Chemoradiotherapy Increases Intratumor Heterogeneity of HPSE Expression in the Relapsed Glioblastoma Tumors

 , and
, and
Abstract
:1. Introduction
2. Results
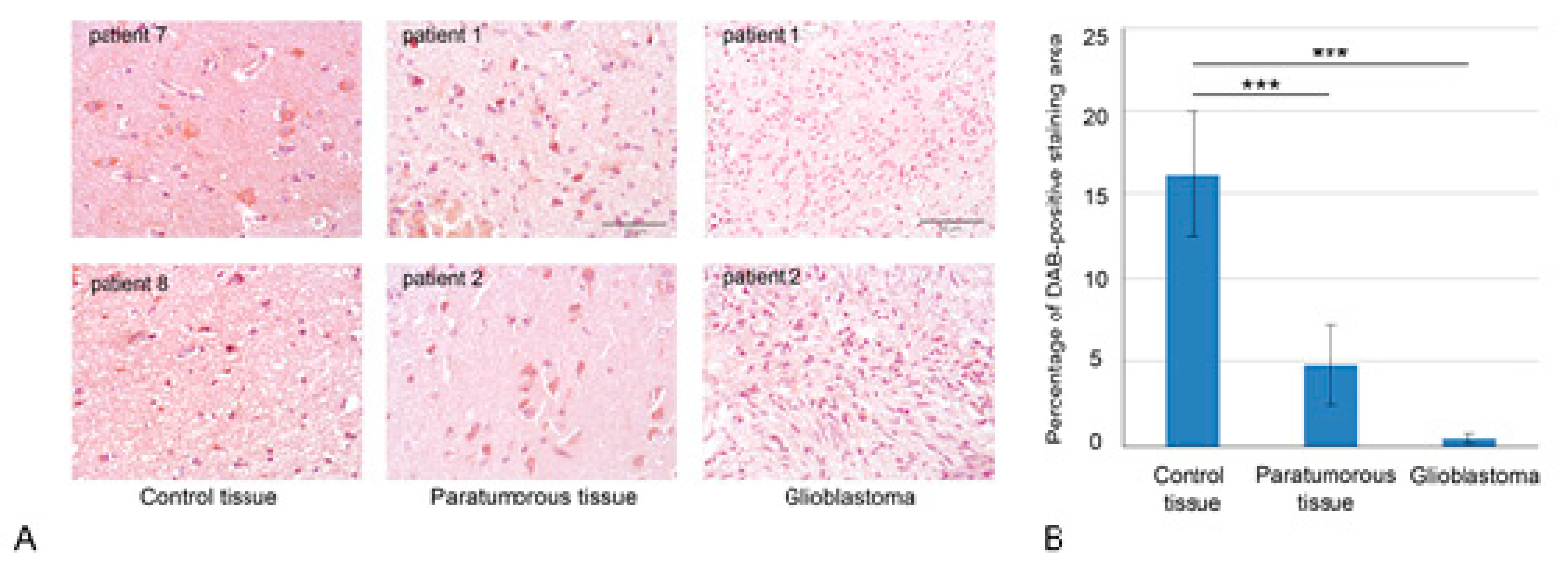
2.1. Chemoradiotherapy Affects Intertumor Heterogeneity of HPSE Content and Distribution
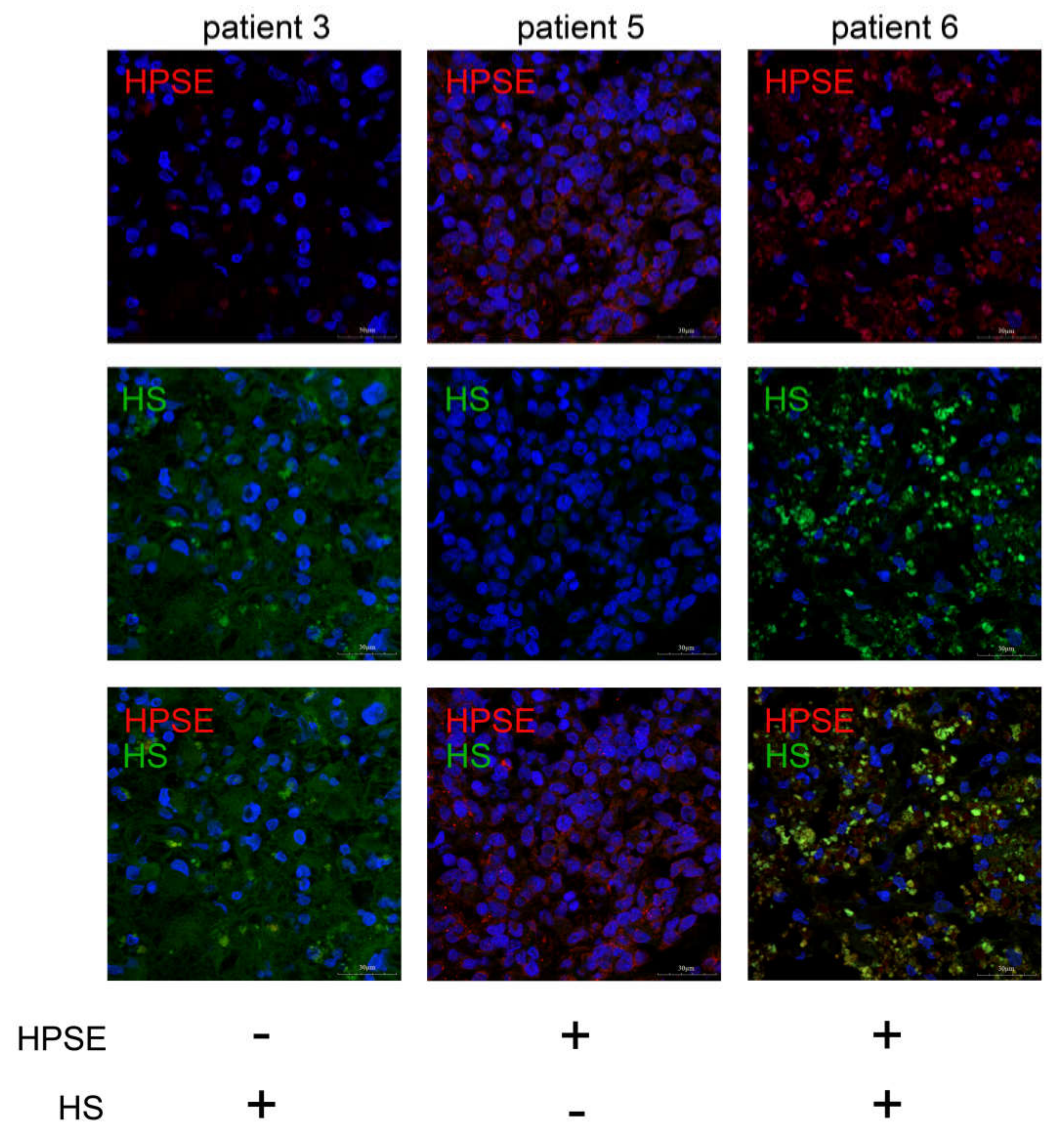
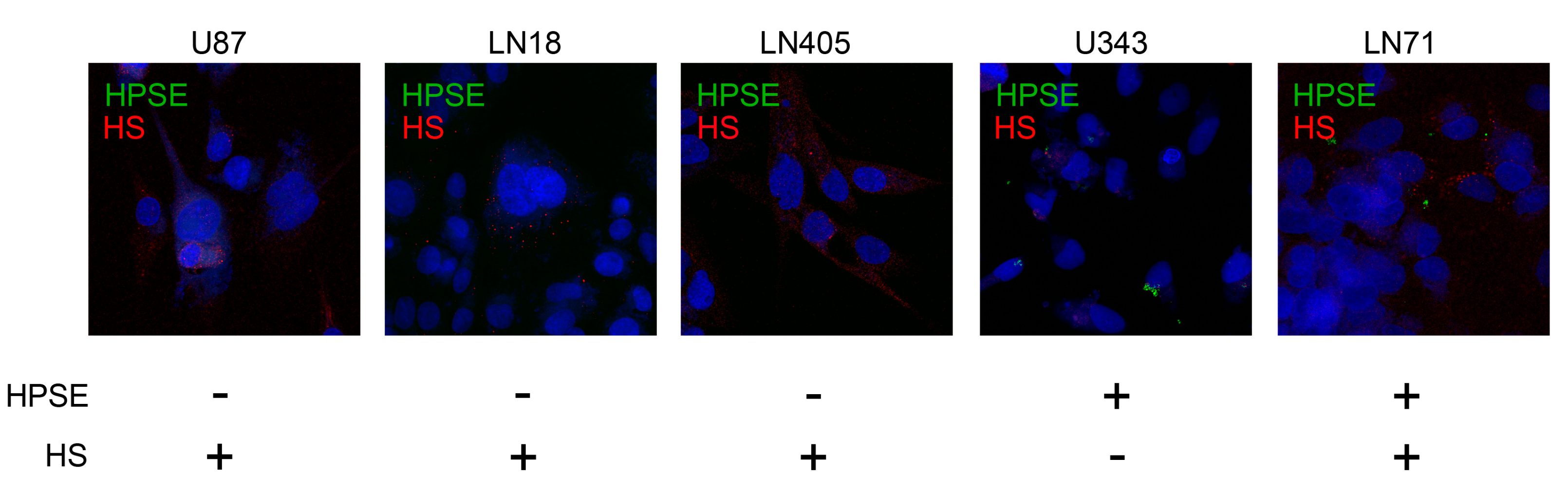
2.2. Interrelation of HPSE and HS Content in GBM Tumors and Cell Lines
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Immunostaining
4.3. Immunocytochemistry
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GBM | Glioblastoma multiforme |
| ECM | Extracellular Matrix |
| Heparanase | HPSE |
| HS | Heparan Sulfate |
References
- Hottinger, A.F.; Stupp, R.; Homicsko, K. Standards of care and novel approaches in the management of glioblastoma multiforme. Chin. J. Cancer. 2014, 33, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.E. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol Nurs. 2016, 20, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar]
- Kazanskaya, G.M.; Tsidulko, A.Y.; Volkov, A.M.; Kiselev, R.S.; Suhovskih, A.V.; Kobozev, V.V.; Gaytan, A.S.; Aidagulova, S.V.; Krivoshapkin, A.L.; Grigorieva, E.V. Heparan sulfate accumulation and perlecan/HSPG2 up-regulation in tumor tissue predict low relapse-free survival for patients with glioblastoma. Histochem Cell Biol. 2018, 149, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, R.D.; Elkin, M.; Rapraeger, A.C.; Ilan, N.; Vlodavsky, I. Heparanase regulation of cancer, autophagy and inflammation: New mechanisms and targets for therapy. FEBS J. 2017, 284, 42–55. [Google Scholar] [CrossRef]
- Li, J.P.; Kusche-Gullberg, M. Heparan sulfate: Biosynthesis, structure, and function. Int. Rev. Cell Mol. Biol. 2016, 325, 215–273. [Google Scholar]
- Wade, A.; Robinson, A.E.; Engler, J.R.; Petritsch, C.; James, C.D.; Phillips, J.J. Proteoglycans and their roles in brain cancer. FEBS J. 2013, 280, 2399–2417. [Google Scholar] [CrossRef] [Green Version]
- Vlodavsky, I.; Singh, P.; Boyango, I.; Gutter-Kapon, L.; Elkin, M.; Sanderson, R.D.; Ilan, N. Heparanase: From basic research to therapeutic applications in cancer and inflammation. Drug Resist. Updat. 2016, 29, 54–75. [Google Scholar] [CrossRef]
- El-Assal, O.N.; Yamanoi, A.; Ono, T.; Kohno, H.; Nagasue, N. The clinicopathological significance of heparanase and basic fibroblast growth factor expressions in hepatocellular carcinoma. Clincancer Res. 2001, 7, 1299–1305. [Google Scholar]
- Kelly, T.; Miao, H.Q.; Yang, Y.; Navarro, E.; Kussie, P.; Huang, Y.; MacLeod, V.; Casciano, J.; Joseph, L.; Zhan, F.; et al. High heparanase activity in multiple myeloma is associated with elevated microvessel density. Cancer Res. 2003, 63, 8749–8756. [Google Scholar]
- Cohen, I.; Pappo, O.; Elkin, M.; San, T.; Bar-Shavit, R.; Hazan, R.; Peretz, T.; Vlodavsky, I.; Abramovitch, R. Heparanase promotes growth, angiogenesis and survival of primary breast tumors. Int. J. Cancer. 2006, 118, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Shafat, I.; Risberg, B.; Ilan, N.; Trope, C.G.; Vlodavsky, I.; Reich, R. Heparanase expression correlates with poor survival in metastatic ovarian carcinoma. Gynecol Oncol. 2007, 104, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Nelson, K.K.; deCarvalho, A.C.; Kalkanis, S.N. Heparanase expression of glioma in human and animal models. J. Neurosurg. 2010, 113, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Xiong, A.; Spyrou, A.; Wicher, G.; Marinescu, V.D.; Edqvist, P.D.; Zhang, L.; Essand, M.; Dimberg, A.; Smits, A.; et al. Heparanase Promotes Glioma Progression and Is Inversely Correlated with Patient Survival. Mol. Cancer Res. 2016, 14, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Spyrou, A.; Kundu, S.; Haseeb, L.; Yu, D.; Olofsson, T.; Dredge, K.; Hammond, E.; Barash, U.; Vlodavsky, I.; Forsberg-Nilsson, K. Inhibition of Heparanase in Pediatric Brain Tumor Cells Attenuates their Proliferation, Invasive Capacity, and In Vivo Tumor Growth. Mol. Cancer 2017, 16, 1705–1716. [Google Scholar] [CrossRef] [Green Version]
- Ueno, Y.; Yamamoto, M.; Vlodavsky, I.; Pecker, I.; Ohshima, K.; Fukushima, T. Decreased expression of heparanase in glioblastoma multiforme. J. Neurosurg. 2005, 102, 513–521. [Google Scholar] [CrossRef]
- Zetser, A.; Bashenko, Y.; Miao, H.Q.; Vlodavsky, I.; Ilan, N. Heparanase affects adhesive and tumorigenic potential of human glioma cells. Cancer Res. 2003, 63, 7733–7741. [Google Scholar]
- Phillips, J.J. Novel therapeutic targets in the brain tumor. Oncotarget. 2012, 3, 568–575. [Google Scholar] [CrossRef]
- Ushakov, V.S.; Tsidulko, A.Y.; de La Bourdonnaye, G.; Kazanskaya, G.M.; Volkov, A.M.; Kiselev, R.S.; Kobozev, V.V.; Kostromskaya, D.V.; Gaytan, A.S.; Krivoshapkin, A.L.; et al. Heparan Sulfate Biosynthetic System Is Inhibited in Human Glioma Due to EXT1/2 and HS6ST1/2 Down-Regulation. Int. J. Mol. Sci. 2017, 18, 2301. [Google Scholar] [CrossRef] [Green Version]
- Hong, X.; Jiang, F.; Kalkanis, S.N.; Zhang, Z.G.; Zhang, X.; Zheng, X.; Jiang, H.; Mikkelsen, T.; Chopp, M. Increased chemotactic migration and growth in heparanase-overexpressing human U251n glioma cells. J. Exp. Clin. Cancer Res. 2008, 27, 23. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Kukula, A.K.; Toyoshima, M.; Nakajima, M. Genomic organization and chromosome localization of the newly identified human heparanase gene. Gene 2000, 253, 171–178. [Google Scholar] [CrossRef]
- Nasser, N.J.; Nevo, E.; Shafat, I.; Ilan, N.; Vlodavsky, I.; Avivi, A. Adaptive evolution of heparanase in hypoxia-tolerant Spalax: Gene cloning and identification of a unique splice variant. Proc. Natl. Acad. Sci. USA 2005, 102, 15161–15166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasser, N.J.; Avivi, A.; Shushy, M.; Vlodavsky, I.; Nevo, E. Cloning, expression, and characterization of an alternatively spliced variant of human heparanase. Biochem. Res. Commun. 2007, 354, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolesi, G.E.; Michaiel, G.; McFarlane, S. Two heparanase splicing variants with distinct properties are necessary in early Xenopus development. J. Biol. Chem. 2008, 283, 16004–16016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasser, N.J.; Avivi, A.; Shafat, I.; Edovitsky, E.; Zcharia, E.; Ilan, N.; Vlodavsky, I.; Nevo, E. Alternatively spliced Spalax heparanase inhibits extracellular matrix degradation, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2009, 106, 2253–2258. [Google Scholar] [CrossRef] [Green Version]
- Hong, X.; Nelson, K.; Lemke, N.; Kalkanis, S.N. Heparanase expression is associated with histone modifications in glioblastoma. Int. J. Oncol. 2012, 40, 494–500. [Google Scholar]
- Shteper, P.J.; Zcharia, E.; Ashhab, Y.; Peretz, T.; Vlodavsky, I.; Ben-Yehuda, D. Role of promoter methylation in regulation of the mammalian heparanase gene. Oncogene 2003, 22, 7737–7749. [Google Scholar] [CrossRef] [Green Version]
- Ogishima, T.; Shiina, H.; Breault, J.E.; Tabatabai, L.; Bassett, W.W.; Enokida, H.; Li, L.C.; Kawakami, T.; Urakami, S.; Ribeiro-Filho, L.A.; et al. Increased heparanase expression is caused by promoter hypomethylation and up-regulation of transcriptional factor early growth response-1 in human prostate cancer. Clin. Cancer Res. 2005, 11, 1028–1036. [Google Scholar]
- Arvatz, G.; Barash, U.; Nativ, O.; Ilan, N.; Vlodavsky, I.; Jiao, W. Post-transcriptional regulation of heparanase gene expression by a 3′ AU-rich element. FASEB J. 2010, 24, 4969–4976. [Google Scholar] [CrossRef] [Green Version]
- Jiao, W.; Chen, Y.; Song, H.; Li, D.; Mei, H.; Yang, F.; Fang, E.; Wang, X.; Huang, K.; Zheng, L.; et al. HPSE enhancer RNA promotes cancer progression through driving chromatin looping and regulating hnRNPU/p300/EGR1/HPSE axis. Oncogene 2018, 37, 2728–2745. [Google Scholar] [CrossRef]
- Shao, C.; Shi, X.; Phillips, J.J.; Zaia, J. Mass spectral profiling of glycosaminoglycans from histological tissue surfaces. Anal. Chem. 2013, 85, 10984–10991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, V.M.; Wade, A.; McKinney, A.; Chen, K.; Lindberg, O.R.; Engler, J.R.; Persson, A.I.; Phillips, J.J. Heparan Sulfate Glycosaminoglycans in Glioblastoma Promote Tumor Invasion. Mol. Cancer Res. 2017, 15, 1623–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | N (patients) = 8 | % |
|---|---|---|
| Age | ||
| Median (range) | 50.5 (38–68) | |
| > 50 median age | 4 | 50 |
| < 50 median age | 4 | 50 |
| Sex | ||
| Male | 4 | 50 |
| Female | 4 | 50 |
| Tumor development | ||
| Primary | 5 | 62.5 |
| Secondary | 3 | 37.5 |
| Death | ||
| Yes | 7 | 87.5 |
| No | 1 | 12.5 |
| IDH status | ||
| wild | 8 | 100 |
| mutant | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suhovskih, A.V.; Kazanskaya, G.M.; Volkov, A.M.; Tsidulko, A.Y.; Aidagulova, S.V.; Grigorieva, E.V. Chemoradiotherapy Increases Intratumor Heterogeneity of HPSE Expression in the Relapsed Glioblastoma Tumors. Int. J. Mol. Sci. 2020, 21, 1301. https://doi.org/10.3390/ijms21041301
Suhovskih AV, Kazanskaya GM, Volkov AM, Tsidulko AY, Aidagulova SV, Grigorieva EV. Chemoradiotherapy Increases Intratumor Heterogeneity of HPSE Expression in the Relapsed Glioblastoma Tumors. International Journal of Molecular Sciences. 2020; 21(4):1301. https://doi.org/10.3390/ijms21041301
Chicago/Turabian StyleSuhovskih, Anastasia V., Galina M. Kazanskaya, Alexander M. Volkov, Alexandra Y. Tsidulko, Svetlana V. Aidagulova, and Elvira V. Grigorieva. 2020. "Chemoradiotherapy Increases Intratumor Heterogeneity of HPSE Expression in the Relapsed Glioblastoma Tumors" International Journal of Molecular Sciences 21, no. 4: 1301. https://doi.org/10.3390/ijms21041301
APA StyleSuhovskih, A. V., Kazanskaya, G. M., Volkov, A. M., Tsidulko, A. Y., Aidagulova, S. V., & Grigorieva, E. V. (2020). Chemoradiotherapy Increases Intratumor Heterogeneity of HPSE Expression in the Relapsed Glioblastoma Tumors. International Journal of Molecular Sciences, 21(4), 1301. https://doi.org/10.3390/ijms21041301