Involvement of Satellite Cell Activation via Nitric Oxide Signaling in Ectopic Orofacial Hypersensitivity


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
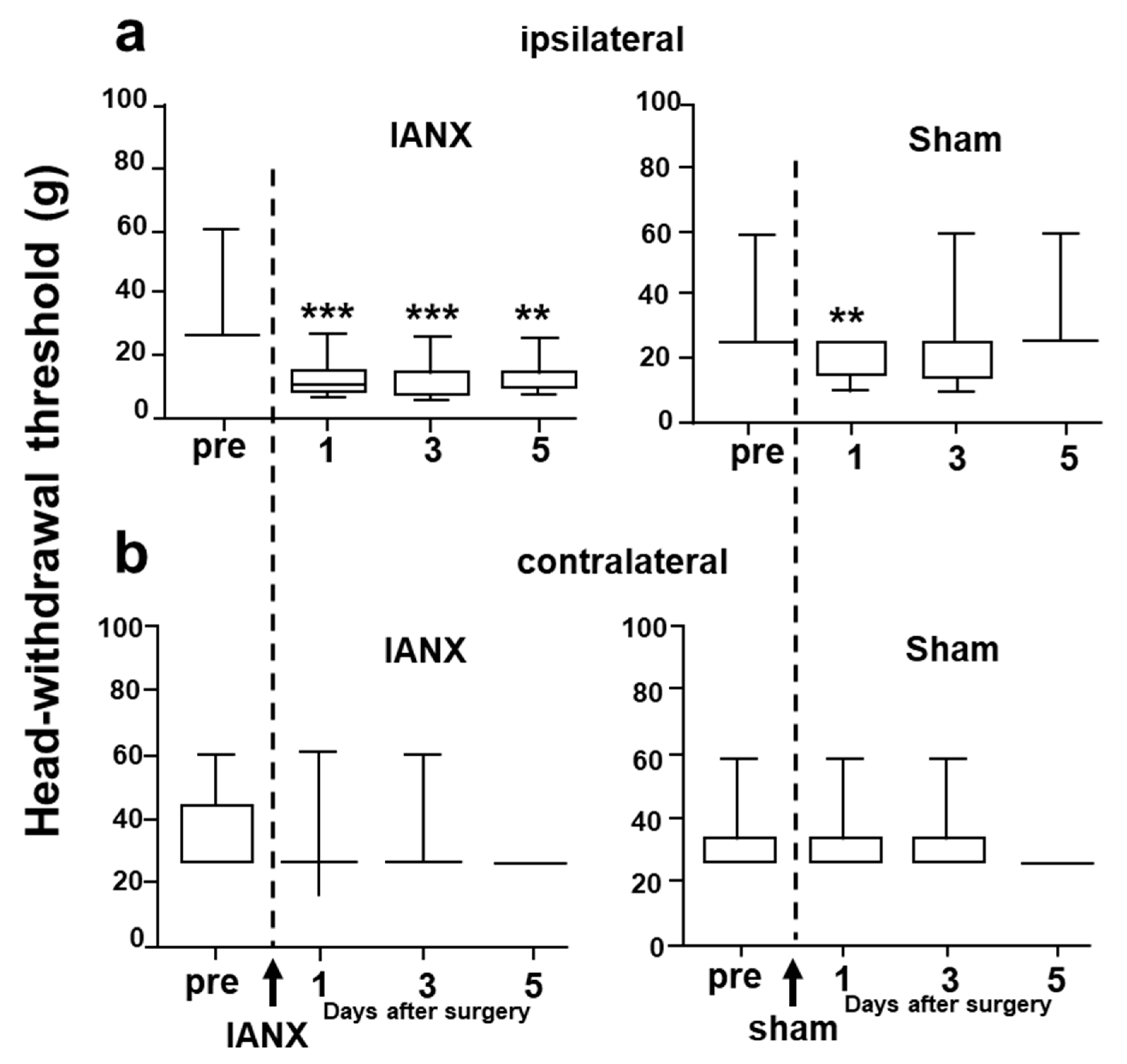
2.1. Mechanical Head-Withdrawal Threshold
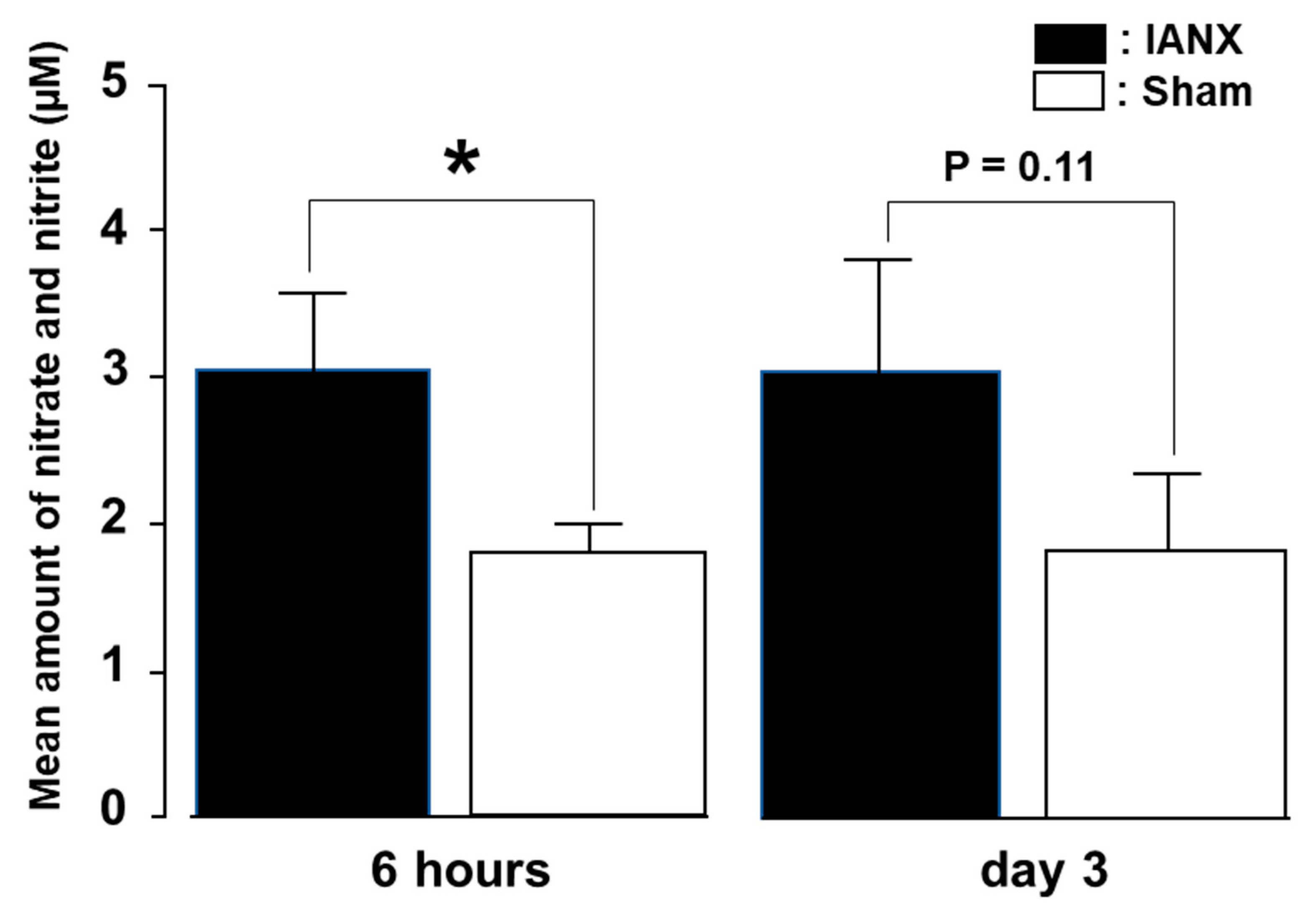
2.2. Nitrate–Nitrite Synthesis
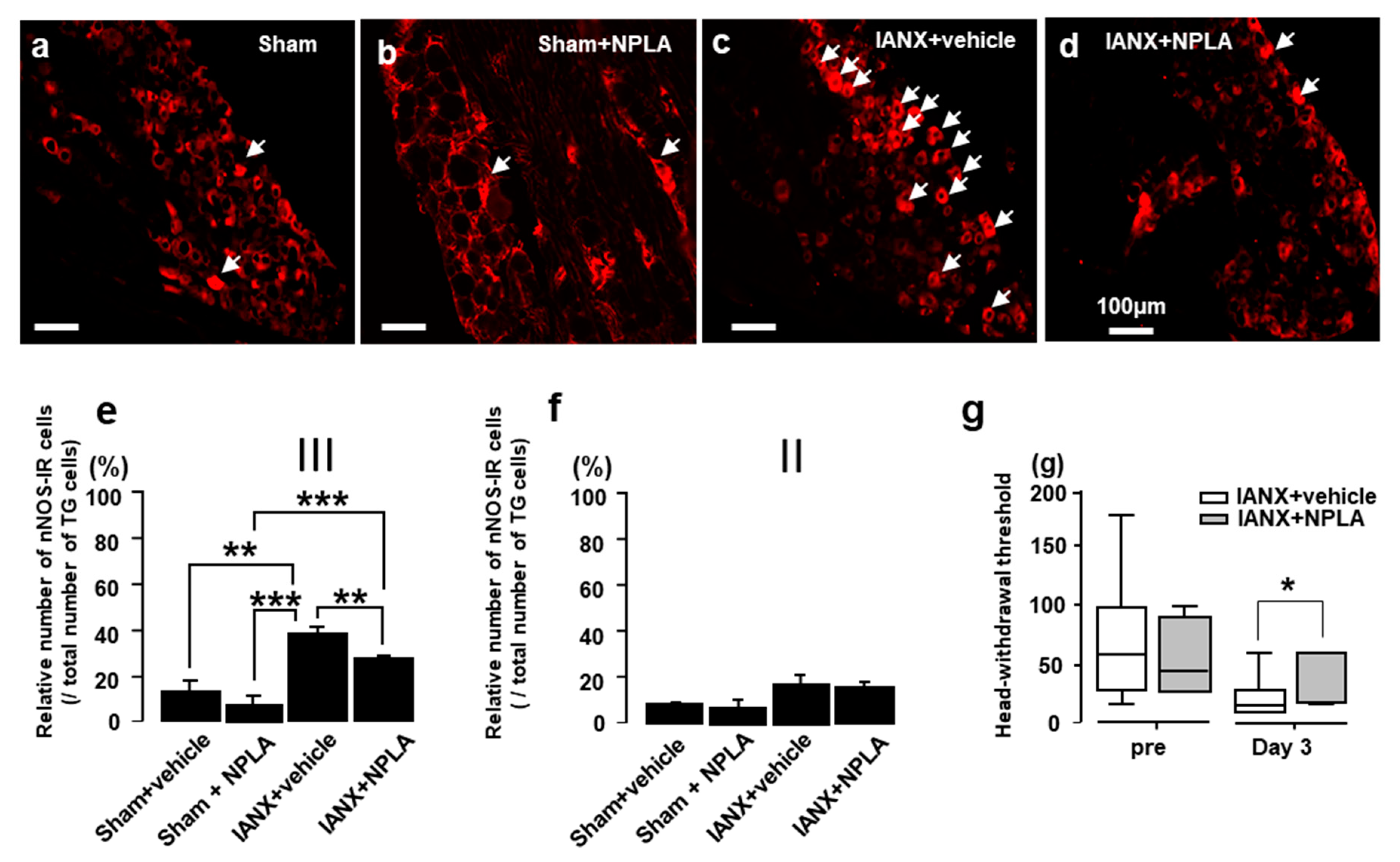
2.3. N-Propyl-L-Arginine (NPLA) Effects on nNOS Expression and MHWT
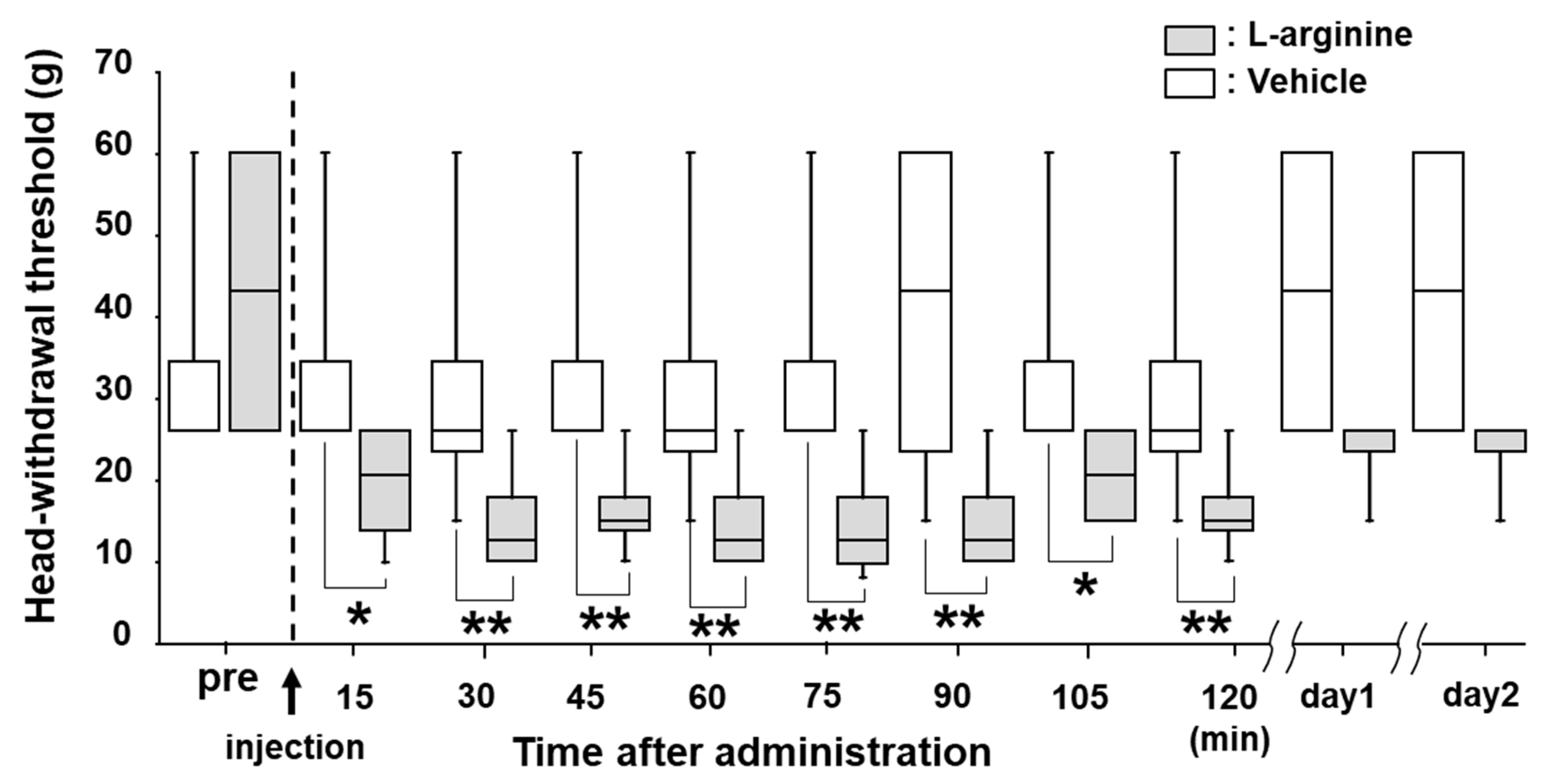
2.4. Effects of L-Arginine Administration on MHWT
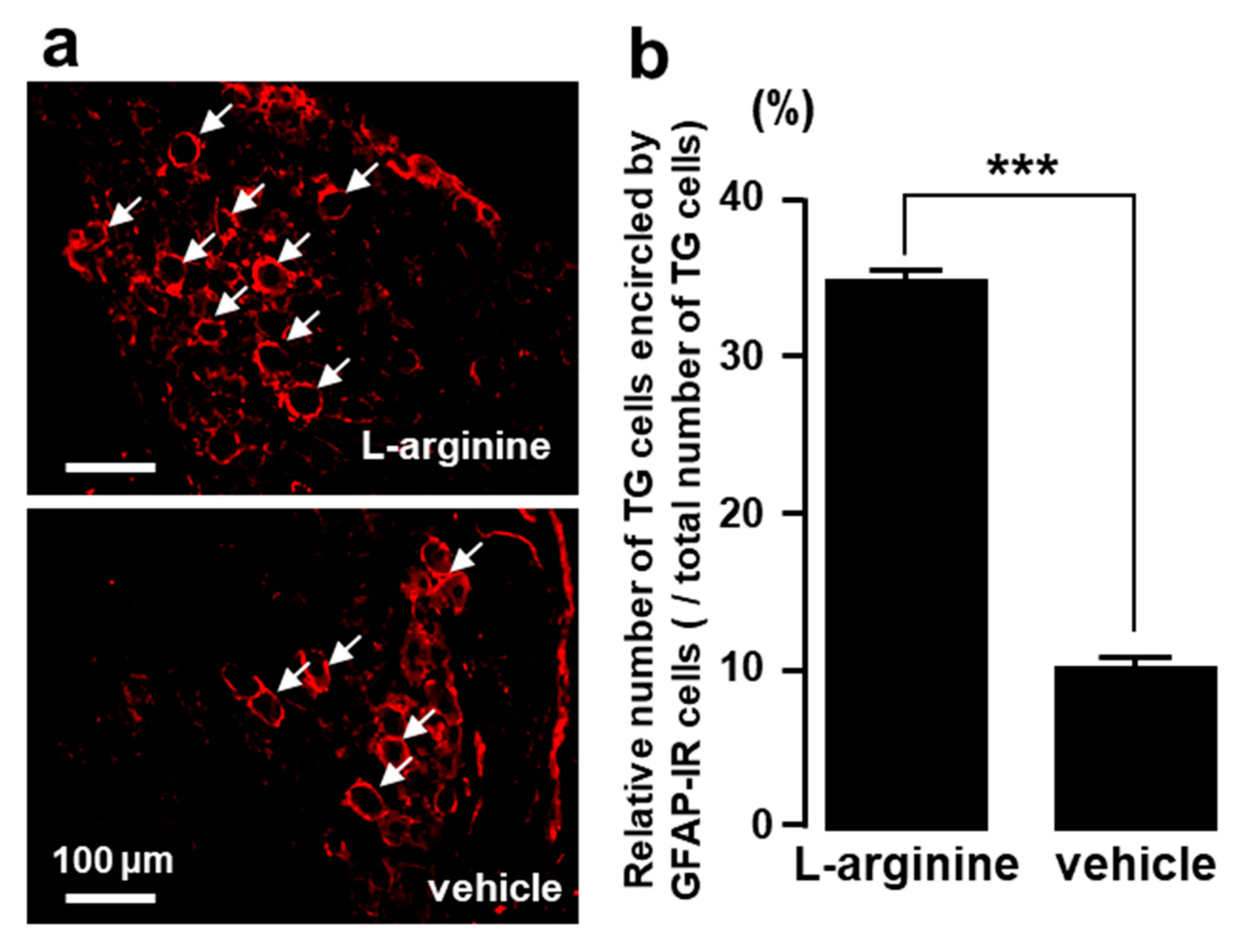
2.5. TG Cells Encircled by GFAP-IR Cells after L-Arginine Administration
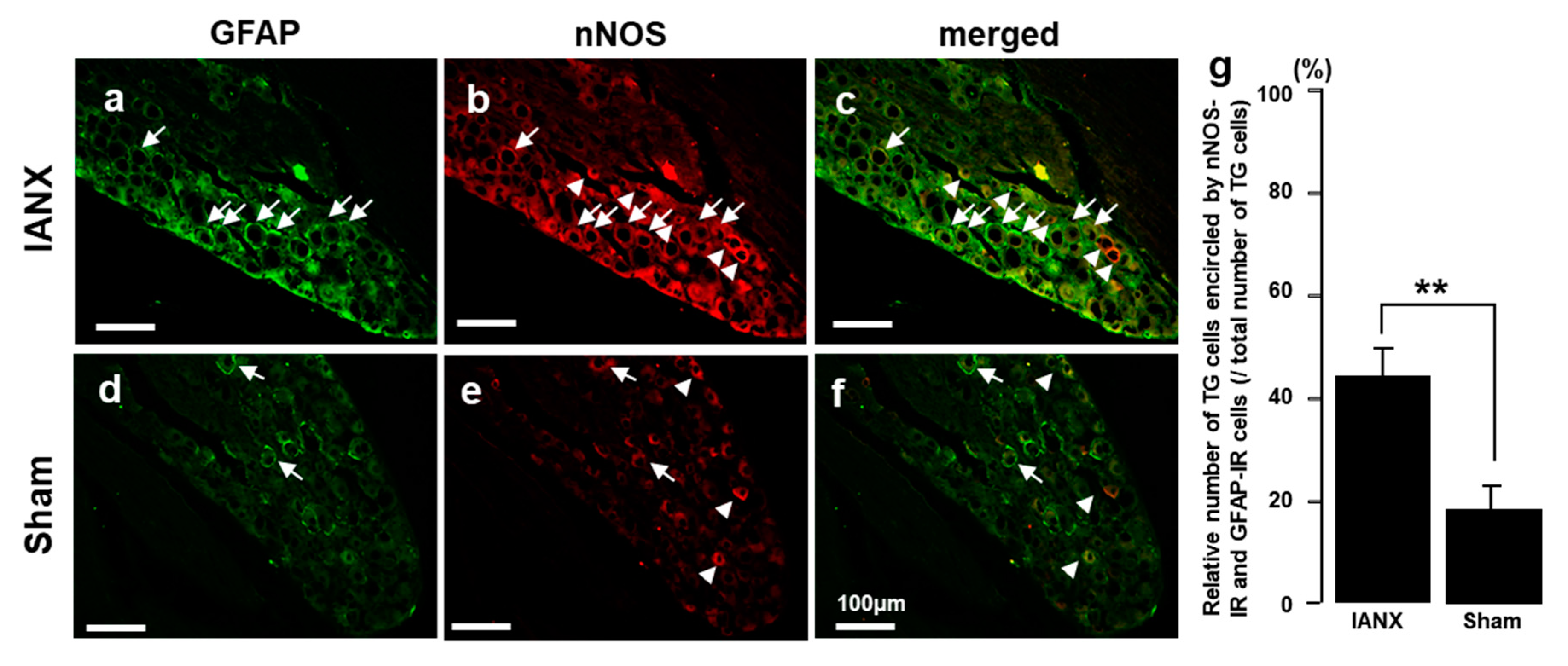
2.6. nNOS-IR Cells Encircled by GFAP-IR Cells
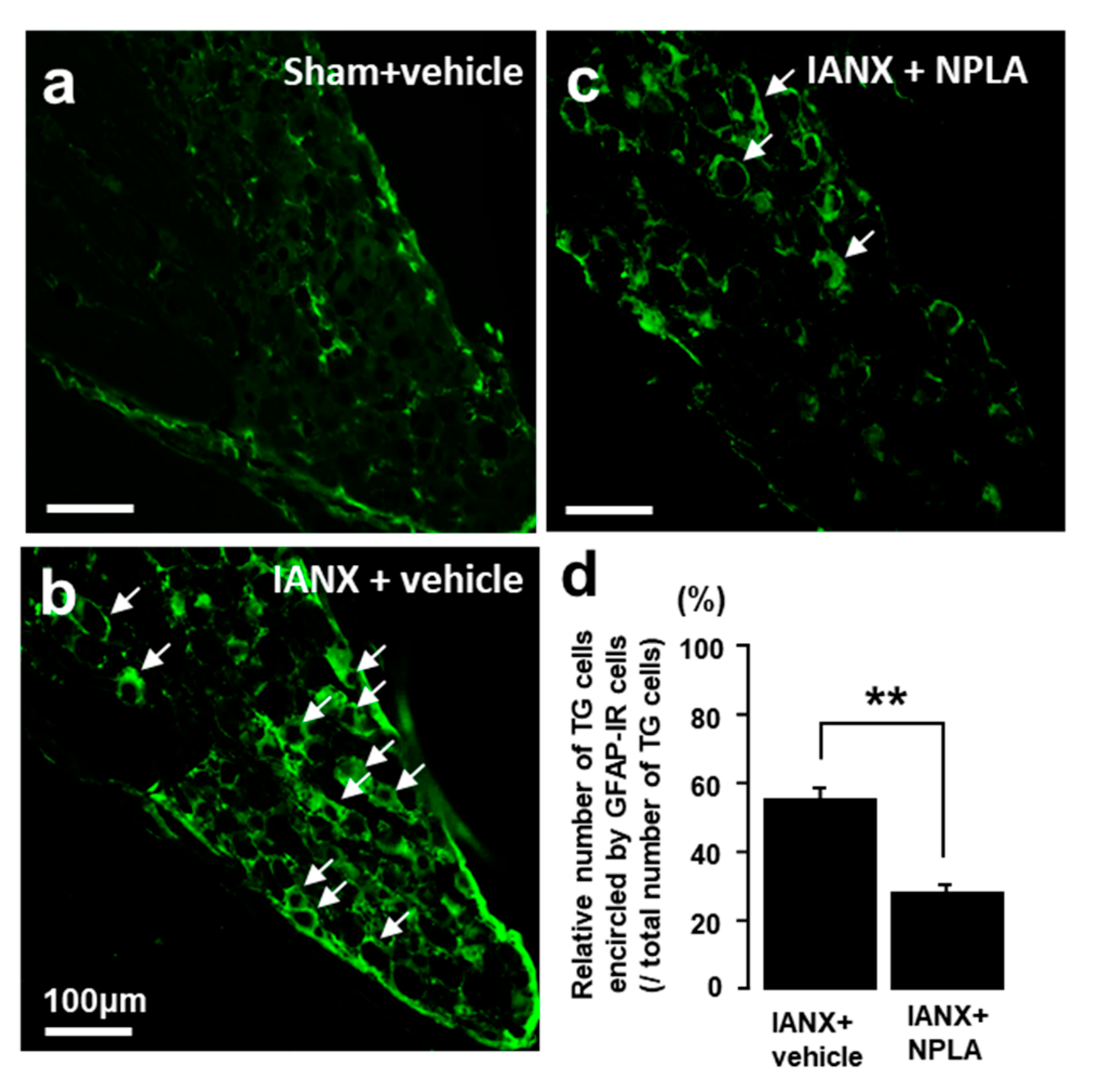
2.7. Effects of NPLA on GFAP Expression in the TG
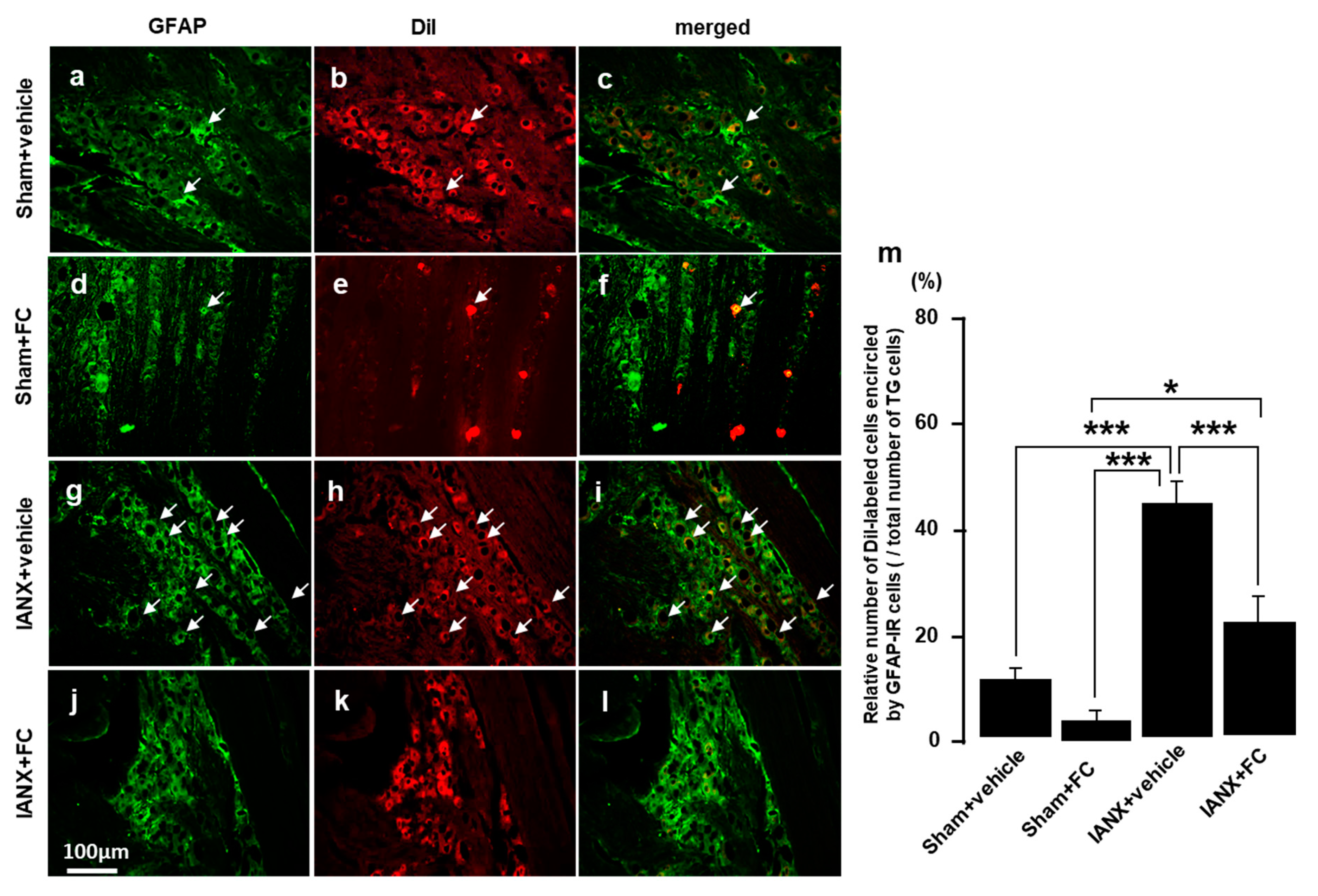
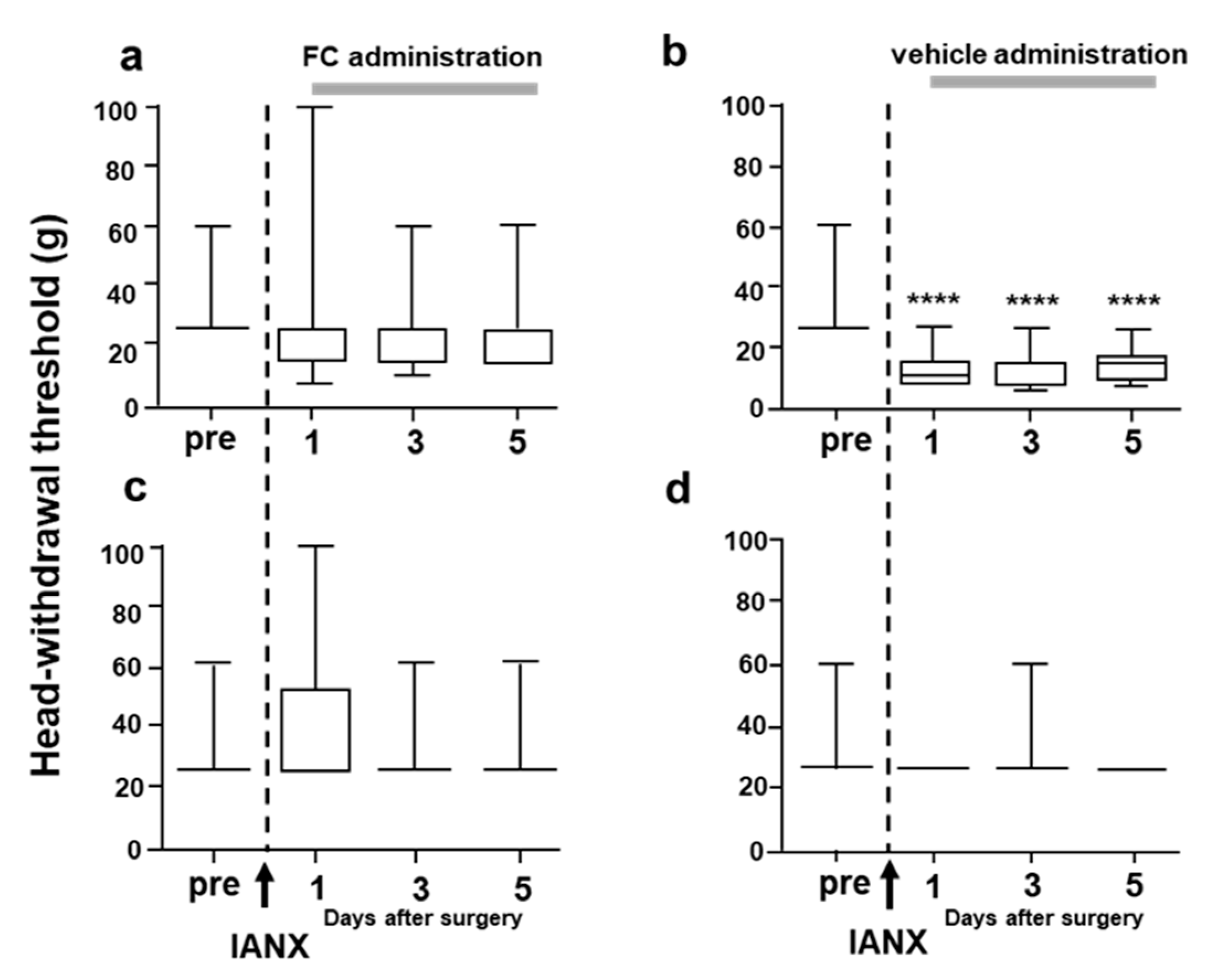
2.8. Effects of Fluorocitrate (FC) on GFAP Expression and MHWT
3. Discussion
3.1. Satellite Glial Cell Activation
3.2. NO Synthesis
3.3. NO and Satellite Glial Cell Activation
4. Materials and Methods
4.1. Animals
4.2. Inferior Alveolar Nerve Transection
4.3. Behavioral Assessment
4.4. Measurement of Nitrate and Nitrite
4.5. Drug Administration into the TG
4.6. Immunohistochemistry
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Iwata, K.; Imamura, Y.; Honda, K.; Shinoda, M. Physiological mechanisms of neuropathic pain: The orofacial region. Int. Rev. Neurobiol. 2011, 97, 227–250. [Google Scholar] [PubMed]
- Nakagawa, K.; Takeda, M.; Tsuboi, Y.; Kondo, M.; Kitagawa, J.; Matsumoto, S.; Kobayashi, A.; Sessle, B.J.; Shinoda, M.; Iwata, K. Alteration of primary afferent activity following inferior alveolar nerve transection in rats. Mol. Pain 2010, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafers, M.; Lee, D.H.; Brors, D.; Yaksh, T.L.; Sorkin, L.S. Increased sensitivity of injured and adjacent uninjured rat primary sensory neurons to exogenous tumor necrosis factor-alpha after spinal nerve ligation. J. Neurosci. 2003, 23, 3028–3038. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, Y.; Takeda, M.; Tanimoto, T.; Ikeda, M.; Matsumoto, S.; Kitagawa, J.; Teramoto, K.; Simizu, K.; Yamazaki, Y.; Shima, A.; et al. Alteration of the second branch of the trigeminal nerve activity following inferior alveolar nerve transection in rats. Pain 2004, 111, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Batbold, D.; Shinoda, M.; Honda, K.; Furukawa, A.; Koizumi, M.; Akasaka, R.; Yamaguchi, S.; Iwata, K. Macrophages in trigeminal ganglion contribute to ectopic mechanical hypersensitivity following inferior alveolar nerve injury in rats. J. Neuroinflamm. 2017, 14, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, T.; Iwai, H.; Kuramoto, E.; Yamanaka, A. Neuropeptides and ATP signaling in the trigeminal ganglion. Jpn. Dent. Sci. Rev. 2017, 53, 117–124. [Google Scholar] [CrossRef]
- Takeda, M.; Tanimoto, T.; Nasu, M.; Ikeda, M.; Kadoi, J.; Matsumoto, S. Activation of NK1 receptor of trigeminal root ganglion via substance P paracrine mechanism contributes to the mechanical allodynia in the temporomandibular joint inflammation in rats. Pain 2005, 116, 375–385. [Google Scholar] [CrossRef]
- Takeda, M.; Nasu, M.; Kanazawa, T.; Takahashi, M.; Shimazu, Y. Chemokine ligand 2/chemokine receptor 2 signaling in the trigeminal ganglia contributes to inflammatory hyperalgesia in rats. Neurosci. Res. 2018, 128, 25–32. [Google Scholar] [CrossRef]
- Vit, J.P.; Jasmin, L.; Bhargava, A.; Ohara, P.T. Satellite glial cells in the trigeminal ganglion as a determinant of orofacial neuropathic pain. Neuron Glia Biol. 2006, 2, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Iwata, K.; Katagiri, A.; Shinoda, M. Neuron-glia interaction is a key mechanism underlying persistent orofacial pain. J. Oral Sci. 2017, 59, 173–175. [Google Scholar] [CrossRef] [Green Version]
- Katagiri, A.; Shinoda, M.; Honda, K.; Toyofuku, A.; Sessle, B.J.; Iwata, K. Satellite glial cell P2Y12 receptor in the trigeminal ganglion is involved in lingual neuropathic pain mechanisms in rats. Mol. Pain 2012, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaji, K.; Shinoda, M.; Honda, K.; Unno, S.; Shimizu, N.; Iwata, K. Connexin 43 contributes to ectopic orofacial pain following inferior alveolar nerve injury. Mol. Pain 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garthwaite, J.; Boulton, C.L. Nitric oxide signaling in the central nervous system. Annu. Rev. Physiol. 1995, 57, 683–706. [Google Scholar] [CrossRef] [PubMed]
- Zundorf, G.; Reiser, G. Calcium dysregulation and homeostasis of neural calcium in the molecular mechanisms of neurodegenerative diseases provide multiple targets for neuroprotection. Antioxid. Redox Signal. 2011, 14, 1275–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Fei, M.; Cheng, C.; Yu, X.; Chen, M.; Shi, S.; Qin, J.; Guo, Z.; Shen, A. Spatiotemporal expression of PSD-95 and nNOS after rat sciatic nerve injury. Neurochem. Res. 2008, 33, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Roh, D.H.; Choi, S.R.; Yoon, S.Y.; Kang, S.Y.; Moon, J.Y.; Kwon, S.G.; Han, H.J.; Beitz, A.J.; Lee, J.H. Spinal neuronal NOS activation mediates sigma-1 receptor-induced mechanical and thermal hypersensitivity in mice: Involvement of PKC-dependent GluN1 phosphorylation. Br. J. Pharmacol. 2011, 163, 1707–1720. [Google Scholar] [CrossRef] [Green Version]
- Lohinai, Z.; Szekely, A.D.; Benedek, P.; Csillag, A. Nitric oxide synthase containing nerves in the cat and dog dental pulp and gingiva. Neurosci. Lett. 1997, 227, 91–94. [Google Scholar] [CrossRef]
- Sugiyama, T.; Shinoda, M.; Watase, T.; Honda, K.; Ito, R.; Kaji, K.; Urata, K.; Lee, J.; Ohara, K.; Takahashi, O.; et al. Nitric oxide signaling contributes to ectopic orofacial neuropathic pain. J. Dent. Res. 2013, 92, 1113–1117. [Google Scholar] [CrossRef]
- Iwata, K.; Tsuboi, Y.; Shima, A.; Harada, T.; Ren, K.; Kanda, K.; Kitagawa, J. Central neuronal changes after nerve injury: Neuroplastic influences of injury and aging. J. Orofac. Pain 2004, 18, 293–298. [Google Scholar]
- Iwata, K.; Takeda, M.; Oh, S.B.; Shinoda, M. Neurophysiology of Orofacial Pain. In Contemporary Oral Medicine; Farah, C., Balasubramaniam, R., McCullough, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–23. [Google Scholar]
- Valmaseda-Castellon, E.; Berini-Aytes, L.; Gay-Escoda, C. Inferior alveolar nerve damage after lower third molar surgical extraction: A prospective study of 1117 surgical extractions. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2001, 92, 377–383. [Google Scholar] [CrossRef]
- Iwata, K.; Imai, T.; Tsuboi, Y.; Tashiro, A.; Ogawa, A.; Morimoto, T.; Masuda, Y.; Tachibana, Y.; Hu, J. Alteration of medullary dorsal horn neuronal activity following inferior alveolar nerve transection in rats. J. Neurophysiol. 2001, 86, 2868–2877. [Google Scholar] [CrossRef] [PubMed]
- Aley, K.O.; McCarter, G.; Levine, J.D. Nitric oxide signaling in pain and nociceptor sensitization in the rat. J. Neurosci. 1998, 18, 7008–7014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, D.; Zochodne, D.W. Local nitric oxide synthase activity in a model of neuropathic pain. Eur. J. Neurosci. 1998, 10, 1846–1855. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, S.; Ossipov, M.H.; Johnson, K.W. The role of calcitonin gene-related peptide in peripheral and central pain mechanisms including migraine. Pain 2017, 158, 543–559. [Google Scholar] [CrossRef] [Green Version]
- Kolesnikov, Y.A.; Chereshnev, I.; Criesta, M.; Pan, Y.X.; Pasternak, G.W. Opposing actions of neuronal nitric oxide synthase isoforms in formalin-induced pain in mice. Brain Res. 2009, 1289, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Freire, M.A.; Guimaraes, J.S.; Leal, W.G.; Pereira, A. Pain modulation by nitric oxide in the spinal cord. Front. Neurosci. 2009, 3, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Kuboyama, K.; Tsuda, M.; Tsutsui, M.; Toyohara, Y.; Tozaki-Saitoh, H.; Shimokawa, H.; Yanagihara, N.; Inoue, K. Reduced spinal microglial activation and neuropathic pain after nerve injury in mice lacking all three nitric oxide synthases. Mol. Pain 2011, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Vause, C.V.; Durham, P.L. Calcitonin gene-related peptide stimulation of nitric oxide synthesis and release from trigeminal ganglion glial cells. Brain Res. 2008, 1196, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Vause, C.V.; Durham, P.L. CGRP stimulation of iNOS and NO release from trigeminal ganglion glial cells involves mitogen-activated protein kinase pathways. J. Neurochem. 2009, 110, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Radovits, T.; Szabo, G.; Mozes, M.M.; Rosivall, L.; Kokeny, G. Selective phosphodiesterase-5 (PDE-5) inhibitor vardenafil ameliorates renal damage in type 1 diabetic rats by restoring cyclic 3′,5′ guanosine monophosphate (cGMP) level in podocytes. Nephrol. Dial. Transplant. 2013, 28, 1751–1761. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Chapman, M.E.; Wideman, R.F., Jr. Evaluation of total plasma nitric oxide concentrations in broilers infused intravenously with sodium nitrite, lipopolysaccharide, aminoguanidine, and sodium nitroprusside. Poult. Sci. 2006, 85, 312–320. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Ohara, K.; Shinoda, M.; Hayashi, Y.; Kubo, A.; Sugawara, S.; Asano, S.; Soma, K.; Kanno, K.; Ando, M.; et al. Involvement of Satellite Cell Activation via Nitric Oxide Signaling in Ectopic Orofacial Hypersensitivity. Int. J. Mol. Sci. 2020, 21, 1252. https://doi.org/10.3390/ijms21041252
Lee J, Ohara K, Shinoda M, Hayashi Y, Kubo A, Sugawara S, Asano S, Soma K, Kanno K, Ando M, et al. Involvement of Satellite Cell Activation via Nitric Oxide Signaling in Ectopic Orofacial Hypersensitivity. International Journal of Molecular Sciences. 2020; 21(4):1252. https://doi.org/10.3390/ijms21041252
Chicago/Turabian StyleLee, Jun, Kinuyo Ohara, Masamichi Shinoda, Yoshinori Hayashi, Asako Kubo, Shiori Sugawara, Sayaka Asano, Kumi Soma, Kohei Kanno, Masatoshi Ando, and et al. 2020. "Involvement of Satellite Cell Activation via Nitric Oxide Signaling in Ectopic Orofacial Hypersensitivity" International Journal of Molecular Sciences 21, no. 4: 1252. https://doi.org/10.3390/ijms21041252
APA StyleLee, J., Ohara, K., Shinoda, M., Hayashi, Y., Kubo, A., Sugawara, S., Asano, S., Soma, K., Kanno, K., Ando, M., Koyama, R., Kimura, Y., Sakanashi, K., Iinuma, T., & Iwata, K. (2020). Involvement of Satellite Cell Activation via Nitric Oxide Signaling in Ectopic Orofacial Hypersensitivity. International Journal of Molecular Sciences, 21(4), 1252. https://doi.org/10.3390/ijms21041252