Structural Changes and Astrocyte Response of the Lateral Geniculate Nucleus in a Ferret Model of Ocular Hypertension

 and
and
Abstract
:1. Introduction
2. Results
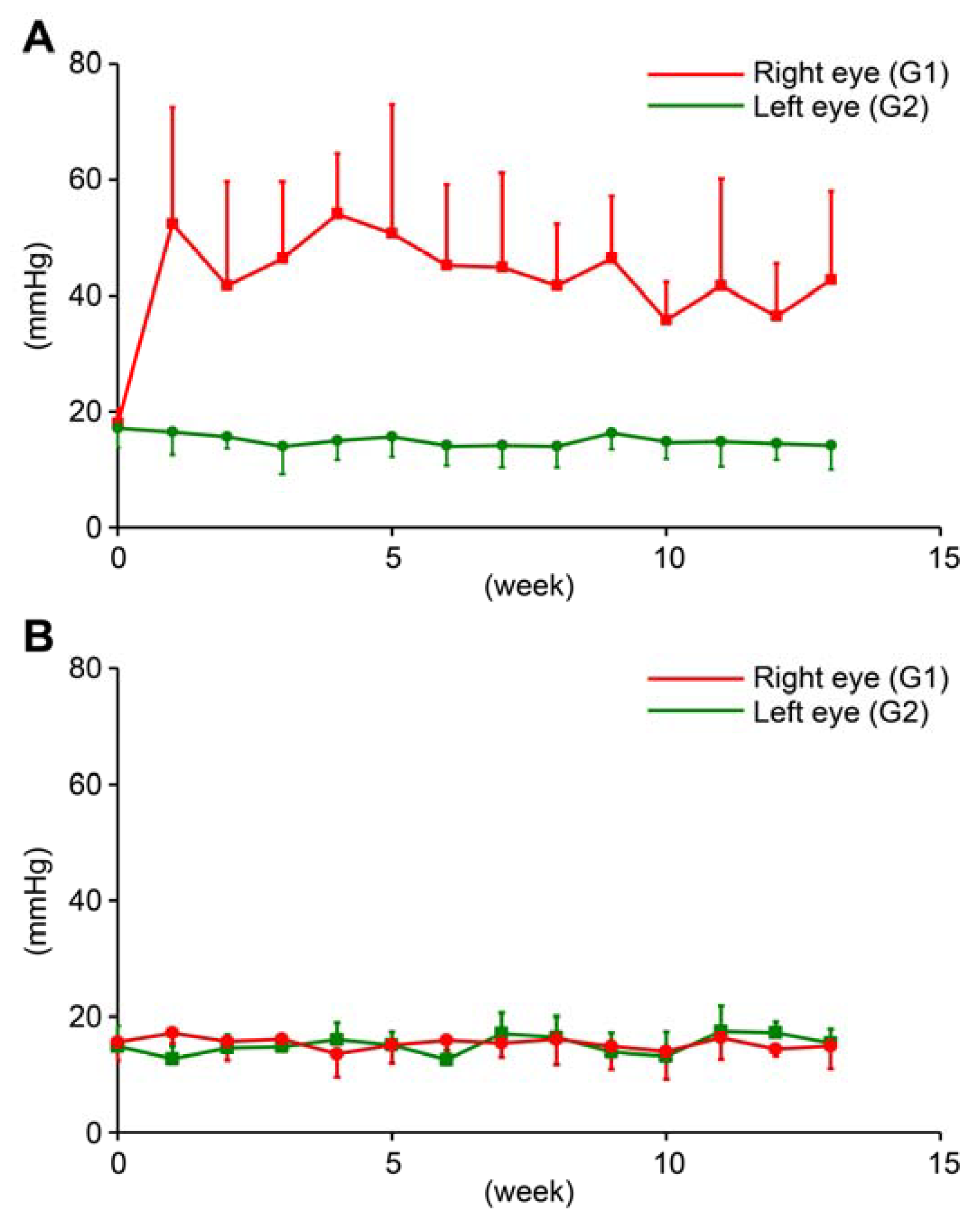
2.1. IOP of the OH Model
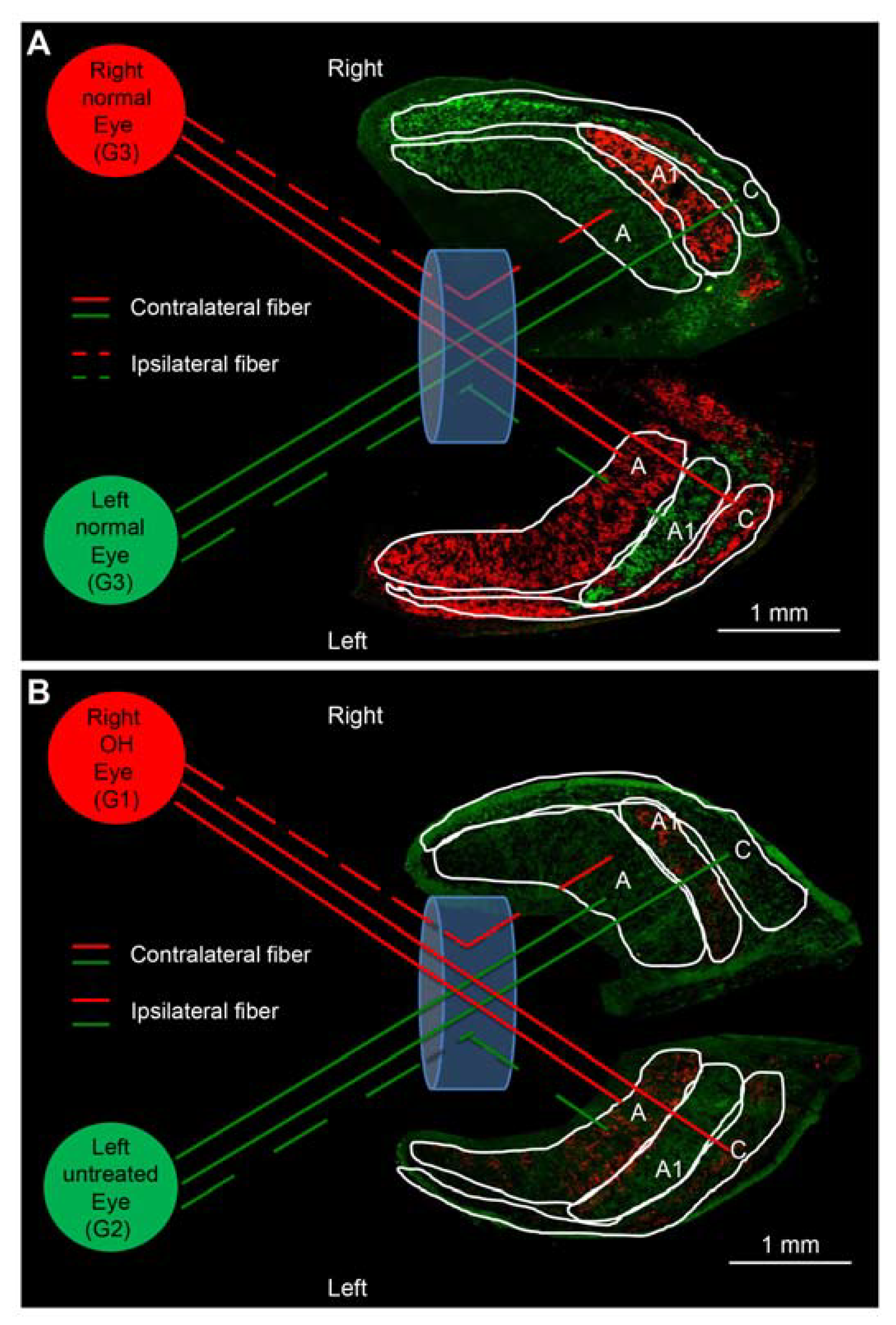
2.2. Microscopic Analysis of the Visual Tract in the OH Model
2.2.1. Histological Analysis
2.2.2. Quantitative Analysis of A, A1, and C Layers of the LGN in the OH Model
2.3. Immunological Analysis of the Visual Tract
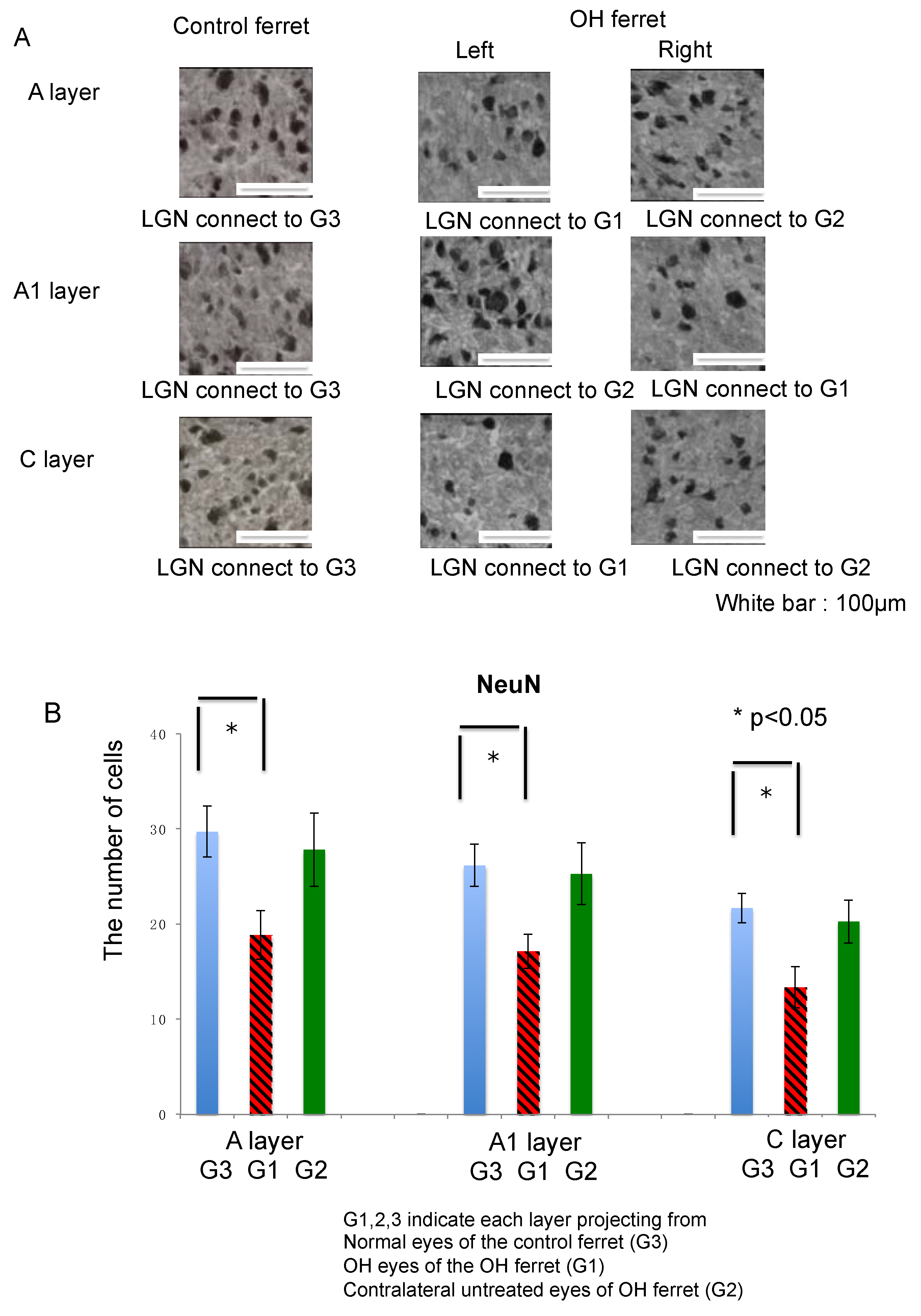
2.3.1. Neurons
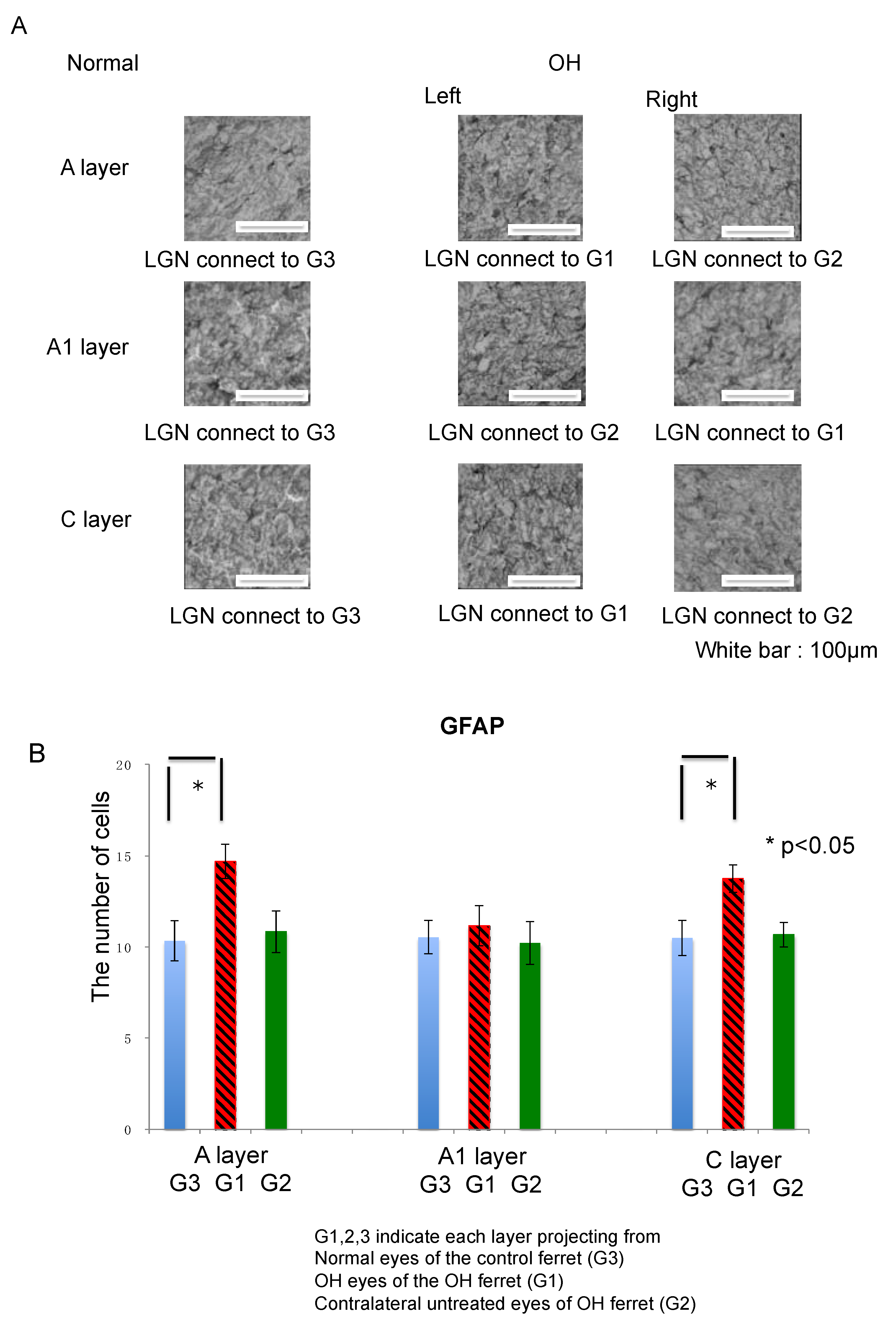
2.3.2. Astrocytes
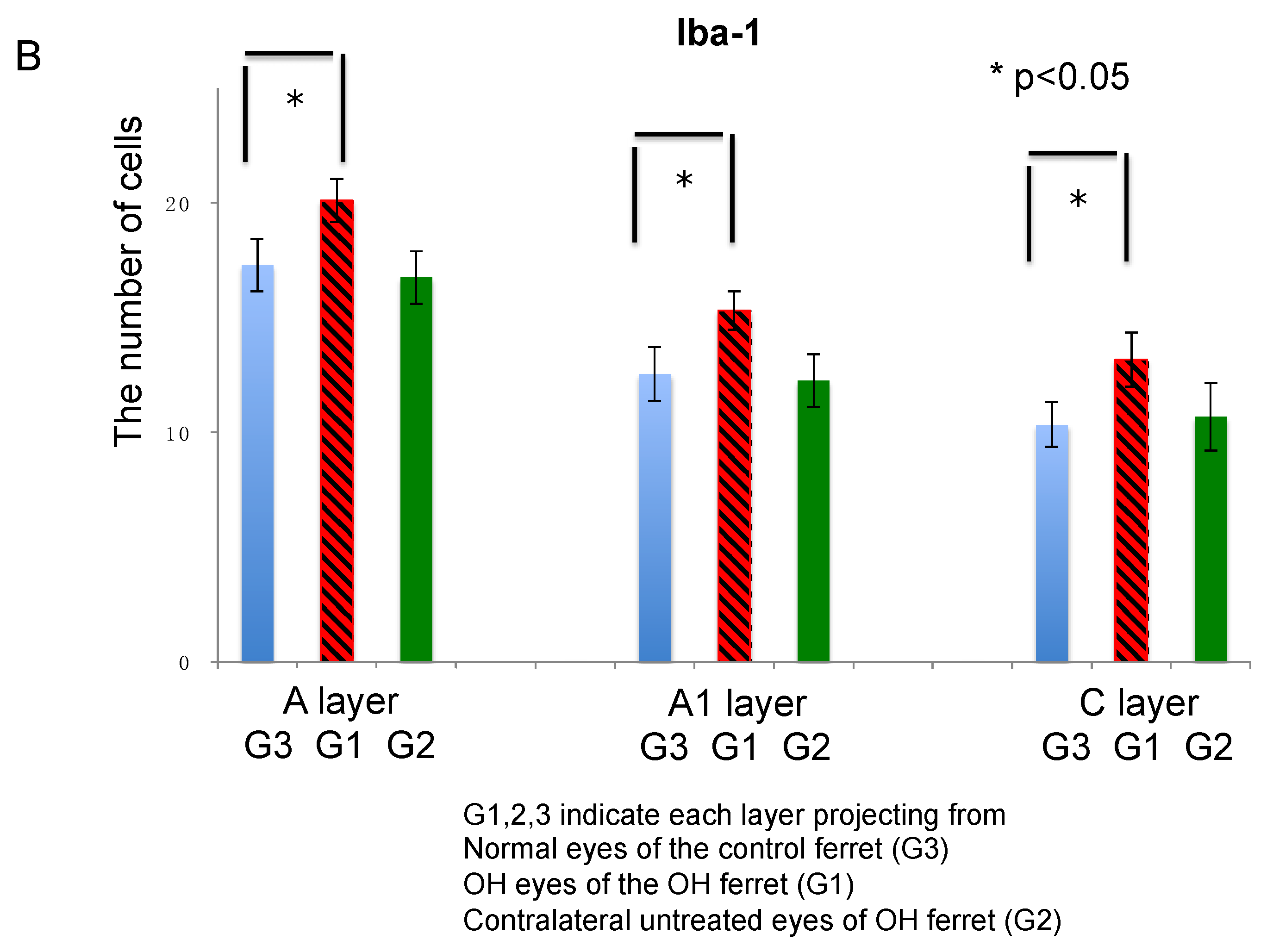
2.3.3. Microglia
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of Conjunctival Cells
4.3. Injection of Cultured Cells and Treatment of Eyes
4.4. Development of the Ferret Model of OH
4.5. Tracer Injection
4.6. Microscopic Analysis of the Visual Tract in the OH Model
4.7. Immunohistochemistry of the Visual Tract in the OH Model
4.8. Quantification
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LGN | lateral geniculate nucleus |
| OH | ocular hypertension |
| GFAP | glial fibrillary acidic protein |
| GON | glaucoma optic neuropathy |
| IOP | intraocular pressure |
| POAG | primary open angle glaucoma |
| CNS | Central nervous system |
| DMEM | Dulbecco’s Minimum Essential Medium |
| FBS | fetal bovine serum |
| PFA | paraformaldehyde |
| GFAP | anti-glial fibrillary acidic protein |
References
- Weinreb, R.N.; Friedman, D.S.; Fechtner, R.D.; Cioffi, G.A.; Coleman, A.L.; Girkin, C.A.; Liebmann, J.M.; Singh, K.; Wilson, M.R.; Wilson, R.; et al. Risk assessment in the management of patients with ocular hypertension. Am. J. Ophthalmol. 2004, 138, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Ly, T.; Zhang, Q.; Kaufman, P.L.; Weinreb, R.N.; Yücel, Y.H. Chronic ocular hypertension induces dendrite pathology in the lateral geniculate nucleus of the brain. Exp. Eye Res. 2007, 84, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Shimazawa, M.; Chen, Y.N.; Tsuruma, K.; Yamashima, T.; Araie, M.; Hara, H. Morphological changes in the visual pathway induced by experimental glaucoma in Japanese monkeys. Exp. Eye Res. 2009, 89, 246–255. [Google Scholar] [CrossRef]
- Yücel, Y.H.; Zhang, Q.; Weinreb, R.N.; Kaufman, P.L.; Gupta, N. Effects of retinal ganglion cell loss on magno-, parvo-, koniocellular pathways in the lateral geniculate nucleus and visual cortex in glaucoma. Prog. Retin. Eye Res. 2003, 22, 465–481. [Google Scholar] [CrossRef]
- Sasaoka, M.; Nakamura, K.; Shimazawa, M.; Ito, Y.; Araie, M.; Hara, H. Changes in visual fields and lateral geniculate nucleus in monkey laser-induced high intraocular pressure model. Exp. Eye Res. 2008, 86, 770–782. [Google Scholar] [CrossRef]
- Weber, A.J.; Chen, H.; Hubbard, W.C.; Kaufman, P.L. Experimental glaucoma and cell size, density, and number in the primate lateral geniculate nucleus. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1370–1379. [Google Scholar]
- Yücel, Y.H.; Zhang, Q.; Gupta, N.; Kaufman, P.L.; Weinreb, R.N. Loss of neurons in magnocellular and parvocellular layers of the lateral geniculate nucleus in glaucoma. Arch. Ophthalmol. 2000, 118, 378–384. [Google Scholar] [CrossRef]
- Sun, X.; Cheng, F.; Meng, B.; Yang, B.; Song, W.; Yuan, H. Pregnenolone sulfate decreases intraocular pressure and changes expression of sigma receptor in a model of chronic ocular hypertension. Mol. Biol. Rep. 2012, 39, 6607–6614. [Google Scholar] [CrossRef]
- Hains, B.C.; Waxman, S.G. Neuroprotection by sodium channel blockade with phenytoin in an experimental model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4164–4169. [Google Scholar] [CrossRef] [Green Version]
- Aihara, M. The use of mice in glaucoma research-to clarify the mechanism of intraocular pressure regulation and retinal ganglion cell damage. Nihon Ganka Gakkai Zasshi 2010, 114, 217–246. [Google Scholar]
- Aihara, M.; Lindsey, J.D.; Weinreb, R.N. Experimental mouse ocular hypertension: Establishment of the model. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4314–4320. [Google Scholar] [CrossRef]
- Aihara, M.; Lindsey, J.D.; Weinreb, R.N. Ocular hypertension in mice with a targeted type I collagen mutation. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1581–1585. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Tanabe, T.; Yoshimura, N. A rat model of glaucoma induced by episcleral vein ligation. Exp. Eye Res. 2006, 83, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Gross, R.L.; Ji, J.; Chang, P.; Pennesi, M.E.; Yang, Z.; Zhang, J.; Wu, S.M. A mouse model of elevated intraocular pressure: Retina and optic nerve findings. Trans. Am. Ophthalmol. Soc. 2003, 101, 163–169. [Google Scholar]
- Grozdanic, S.D.; Betts, D.M.; Sakaguchi, D.S.; Allbaugh, R.A.; Kwon, Y.H.; Kardon, R.H. Laser-induced mouse model of chronic ocular hypertension. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4337–4346. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Chang, P.; Pennesi, M.E.; Yang, Z.; Zhang, J.; Li, D.; Wu, S.M.; Gross, R.L. Effects of elevated intraocular pressure on mouse retinal ganglion cells. Vis. Res. 2005, 45, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ederra, J.; Verkman, A.S. Mouse model of sustained elevation in intraocular pressure produced by episcleral vein occlusion. Exp. Eye Res. 2006, 82, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Sawada, A.; Neufeld, A.H. Confirmation of the rat model of chronic, moderately elevated intraocular pressure. Exp. Eye Res. 1999, 69, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Tsuruga, H.; Murata, H.; Araie, M.; Aihara, M. A model for the easy assessment of pressure-dependent damage to retinal ganglion cells using cyan fluorescent protein-expressing transgenic mice. Mol. Vis. 2012, 18, 2468–2478. [Google Scholar] [PubMed]
- Urcola, J.H.; Hernández, M.; Vecino, E. Three experimental glaucoma models in rats: Comparison of the effects of intraocular pressure elevation on retinal ganglion cell size and death. Exp. Eye Res. 2006, 83, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.Y.; Kaufman, P.L.; Gabelt, B.T.; To, E.C.; Matsubara, J.A. Neurochemical correlates of cortical plasticity after unilateral elevated intraocular pressure in a primate model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2573–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.J.; Zelenak, D. Experimental glaucoma in the primate induced by latex microspheres. J. Neurosci. Methods 2001, 111, 39–48. [Google Scholar] [CrossRef]
- Huberman, A.D.; Stellwagen, D.; Chapman, B. Decoupling eye-specific segregation from lamination in the lateral geniculate nucleus. J. Neurosci. 2002, 22, 9419–9429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazawa, M.; Tomita, G.; Taniguchi, T.; Sasaoka, M.; Hara, H.; Kitazawa, Y.; Araie, M. Morphometric evaluation of changes with time in optic disc structure and thickness of retinal nerve fibre layer in chronic ocular hypertensive monkeys. Exp. Eye Res. 2006, 82, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, T.; Kawasaki, H.; Aihara, M.; Saeki, T.; Ymagishi, R.; Atarashi, T.; Mayama, C.; Araie, M. Establishment of an experimental ferret ocular hypertension model for the analysis of central visual pathway damage. Sci. Rep. 2014, 4, 6501. [Google Scholar] [CrossRef] [Green Version]
- Dunn, D.G.; Harris, R.K.; Meis, J.M.; Sweet, D.E. A histomorphologic and immunohistochemical study of chordoma in twenty ferrets (Mustela putorius furo). Vet. Pathol. 1991, 28, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, K.P.; Garipis, N.; Distler, C. Optokinetic deficits in albino ferrets (Mustela putorius furo): A behavioral and electrophysiological study. J. Neurosci. 2004, 24, 4061–4069. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, H.; Crowley, J.C.; Livesey, F.J.; Katz, L.C. Molecular organization of the ferret visual thalamus. J. Neurosci. 2004, 24, 9962–9970. [Google Scholar] [CrossRef] [Green Version]
- Iwai, L.; Ohashi, Y.; van der List, D.; Usrey, W.M.; Miyashita, Y.; Kawasaki, H. FoxP2 is a parvocellular-specific transcription factor in the visual thalamus of monkeys and ferrets. Cereb. Cortex. 2013, 23, 2204–2212. [Google Scholar] [CrossRef]
- Nigel, I.C. Neuroanatomy of the ferret brain. In Biology and Disease of the Ferret; Games, G.F., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 1998; pp. 71–102. [Google Scholar]
- Mizuguchi, K.; Horiike, T.; Matsumoto, N.; Ichikawa, Y.; Shinmyo, Y.; Kawasaki, H. Distribution and Morphological Features of Microglia in the Developing Cerebral Cortex of Gyrencephalic Mammals. Neurochem. Res. 2018, 43, 1075–1085. [Google Scholar] [CrossRef]
- Thompson, I.D.; Morgan, J.E. The development of retinal ganglion cell decussation patterns in postnatal pigmented and albino ferrets. Eur. J. Neurosci. 1993, 5, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Montiani-Ferreira, F.; Mattos, B.C.; Russ, H.H. Reference values for selected ophthalmic diagnostic tests of the ferret (Mustela putorius furo). Vet. Ophthalmol. 2006, 9, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Sapienza, J.S.; Porcher, D.; Collins, B.R.; Gum, G.G.; Brooks, D.E. Tonometry in clinically normal ferrets (Mustela putorius furo). Prog. Vet. Comp. Ophthalmol. 1991, 1, 291–294. [Google Scholar]
- Blanco, R.; Martinez-Navarrete, G.; Pérez-Rico, C.; Valiente-Soriano, F.J.; Avilés-Trigueros, M.; Vicente, J.; Fernandez, E.; Vidal-Sanz, M.; de la Villa, P. A Chronic Ocular-Hypertensive Rat Model induced by Injection of the Sclerosant Agent Polidocanol in the Aqueous Humor Outflow Pathway. Int. J. Mol. Sci. 2019, 20, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Girolamo, N.; Andreani, V.; Guandalini, A.; Selleri, P. Evaluation of intraocular pressure in conscious ferrets (Mustela putorius furo) by means of rebound tonometry and comparison with applanation tonometry. Vet. Rec. 2013, 172, 396. [Google Scholar] [CrossRef]
- Hernández-Guerra, A.M.; Rodilla, V.; López-Murcia, M.M. Ocular biometry in the adult anesthetized ferret (Mustela putorius furo). Vet. Ophthalmol. 2007, 10, 50–52. [Google Scholar] [CrossRef]
- Cucchiaro, J.B. Early development of the retinal line of decussation in normal and albino ferrets. J. Comp. Neurol. 1991, 312, 193–206. [Google Scholar] [CrossRef]
- Korth, M.J.; Jünemann, A.M.; Horn, F.K.; Bergua, A.; Cursiefen, C.; Velten, I.; Budde, W.M.; Wisse, M.; Martus, P. Synopsis of various electrophysiological tests in early glaucoma diagnosis--temporal and spatiotemporal contrast sensitivity, light- and color-contrast pattern-reversal electroretinogram, blue-yellow VEP. Klin. Monbl. Augenheilkd. 2000, 216, 360–368. [Google Scholar] [CrossRef]
- Sato, C.; Iwai-Takekoshi, L.; Ichikawa, Y.; Kawasaki, H. Cell type-specific expression of FoxP2 in the ferret and mouse retina. Neurosci. Res. 2017, 117, 1–13. [Google Scholar] [CrossRef]
- Yen, M.Y.; Cheng, C.Y.; Wang, A.G. Retinal nerve fiber layer thickness in unilateral amblyopia. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2224–2230. [Google Scholar] [CrossRef] [PubMed]
- Shimazawa, M.; Ito, Y.; Inokuchi, Y.; Yamanaka, H.; Nakanishi, T.; Hayashi, T.; Ji, B.; Higuchi, M.; Suhara, T.; Imamura, K.; et al. An alteration in the lateral geniculate nucleus of experimental glaucoma monkeys: In vivo positron emission tomography imaging of glial activation. PLoS ONE 2012, 7, e30526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, D.; Jim, J.; To, E.; Rasmussen, C.; Kaufman, P.L.; Matsubara, J. Astrocyte and microglial activation in the lateral geniculate nucleus and visual cortex of glaucomatous and optic nerve transected primates. Mol. Vis. 2009, 15, 2217–2229. [Google Scholar] [PubMed]
- Dai, Y.; Sun, X.; Yu, X.; Guo, W.; Yu, D. Astrocytic responses in the lateral geniculate nucleus of monkeys with experimental glaucoma. Vet. Ophthalmol. 2012, 15, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.A.; Knott, M.; Heidemann, R.; Michelson, G.; Kober, T.; Dörfler, A.; Engelhorn, T. Investigation of lateral geniculate nucleus volume and diffusion tensor imaging in patients with normal tension glaucoma using 7 tesla magnetic resonance imaging. PLoS ONE 2018, 13, e0198830. [Google Scholar] [CrossRef]
- Liu, M.; Guo, L.; Salt, T.E.; Cordeiro, M.F. Dendritic changes in rat visual pathway associated with experimental ocular hypertension. Curr. Eye Res. 2014, 39, 953–963. [Google Scholar] [CrossRef]
- Sapienza, A.; Raveu, A.L.; Reboussin, E.; Roubeix, C.; Boucher, C.; Dégardin, J.; Godefroy, D.; Rostène, W.; Reaux-Le Goazigo, A.; Baudouin, C.; et al. Bilateral neuroinflammatory processes in visual pathways induced by unilateral ocular hypertension in the rat. J. Neuroinflamm. 2016, 13, 44. [Google Scholar] [CrossRef]
- Gallego, B.I.; Salazar, J.J.; de Hoz, R.; Rojas, B.; Ramírez, A.I.; Salinas-Navarro, M.; Ortín-Martínez, A.; Valiente-Soriano, F.J.; Avilés-Trigueros, M.; Villegas-Perez, M.P.; et al. IOP induces upregulation of GFAP and MHC-II and microglia reactivity in mice retina contralateral to experimental glaucoma. J. Neuroinflamm. 2012, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Sam-Wah Tay, S.; Ng, Y.K. Nitric oxide, microglial activities and neuronal cell death in the lateral geniculate nucleus of glaucomatous rats. Brain Res. 2000, 878, 136–147. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Labeling Method | Red (Elevated IOP) | Green (Normal IOP) | ||||
|---|---|---|---|---|---|---|---|
| A | A1 | C | A | A1 | C | ||
| Axon | CTB | −66.79% | −71.5% | −80.9% | −25.1% | −22.1% | −26.8% |
| Neuron | NeuN | −36.6% | −34.6% | −38.4% | −6.4% | −3.5% | −6.6% |
| Astrocyte | GFAP | 42.3% | 6.4% | 30.4% | 4.9% | −3.1% | 1.6% |
| Microglia | Iba-1 | 16.2% | 22.1% | 27.5% | −3.2% | −2.3% | 3.3% |
| Target | Labeling Method | Ferret | Monkey27, 28 | Rat31, 33–36 |
|---|---|---|---|---|
| IOP (Elevated IOP duration) | 13 w | 6 m to 1 y | 14 d to 8 m | |
| Axon activity | CTB | Bilateral damage | − | − |
| Neuron | NeuN | A, A1, and C layers damage | damage | damage |
| Astrocyte | GFAP | Only A layer damage | activation | activation |
| Microglia | Iba-1 | A, A1, and C layers activation | activation | activation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujishiro, T.; Honjo, M.; Kawasaki, H.; Asaoka, R.; Yamagishi, R.; Aihara, M. Structural Changes and Astrocyte Response of the Lateral Geniculate Nucleus in a Ferret Model of Ocular Hypertension. Int. J. Mol. Sci. 2020, 21, 1339. https://doi.org/10.3390/ijms21041339
Fujishiro T, Honjo M, Kawasaki H, Asaoka R, Yamagishi R, Aihara M. Structural Changes and Astrocyte Response of the Lateral Geniculate Nucleus in a Ferret Model of Ocular Hypertension. International Journal of Molecular Sciences. 2020; 21(4):1339. https://doi.org/10.3390/ijms21041339
Chicago/Turabian StyleFujishiro, Takashi, Megumi Honjo, Hiroshi Kawasaki, Ryo Asaoka, Reiko Yamagishi, and Makoto Aihara. 2020. "Structural Changes and Astrocyte Response of the Lateral Geniculate Nucleus in a Ferret Model of Ocular Hypertension" International Journal of Molecular Sciences 21, no. 4: 1339. https://doi.org/10.3390/ijms21041339
APA StyleFujishiro, T., Honjo, M., Kawasaki, H., Asaoka, R., Yamagishi, R., & Aihara, M. (2020). Structural Changes and Astrocyte Response of the Lateral Geniculate Nucleus in a Ferret Model of Ocular Hypertension. International Journal of Molecular Sciences, 21(4), 1339. https://doi.org/10.3390/ijms21041339