, D.; Kruczek, K.; Naeem, A.; Fernando, M.; Holthaus, S.-M.K.; Takaaki, M.; Blackford, S.J.I.; Kloc, M.; Agundez, L.; et al. Assessment of AAV Vector Tropisms for Mouse and Human Pluripotent Stem Cell–Derived RPE and Photoreceptor Cells. Hum. Gene Ther. 2018, 29, 1124–1139. [Google Scholar] [CrossRef]
- Quinn, P.M.; Buck, T.M.; Ohonin, C.; Mikkers, H.M.M.; Wijnholds, J. Production of iPS-Derived Human Retinal Organoids for Use in Transgene Expression Assays; Humana Press: New York, NY, USA, 2018; pp. 261–273. [Google Scholar]
- Duong, T.T.; Lim, J.; Vasireddy, V.; Papp, T.; Nguyen, H.; Leo, L.; Pan, J.; Zhou, S.; Chen, H.I.; Bennett, J.; et al. Comparative AAV-eGFP Transgene Expression Using Vector Serotypes 1–9, 7m8, and 8b in Human Pluripotent Stem Cells, RPEs, and Human and Rat Cortical Neurons. Stem Cells Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Pillay, S.; Meyer, N.L.; Puschnik, A.S.; Davulcu, O.; Diep, J.; Ishikawa, Y.; Jae, L.T.; Wosen, J.E.; Nagamine, C.M.; Chapman, M.S.; et al. An essential receptor for adeno-associated virus infection. Nature 2016, 530, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Auricchio, A.; Kobinger, G.; Anand, V.; Hildinger, M.; O’Connor, E.; Maguire, A.M.; Wilson, J.M.; Bennett, J. Exchange of surface proteins impacts on viral vector cellular specificity and transduction characteristics: the retina as a model. Hum. Mol. Genet. 2001, 10, 3075–3081. [Google Scholar] [CrossRef] [Green Version]
- Natkunarajah, M.; Trittibach, P.; Mcintosh, J.; Duran, Y.; Barker, S.E.; Smith, A.J.; Nathwani, A.C.; Ali, R.R. Assessment of ocular transduction using single-stranded and self-complementary recombinant adeno-associated virus serotype 2/8. Gene Ther. 2008, 15, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allocca, M.; Mussolino, C.; Garcia-Hoyos, M.; Sanges, D.; Iodice, C.; Petrillo, M.; Vandenberghe, L.H.; Wilson, J.M.; Marigo, V.; Surace, E.M.; et al. Novel adeno-associated virus serotypes efficiently transduce murine photoreceptors. J. Virol. 2007, 81, 11372–11380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernet, V.; Joly, S.; Jordi, N.; Dalkara, D.; Guzik-Kornacka, A.; Flannery, J.G.; Schwab, M.E. Misguidance and modulation of axonal regeneration by Stat3 and Rho/ROCK signaling in the transparent optic nerve. Cell Death Dis. 2013, 4, e734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichman, S.; Slembrouck, A.; Gagliardi, G.; Chaffiol, A.; Terray, A.; Nanteau, C.; Potey, A.; Belle, M.; Rabesandratana, O.; Duebel, J.; et al. Generation of Storable Retinal Organoids and Retinal Pigmented Epithelium from Adherent Human iPS Cells in Xeno-Free and Feeder-Free Conditions. Stem Cells 2017, 35, 1176–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inatani, M.; Honjo, M.; Oohira, A.; Kido, N.; Otori, Y.; Tano, Y.; Honda, Y.; Tanihara, H. a Transmembrane Heparan Sulfate Proteoglycan, in Developing Retina. Invest. Ophthalmol. 2002, 43, 1616–1621. [Google Scholar]
- De Luca, M.; Klimentidis, Y.C.; Casazza, K.; Chambers, M.M.; Cho, R.; Harbison, S.T.; Jumbo-Lucioni, P.; Zhang, S.; Leips, J.; Fernandez, J.R. A conserved role for syndecan family members in the regulation of whole-body energy metabolism. PLoS ONE 2010, 5, e11286. [Google Scholar] [CrossRef]
- Vandenberghe, L.H.; Bell, P.; Maguire, A.M.; Xiao, R.; Hopkins, T.B.; Grant, R.; Bennett, J.; Wilson, J.M. AAV9 Targets Cone Photoreceptors in the Nonhuman Primate Retina. PLoS ONE 2013, 8, e53463. [Google Scholar] [CrossRef]
- Khabou, H.; Garita-Hernandez, M.; Chaffiol, A.; Reichman, S.; Jaillard, C.; Brazhnikova, E.; Bertin, S.; Forster, V.; Desrosiers, M.; Winckler, C.; et al. Noninvasive gene delivery to foveal cones for vision restoration. JCI Insight 2018, 3, e96029. [Google Scholar] [CrossRef] [Green Version]
- Garita-Hernandez, M.; Goureau, O.; Dalkara, D. Gene and Cell Therapy for Inherited Retinal Dystrophies. eLS 2001, 1–16. [Google Scholar] [CrossRef]
- Watanabe, S.; Sanuki, R.; Ueno, S.; Koyasu, T.; Hasegawa, T.; Furukawa, T. Tropisms of AAV for Subretinal Delivery to the Neonatal Mouse Retina and Its Application for In Vivo Rescue of Developmental Photoreceptor Disorders. PLoS ONE 2013, 8, e54146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, L.; Ma, S.; Cheng, S.Y.; Gao, G.; Punzo, C. Rod Outer Segment Development Influences AAV-Mediated Photoreceptor Transduction after Subretinal Injection. Hum. Gene Ther. 2017, 28, 464–481. [Google Scholar] [CrossRef] [PubMed]
- Dudek, A.M.; Pillay, S.; Puschnik, A.S.; Nagamine, C.M.; Cheng, F.; Qiu, J.; Carette, J.E.; Vandenberghe, L.H. An alternate route for adeno-associated virus entry independent of AAVR. J. Virol. 2018, 92, e02213-17. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proneural Medium | Essential 6 Medium (A1516401) N-2 supplement (100X) 1% (17502048) Penicillin-Streptomycin 1% (15140122) |
| Maturation Medium | DMEM/F-12 (11320074) B27 supplement (100X), 2% (17504044) MEM Non-Essential Amino Acid Solution (100X), 1% (11140035) Penicillin-Streptomycin 1% (15140122) |
| RPE Medium | DMEM/F-12 (11320074) N-2 supplement (100X) 1% (17502048) Penicillin-Streptomycin 1% (15140122) |
| Antigen | Manufacturer/Reference | Specie/Clonality | Dilution |
|---|---|---|---|
| GFP | Abcam ab13970 | Chicken polyclonal | 1/500 |
| Syndecan3 | Antibodies-online ABIN4352331 | Rabbit polyclonal | 1/200 |
| RPSA (Laminin Receptor 1) | Abcam ab137388 | Rabbit polyclonal | 1/500 |
| PNA Rhodamine conjugated | Vector Laboratories RL-1072 | Lectin | 1/100 |
| KIAA0319L (AAVR) | Abcam ab105385 | Mouse polyclonal | 1/200 |
| ZO-1 | Invitrogen 61-7300 | Rabbit polyclonal | 1/2000 |
| MITF | DAKO M3621 | Mouse monoclonal | 1/100 |
| FAK | Millipore 05-185 | Mouse monoclonal | 1/100 |
| Cone Arrestin | Millipore MAB15282 | Rabbit polyclonal | 1/2000 |
| CRX | Abnova H00001406-M02 | Mouse monoclonal | 1/5000 |
| FGFR1 | Forward: CCAGACAACCTGCCTTATGT Reverse: GCTGTGGAAGTCACTCTTCTT Amplicon size 327 bp |
| RPSA (Laminin Receptor 1) | Forward: CACTCCTGGAACCTTCACTAAC Reverse: CTCCAGTCTTCAGTAGGGAATTG Amplicon size 495 bp |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garita-Hernandez, M.; Routet, F.; Guibbal, L.; Khabou, H.; Toualbi, L.; Riancho, L.; Reichman, S.; Duebel, J.; Sahel, J.-A.; Goureau, O.; et al. AAV-Mediated Gene Delivery to 3D Retinal Organoids Derived from Human Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2020, 21, 994. https://doi.org/10.3390/ijms21030994
Garita-Hernandez M, Routet F, Guibbal L, Khabou H, Toualbi L, Riancho L, Reichman S, Duebel J, Sahel J-A, Goureau O, et al. AAV-Mediated Gene Delivery to 3D Retinal Organoids Derived from Human Induced Pluripotent Stem Cells. International Journal of Molecular Sciences. 2020; 21(3):994. https://doi.org/10.3390/ijms21030994
Chicago/Turabian StyleGarita-Hernandez, Marcela, Fiona Routet, Laure Guibbal, Hanen Khabou, Lyes Toualbi, Luisa Riancho, Sacha Reichman, Jens Duebel, Jose-Alain Sahel, Olivier Goureau, and et al. 2020. "AAV-Mediated Gene Delivery to 3D Retinal Organoids Derived from Human Induced Pluripotent Stem Cells" International Journal of Molecular Sciences 21, no. 3: 994. https://doi.org/10.3390/ijms21030994
APA StyleGarita-Hernandez, M., Routet, F., Guibbal, L., Khabou, H., Toualbi, L., Riancho, L., Reichman, S., Duebel, J., Sahel, J. -A., Goureau, O., & Dalkara, D. (2020). AAV-Mediated Gene Delivery to 3D Retinal Organoids Derived from Human Induced Pluripotent Stem Cells. International Journal of Molecular Sciences, 21(3), 994. https://doi.org/10.3390/ijms21030994