Synthetic Cathinones Induce Cell Death in Dopaminergic SH-SY5Y Cells via Stimulating Mitochondrial Dysfunction



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
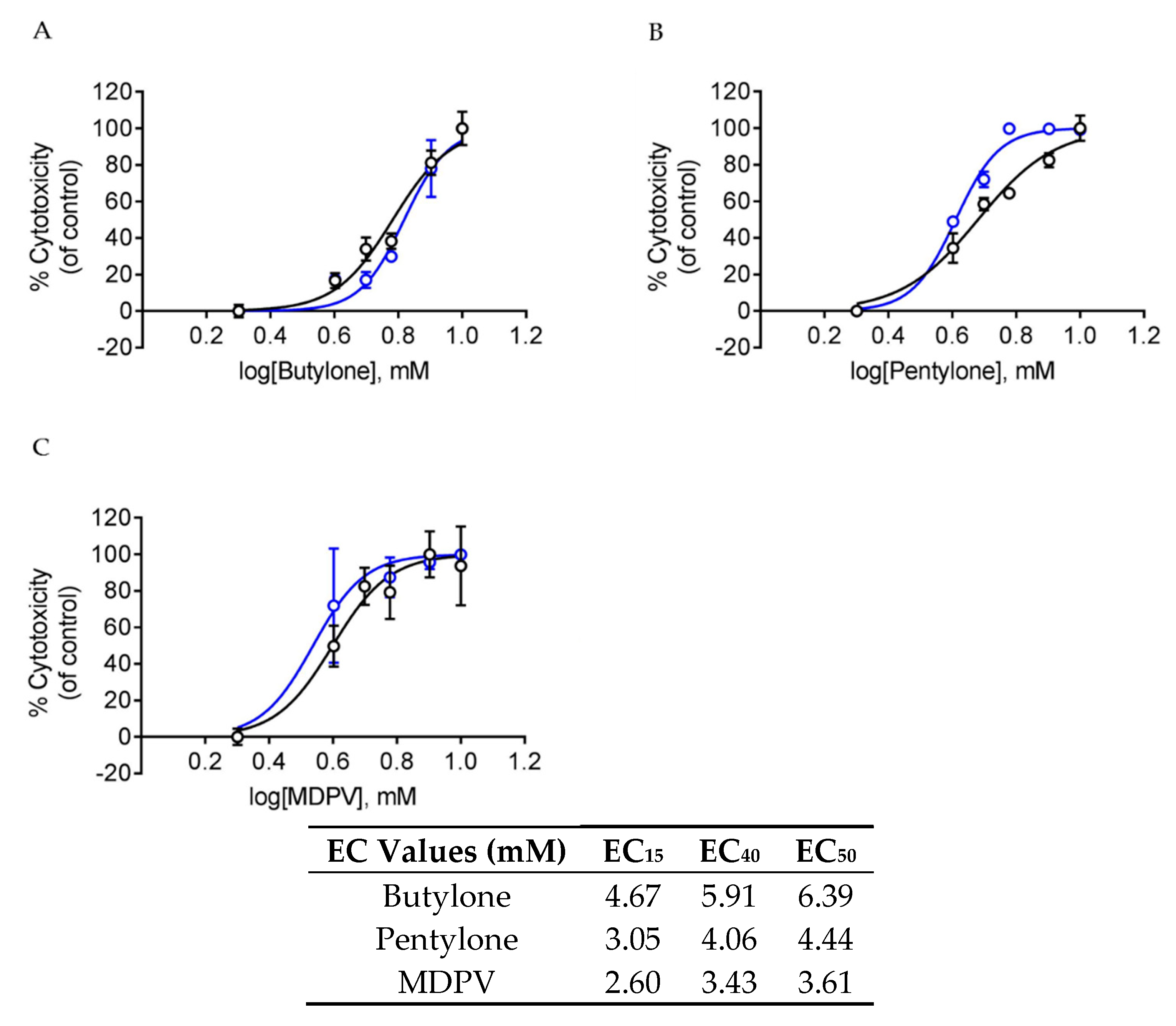
2.1. Butylone, Pentylone, and MDPV Elicited Neurotoxicity with Different Potencies
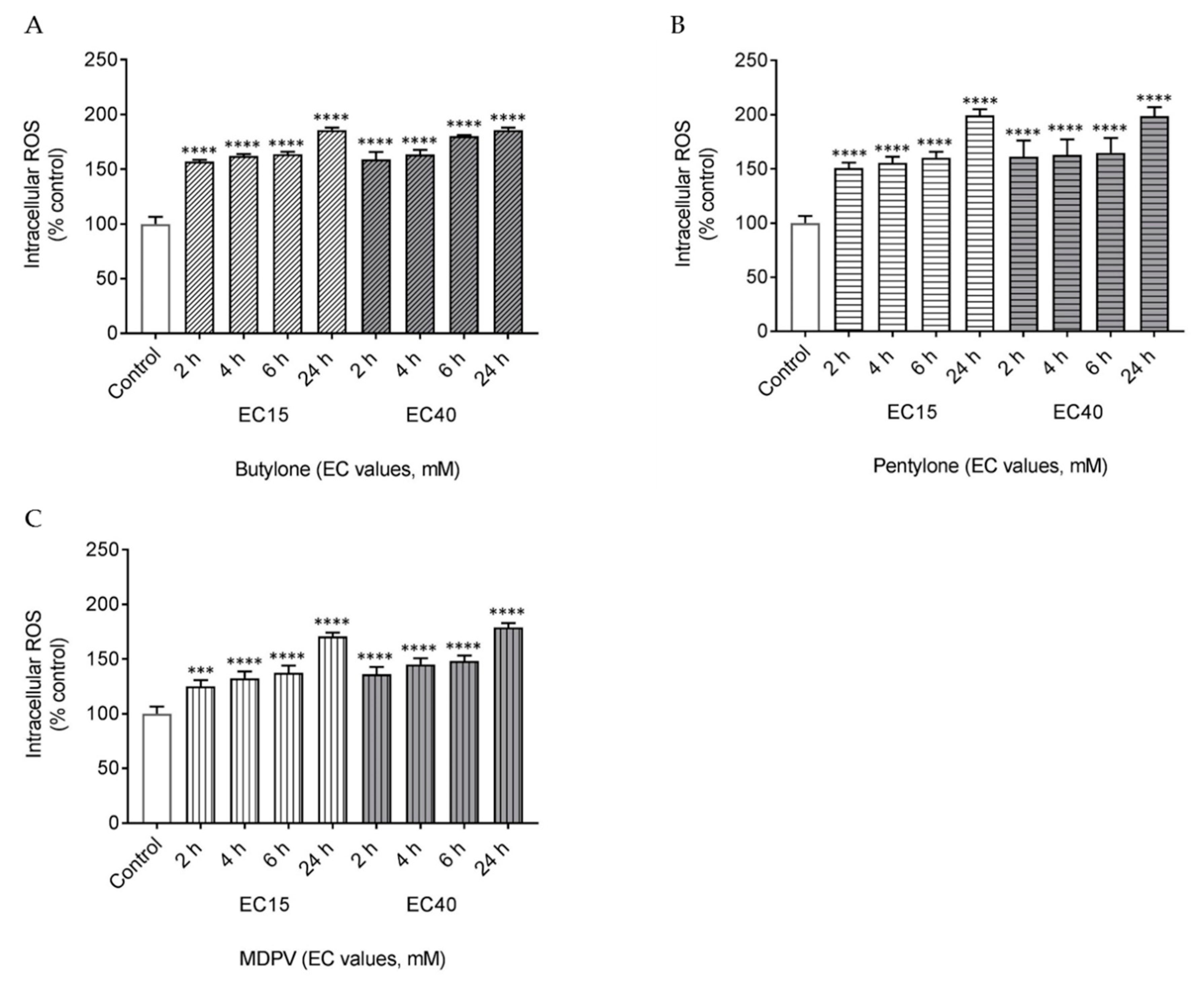
2.2. Butylone, Pentylone, and MDPV Triggered Oxidative Stress
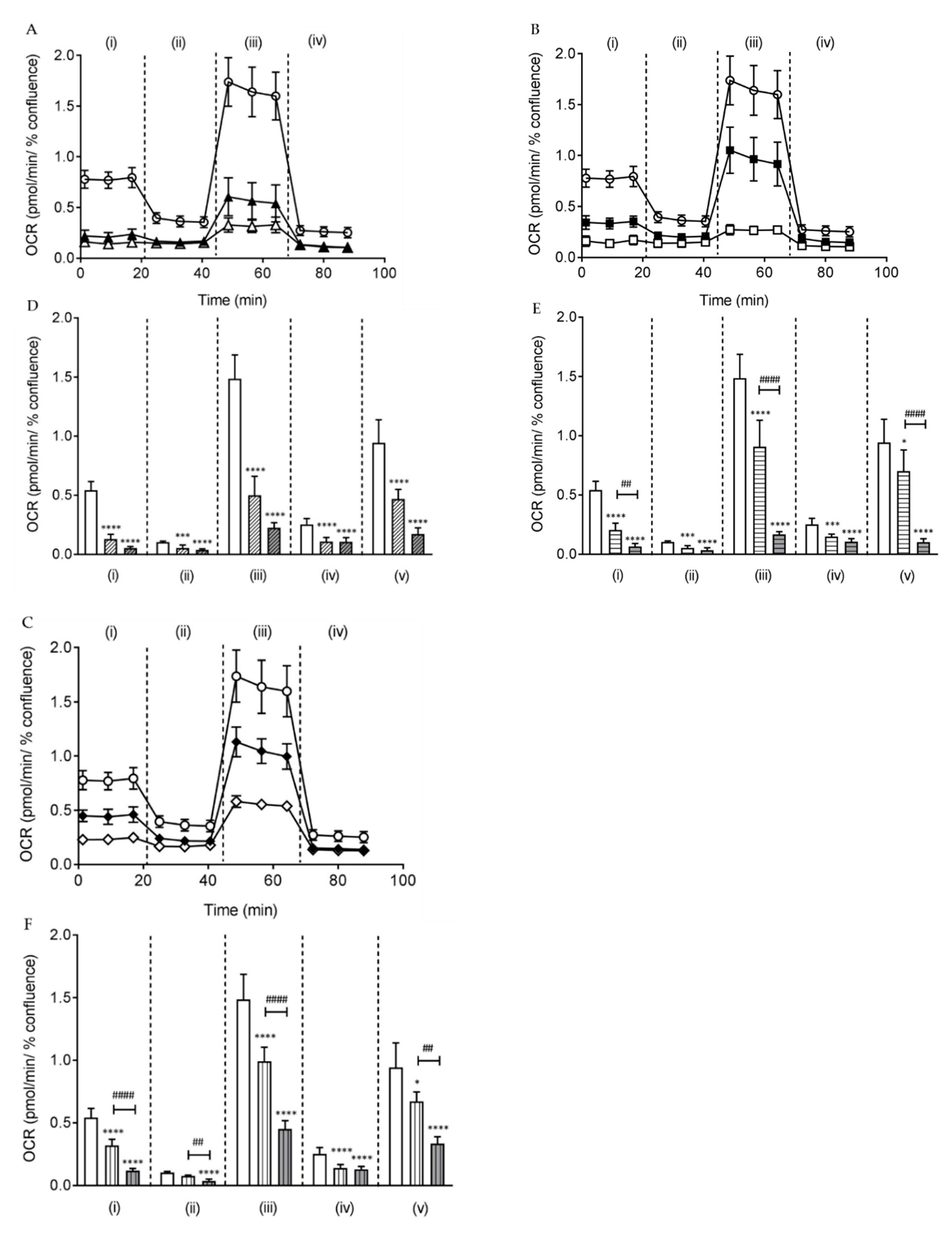
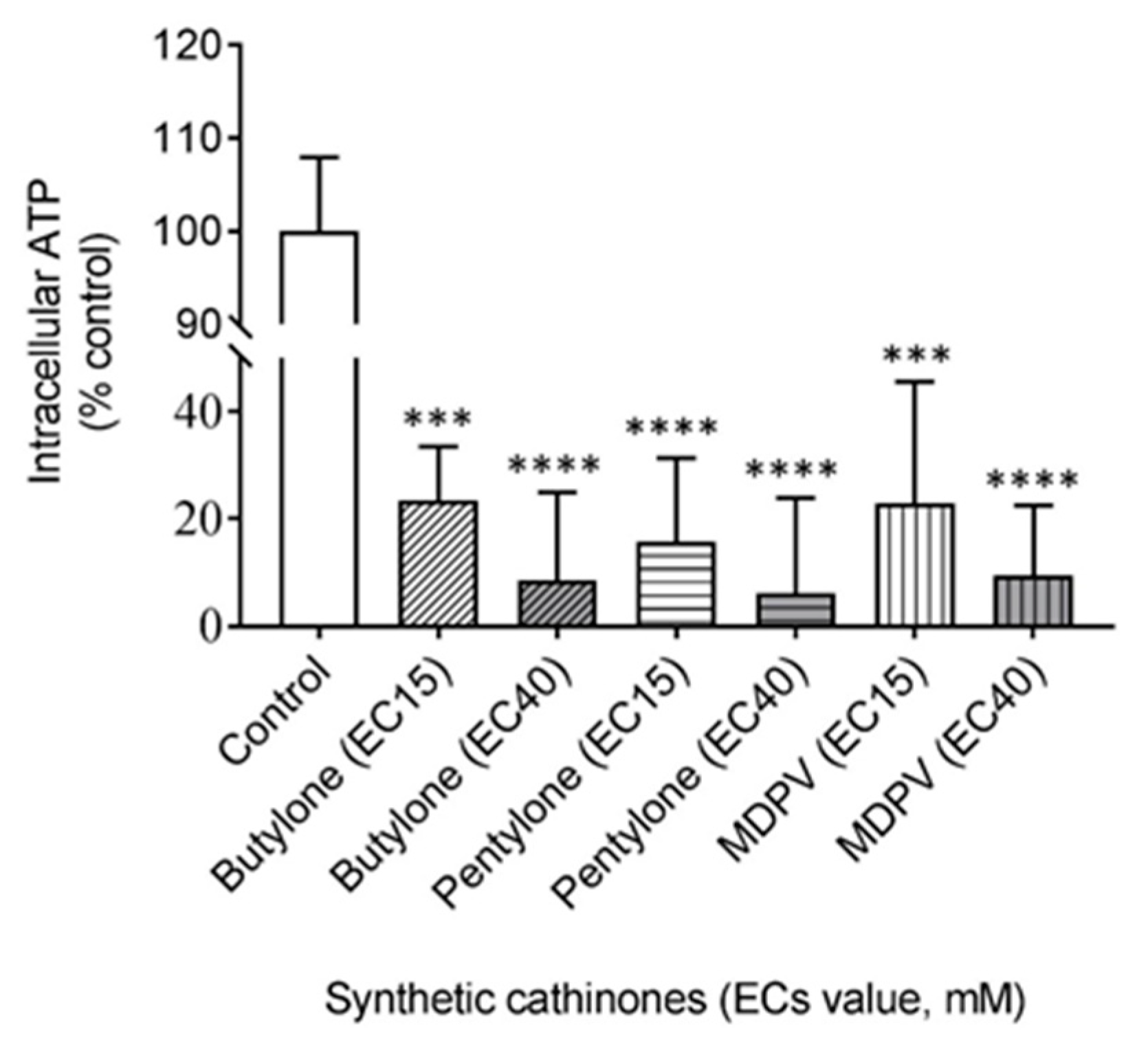
2.3. Butylone, Pentylone, and MDPV Compromised Mitochondrial Bioenergetics
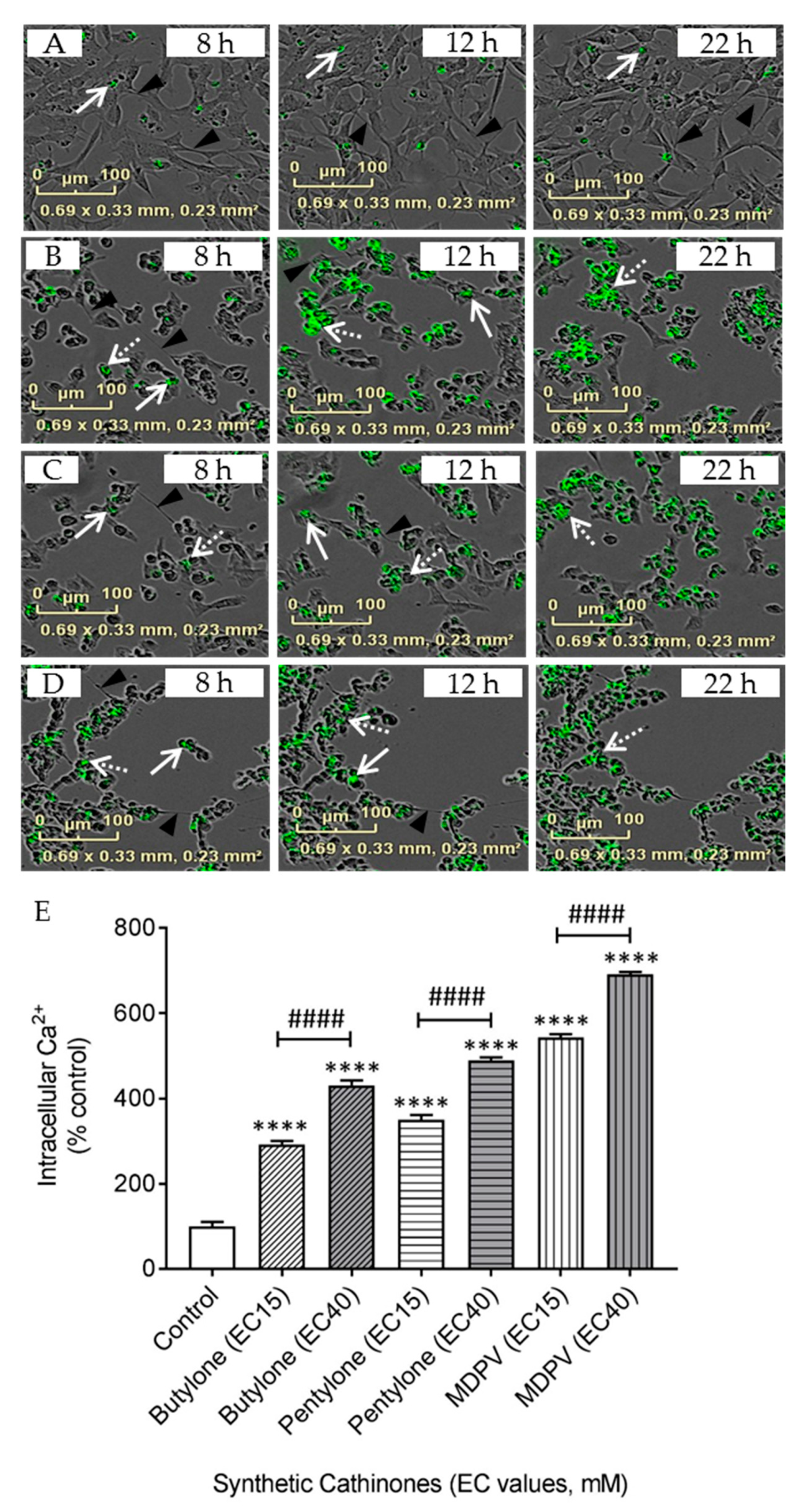
2.4. Butylone, Pentylone, and MDPV Altered Neuronal Ca2+ Homeostasis
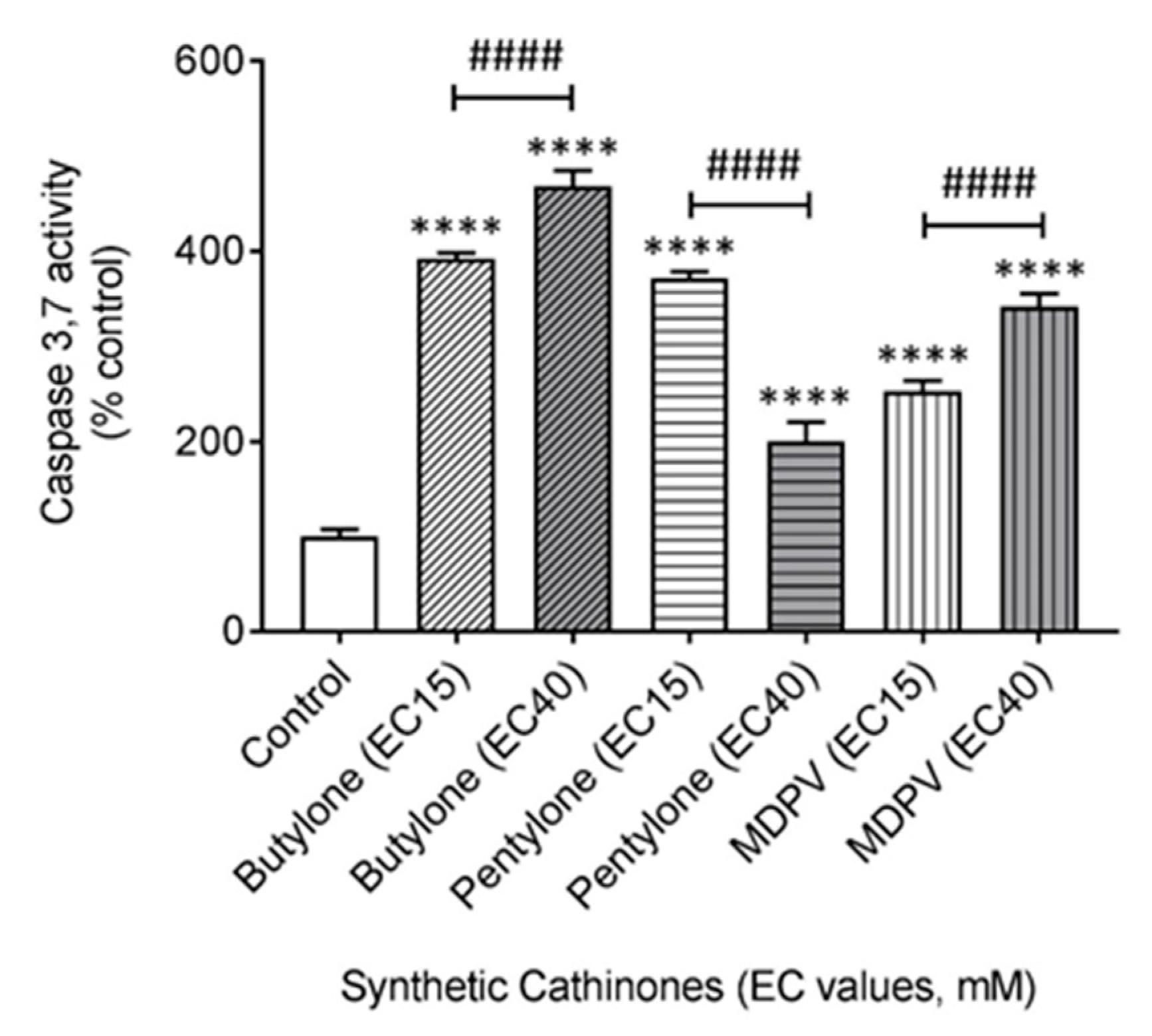
2.5. Butylone, Pentylone, and MDPV Induce an Apoptotic Cell Death Pathway
3. Discussion
4. Materials and Methods
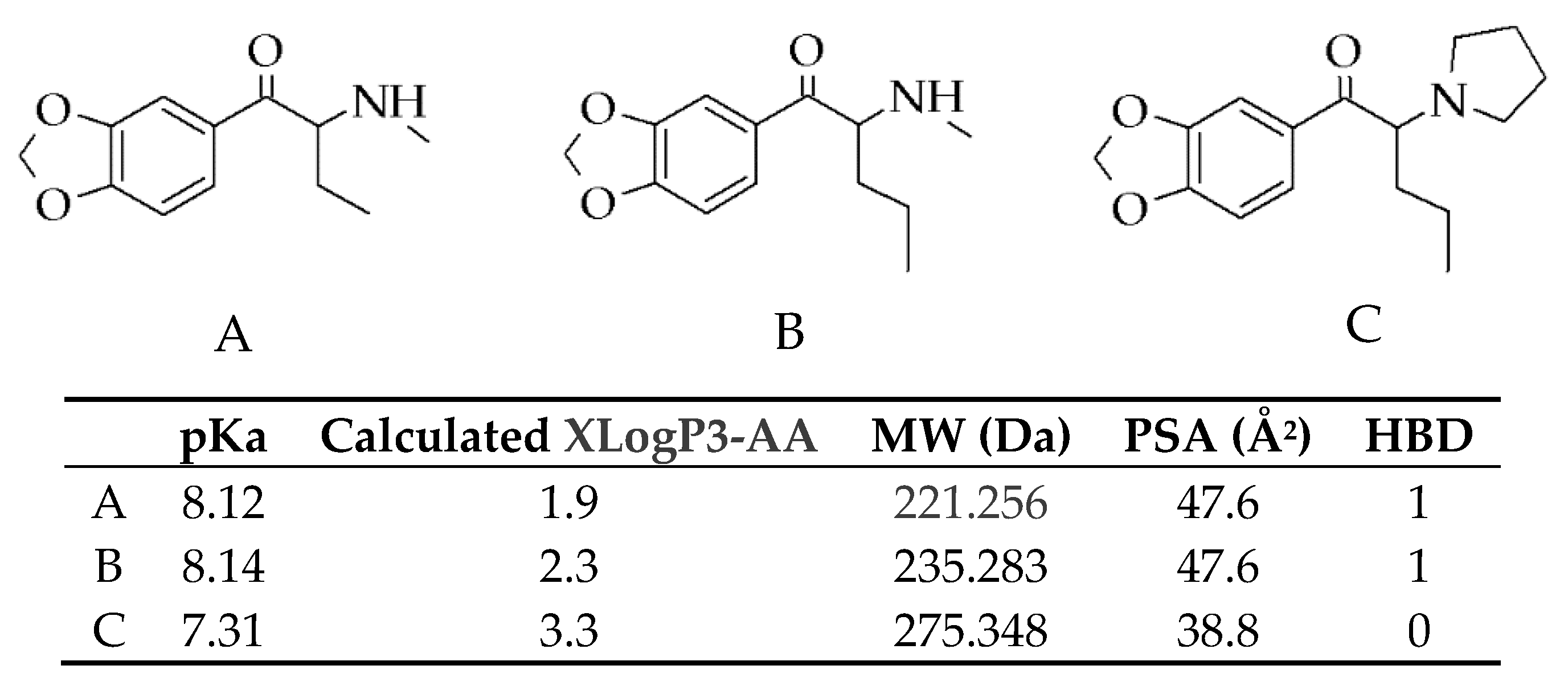
4.1. Chemical Synthesis
4.2. Cellular Studies
4.3. Cell Culture
4.4. Cytotoxicity Assays
4.5. TB Assay
4.6. LDH Assay
4.7. Measurement of Intracellular ROS
4.8. Measurement of Mitochondrial Respiration
4.9. Measurement of Intracellular Adenosine Triphosphate (ATP)
4.10. Real-Time Imaging and Measurement of Intracellular Ca2+
4.11. Measurement of Caspase 3 and 7 Activity
4.12. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- German, C.L.; Fleckenstein, A.E.; Hanson, G.R. Bath salts and synthetic cathinones: An emerging designer drug phenomenon. Life Sci. 2014, 97, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huestis, M.A.; Brandt, S.D.; Rana, S.; Auwärter, V.; Baumann, M.H. Impact of Novel Psychoactive Substances on Clinical and Forensic Toxicology and Global Public Health. Clin. Chem. 2017, 63, 1564–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannotti, G.; Canazza, I.; Caffino, L.; Bilel, S.; Ossato, A.; Fumagalli, F.; Marti, M. The Cathinones MDPV and α-PVP Elicit Different Behavioral and Molecular Effects Following Acute Exposure. Neurotox. Res. 2017, 32, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.D.; Freeman, S.; Sumnall, H.R.; Measham, F.; Cole, J. Analysis of NRG ‘legal highs’ in the UK: Identification and formation of novel cathinones. Drug Test. Anal. 2011, 3, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Leffler, A.M.; Smith, P.B.; de Armas, A.; Dorman, F.L. The analytical investigation of synthetic street drugs containing cathinone analogs. Forensic Sci. Int. 2014, 234, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Zuba, D.; Byrska, B. Prevalence and co-existence of active components of ‘legal highs’: Analysis of ‘legal highs’ composition. Drug Test. Anal. 2013, 5, 420–429. [Google Scholar] [CrossRef]
- Reitzel, L.A.; Dalsgaard, P.W.; Müller, I.B.; Cornett, C. Identification of ten new designer drugs by GC-MS, UPLC-QTOF-MS, and NMR as part of a police investigation of a Danish Internet company. Drug Test. Anal. 2012, 4, 342–354. [Google Scholar] [CrossRef]
- Westphal, F.; Junge, T.; Girreser, U.; Greibl, W.; Doering, C. Mass, NMR and IR spectroscopic characterization of pentedrone and pentylone and identification of their isocathinone by-products. Forensic Sci. Int. 2012, 217, 157–167. [Google Scholar] [CrossRef]
- Majchrzak, M.; Celiński, R.; Kuś, P.; Kowalska, T.; Sajewicz, M. The newest cathinone derivatives as designer drugs: An analytical and toxicological review. Forensic Toxicol. 2018, 36, 33–50. [Google Scholar] [CrossRef] [Green Version]
- Palamar, J.; Salomone, A.; Gerace, E.; Di Corcia, D.; Vincenti, M.; Cleland, C. Hair testing to assess both known and unknown use of drugs amongst ecstasy users in the electronic dance music scene. Int. J. Drug Policy 2017, 48, 91. [Google Scholar] [CrossRef]
- Salomone, A.; Palamar, J.J.; Gerace, E.; Di Corcia, D.; Vincenti, M. Hair Testing for Drugs of Abuse and New Psychoactive Substances in a High-Risk Population. J. Anal. Toxicol. 2017, 41, 376–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, A.L.A.; Friscia, M.; Yeakel, J.K.; Logan, B.K. Use of synthetic stimulants and hallucinogens in a cohort of electronic dance music festival attendees. Forensic Sci. Int. 2018, 282, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Saha, K.; Li, Y.; Holy, M.; Lehner, K.R.; Bukhari, M.O.; Partilla, J.S.; Sandtner, W.; Sitte, H.H.; Baumann, M.H. The synthetic cathinones, butylone and pentylone, are stimulants that act as dopamine transporter blockers but 5-HT transporter substrates. Psychopharmacology 2019, 236, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Karila, L.; Lafaye, G.; Scocard, A.; Cottencin, O.; Benyamina, A. MDPV and α-PVP use in humans: The twisted sisters. Neuropharmacology 2018, 134, 65–72. [Google Scholar] [CrossRef]
- Dolan, S.B.; Chen, Z.; Huang, R.; Gatch, M.B. “Ecstasy” to addiction: Mechanisms and reinforcing effects of three synthetic cathinone analogs of MDMA. Neuropharmacology 2018, 133, 171–180. [Google Scholar] [CrossRef]
- Simmons, S.J.; Gregg, R.A.; Tran, F.H.; Mo, L.; Weltin, E.; Barker, D.J.; Gentile, T.A.; Watterson, L.R.; Rawls, S.M.; Muschamp, J.W. Comparing rewarding and reinforcing properties between ‘bath salt’ 3,4-methylenedioxypyrovalerone (MDPV) and cocaine using ultrasonic vocalizations in rats. Addict. Biol. 2018, 23, 102–110. [Google Scholar] [CrossRef]
- Zawilska, J.B.; Wojcieszak, J. Novel Psychoactive Substances: Classification and General Information. In Synthetic Cathinones: Novel Addictive and Stimulatory Psychoactive Substances; Zawilska, J.B., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 11–24. [Google Scholar] [CrossRef]
- Weinstein, A.M.; Rosca, P.; Fattore, L.; London, E.D. Synthetic Cathinone and Cannabinoid Designer Drugs Pose a Major Risk for Public Health. Front. Psychiatr. 2017, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Elliott, S.; Evans, J. A 3-year review of new psychoactive substances in casework. Forensic Sci. Int. 2014, 243, 55–60. [Google Scholar] [CrossRef]
- Rojek, S.; Klys, M.; Strona, M.; Maciow, M.; Kula, K. “Legal highs”-Toxicity in the clinical and medico-legal aspect as exemplified by suicide with bk-MBDB administration. Forensic Sci. Int. 2012, 222, E1–E6. [Google Scholar] [CrossRef]
- Marinetti, L.J.; Antonides, H.M. Analysis of Synthetic Cathinones Commonly Found in Bath Salts in Human Performance and Postmortem Toxicology: Method Development, Drug Distribution and Interpretation of Results. J. Anal. Toxicol. 2013, 37, 135–146. [Google Scholar] [CrossRef]
- Kesha, K.; Boggs, C.L.; Ripple, M.G.; Allan, C.H.; Levine, B.; Jufer-Phipps, R.; Doyon, S.; Chi, P.; Fowler, D.R. Methylenedioxypyrovalerone (“Bath Salts”), Related Death: Case Report and Review of the Literature. J. Forensic Sci. 2013, 58, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Kriikku, P.; Wilhelm, L.; Schwarz, O.; Rintatalo, J. New designer drug of abuse: 3,4-Methylenedioxypyrovalerone (MDPV). Findings from apprehended drivers in Finland. Forensic Sci. Int. 2011, 210, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Froberg, B.A.; Levine, M.; Beuhler, M.C.; Judge, B.S.; Moore, P.W.; Engebretsen, K.M.; McKeown, N.J.; Rosenbaum, C.D.; Young, A.C.; Rusyniak, D.E.; et al. Acute Methylenedioxypyrovalerone Toxicity. J. Med. Toxicol. 2015, 11, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krotulski, A.J.; Papsun, D.M.; De Martinis, B.S.; Mohr, A.L.A.; Logan, B.K. N-Ethyl Pentylone (Ephylone) Intoxications: Quantitative Confirmation and Metabolite Identification in Authentic Human Biological Specimens. J. Anal. Toxicol. 2018, 42, 467–475. [Google Scholar] [CrossRef]
- Assi, S.; Gulyamova, N.; Kneller, P.; Osselton, D. The effects and toxicity of cathinones from the users’ perspectives: A qualitative study. Hum. Psychopharmacol. Clin. Exp. 2017, 32, e2610. [Google Scholar] [CrossRef]
- Archer, T.; Kostrzewa, R.M. Synthetic Cathinones: Neurotoxic Health Hazards and Potential for Abuse. In Synthetic Cathinones: Novel Addictive and Stimulatory Psychoactive Substances; Zawilska, J.B., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–10. [Google Scholar] [CrossRef]
- Leyrer-Jackson, J.M.; Nagy, E.K.; Olive, M.F. Cognitive Deficits and Neurotoxicity Induced by Synthetic Cathinones: Is There a Role for Neuroinflammation? Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–17. [Google Scholar] [CrossRef]
- Costa, G.; De Luca, M.A.; Piras, G.; Marongiu, J.; Fattore, L.; Simola, N. Neuronal and peripheral damages induced by synthetic psychoactive substances: An update of recent findings from human and animal studies. Neural Regen. Res. 2020, 15, 802–816. [Google Scholar] [CrossRef]
- Ádám, Á.; Gerecsei, L.I.; Lepesi, N.; Csillag, A. Apoptotic effects of the ‘designer drug’ methylenedioxypyrovalerone (MDPV) on the neonatal mouse brain. Neurotoxicology 2014, 44, 231–236. [Google Scholar] [CrossRef]
- Naseri, G.; Fazel, A.; Golalipour, M.J.; Haghir, H.; Sadeghian, H.; Mojarrad, M.; Hosseini, M.; Shahrokhi Sabzevar, S.; Beheshti, F.; Ghorbani, A. Exposure to mephedrone during gestation increases the risk of stillbirth and induces hippocampal neurotoxicity in mice offspring. Neurotoxicol. Teratol. 2018, 67, 10–17. [Google Scholar] [CrossRef]
- Sewalia, K.; Watterson, L.R.; Hryciw, A.; Belloc, A.; Ortiz, J.B.; Olive, M.F. Neurocognitive dysfunction following repeated binge-like self-administration of the synthetic cathinone 3,4-methylenedioxypyrovalerone (MDPV). Neuropharmacology 2018, 134, 36–45. [Google Scholar] [CrossRef]
- Wojcieszak, J.; Andrzejczak, D.; Woldan-Tambor, A.; Zawilska, J.B. Cytotoxic Activity of Pyrovalerone Derivatives, an Emerging Group of Psychostimulant Designer Cathinones. Neurotox. Res. 2016, 30, 239–250. [Google Scholar] [CrossRef]
- Den Hollander, B.; Sundström, M.; Pelander, A.; Ojanperä, I.; Mervaala, E.; Korpi, E.R.; Kankuri, E. Keto amphetamine toxicity-focus on the redox reactivity of the cathinone designer drug mephedrone. Toxicol. Sci. 2014, 141, 120–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siedlecka-Kroplewska, K.; Szczerba, A.; Lipinska, A.; Slebioda, T.; Kmiec, Z. 3-Fluoromethcathinone, a structural analog of mephedrone, inhibits growth and induces cell cycle arrest in HT22 mouse hippocampal cells. J. Physiol. Pharmacol. 2014, 65, 241–246. [Google Scholar] [PubMed]
- Valente, M.J.; Bastos, M.D.L.; Fernandes, E.; Carvalho, F.; Guedes De Pinho, P.; Carvalho, M. Neurotoxicity of β-Keto Amphetamines: Deathly Mechanisms Elicited by Methylone and MDPV in Human Dopaminergic SH-SY5Y Cells. ACS Chem. Neurosci. 2017, 8, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.; Morikawa, Y.; Kamata, K.; Shibata, A.; Miyazono, H.; Sasajima, Y.; Suenami, K.; Sato, K.; Takekoshi, Y.; Endo, S.; et al. α-Pyrrolidinononanophenone provokes apoptosis of neuronal cells through alterations in antioxidant properties. Toxicology 2017, 386, 93–102. [Google Scholar] [CrossRef]
- Soares, J.; Costa, V.M.; Bronze, S.; Gaspar, H.; Santos, S.; Bastos, M.L.; Carvalho, F.; Capela, J. Neurotoxicity of synthetic cathinones on differentiated SH-SY5Y human dopaminergic cells. Toxicol. Lett. 2016, 258, S181. [Google Scholar] [CrossRef]
- Den Hollander, B.; Sundström, M.; Pelander, A.; Siltanen, A.; Ojanperä, I.; Mervaala, E.; Korpi, E.R.; Kankuri, E. Mitochondrial respiratory dysfunction due to the conversion of substituted cathinones to methylbenzamides in SH-SY5Y cells. Sci. Rep. 2015, 5, 14924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siedlecka-Kroplewska, K.; Wrońska, A.; Stasiłojć, G.; Kmieć, Z. The Designer Drug 3-Fluoromethcathinone Induces Oxidative Stress and Activates Autophagy in HT22 Neuronal Cells. Neurotox. Res. 2018, 34, 388–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presgraves, S.P.; Ahmed, T.; Borwege, S.; Joyce, J.N. Terminally differentiated SH-SY5Y cells provide a model system for studying neuroprotective effects of dopamine agonists. Neurotox. Res. 2004, 5, 579. [Google Scholar] [CrossRef] [PubMed]
- Moszczynska, A.; Callan, S.P. Molecular, Behavioral, and Physiological Consequences of Methamphetamine Neurotoxicity: Implications for Treatment. J. Pharmacol. Exp. Ther. 2017, 362, 474–488. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, T.; Morikawa, Y.; Tanigawa, M.; Kamata, K.; Shibata, A.; Sasajima, Y.; Suenami, K.; Sato, K.; Takekoshi, Y.; Endo, S.; et al. Structure-activity relationship for toxicity of α-pyrrolidinophenones in human aortic endothelial cells. Forensic Toxicol. 2017, 35, 309–316. [Google Scholar] [CrossRef]
- Wojcieszak, J.; Andrzejczak, D.; Kedzierska, M.; Milowska, K.; Zawilska, J.B. Cytotoxicity of α-Pyrrolidinophenones: An Impact of α-Aliphatic Side-chain Length and Changes in the Plasma Membrane Fluidity. Neurotox. Res. 2018, 34, 613–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Waterbeemd, H.; Camenisch, G.; Folkers, G.; Chretien, J.R.; Raevsky, O.A. Estimation of Blood-Brain Barrier Crossing of Drugs Using Molecular Size and Shape, and H-Bonding Descriptors. J. Drug Target. 1998, 6, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, S.A.; Pennington, L.D. Structure-brain exposure relationships. J. Med. Chem. 2006, 49, 7559–7583. [Google Scholar] [CrossRef] [PubMed]
- Simmler, L.D.; Buser, T.A.; Donzelli, M.; Schramm, Y.; Dieu, L.H.; Huwyler, J.; Chaboz, S.; Hoener, M.C.; Liechti, M.E. Pharmacological characterization of designer cathinones in vitro. Br. J. Pharmacol. 2013, 168, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Simmler, L.D.; Rickli, A.; Hoener, M.C.; Liechti, M.E. Monoamine transporter and receptor interaction profiles of a new series of designer cathinones. Neuropharmacology 2014, 79, 152–160. [Google Scholar] [CrossRef]
- Valente, M.J.; Amaral, C.; Correia-da-Silva, G.; Duarte, J.A.; Bastos, M.D.L.; Carvalho, F.; Guedes de Pinho, P.; Carvalho, M. Methylone and MDPV activate autophagy in human dopaminergic SH-SY5Y cells: A new insight into the context of β-keto amphetamines-related neurotoxicity. Arch. Toxicol. 2017, 91, 3663–3676. [Google Scholar] [CrossRef]
- Soares, J.; Costa, V.; Bronze, S.; Gaspar, H.; Santos, S.; Bastos, M.D.L.; Carvalho, F.; Capela, J. Clorgyline and N-acetyl-L-cysteine provide partial protection against the toxicity of synthetic cathinones and methamphetamine on SH-SY5Y humans cells. Toxicol. Lett. 2018, 295, S274. [Google Scholar] [CrossRef]
- Barbosa, D.J.; Capela, J.P.; Feio-Azevedo, R.; Teixeira-Gomes, A.; Bastos, M.d.L.; Carvalho, F. Mitochondria: Key players in the neurotoxic effects of amphetamines. Arch. Toxicol. 2015, 89, 1695–1725. [Google Scholar] [CrossRef]
- Luethi, D.; Liechti, M.E.; Krahenbuhl, S. Mechanisms of hepatocellular toxicity associated with new psychoactive synthetic cathinones. Toxicology 2017, 387, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Valente, M.J.; Araújo, A.M.; Silva, R.; Bastos, M.d.L.; Carvalho, F.; Guedes de Pinho, P.; Carvalho, M. 3,4-Methylenedioxypyrovalerone (MDPV): In vitro mechanisms of hepatotoxicity under normothermic and hyperthermic conditions. Arch. Toxicol. 2016, 90, 1959–1973. [Google Scholar] [CrossRef]
- Chen, X.; Xing, J.; Jiang, L.; Qian, W.; Wang, Y.; Sun, H.; Wang, Y.; Xiao, H.; Wang, J.; Zhang, J. Involvement of calcium/calmodulin-dependent protein kinase II in methamphetamine-induced neural damage: Methamphetamine induces neural damage through Ca2+signaling. J. Appl. Toxicol. 2016, 36, 1460–1467. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.; Costa, V.; Gaspar, H.; Santos, S.; Bastos, M.; Carvalho, F.; Capela, J. Structure-cytotoxicity relationship profile of 13 synthetic cathinones in differentiated human SH-SY5Y neuronal cells. Neurotoxicology 2019, 75, 158. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Hernandez, H.; Cuevas, E.; Lantz, S.M.; Imam, S.Z.; Rice, K.C.; Gannon, B.M.; Fantegrossi, W.E.; Paule, M.G.; Ali, S.F. 3,4-methylenedioxypyrovalerone (MDPV) induces cytotoxic effects on human dopaminergic SH-SY5Y cells. J. Drug Alcohol Res. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- García-Repetto, R.; Moreno, E.; Soriano, T.; Jurado, C.; Giménez, M.P.; Menéndez, M. Tissue concentrations of MDMA and its metabolite MDA in three fatal cases of overdose. Forensic Sci. Int. 2003, 135, 110–114. [Google Scholar] [CrossRef]
- Glicksberg, L.; Winecker, R.; Miller, C.; Kerrigan, S. Postmortem distribution and redistribution of synthetic cathinones. Forensic Toxicol. 2018, 36, 291–303. [Google Scholar] [CrossRef]
- Coccini, T.; Vecchio, S.; Crevani, M.; Simone, U.D. Cytotoxic Effects of 3,4-Catechol-PV (One Major MDPV Metabolite) on Human Dopaminergic SH-SY5Y Cells. Neurotox. Res. 2018, 35, 49–62. [Google Scholar] [CrossRef]
- Zhao, F.; Wu, T.; Lau, A.; Jiang, T.; Huang, Z.; Wang, X.-J.; Chen, W.; Wong, P.K.; Zhang, D.D. Nrf2 promotes neuronal cell differentiation. Free Radic. Biol. Med. 2009, 47, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Colon-Perez, L.M.; Tran, K.; Thompson, K.; Pace, M.C.; Blum, K.; Goldberger, B.A.; Gold, M.S.; Bruijnzeel, A.W.; Setlow, B.; Febo, M. The psychoactive designer drug and bath salt constituent MDPV causes widespread disruption of brain functional connectivity. Neuropsychopharmacology 2016, 41, 2352–2365. [Google Scholar] [CrossRef] [Green Version]
- Bernheim, A.; See, R.E.; Reichel, C.M. Chronic methamphetamine self-administration disrupts cortical control of cognition. Neurosci. Biobehav. Rev. 2016, 69, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Curtin, K.; Fleckenstein, A.E.; Robison, R.J.; Crookston, M.J.; Smith, K.R.; Hanson, G.R. Methamphetamine/amphetamine abuse and risk of Parkinson's disease in Utah: A population-based assessment. Drug Alcohol Depend. 2014, 146, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Granado, N.; Ares-Santos, S.; Moratalla, R. Methamphetamine and parkinson’s disease. Parkinson’s Dis. 2013, 2013, 308052. [Google Scholar] [CrossRef] [PubMed]
- Kish, S.J.; Boileau, I.; Callaghan, R.C.; Tong, J.; Bolam, P. Brain dopamine neurone ‘damage’: Methamphetamine users vs. Parkinson’s disease—A critical assessment of the evidence. Eur. J. Neurosci. 2017, 45, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.E.; Bunting, A.M.; Staton, M.; Walker, R.; Shalash, S.; Winston, E.; Pangburn, K. Examination of Synthetic Cannabinoid and Cathinone Use among a Drug-Using Offender Sample, 2013–2015. J. Psychoact. Drugs 2017, 49, 436–445. [Google Scholar] [CrossRef] [PubMed]
- AIHW. National Drug Strategy Household Survey, Detailed Findings; Australian Institute of Health and Welfare, Australian’s Health: Canberra, Australia, 2017.
- Toborek, M.; Seelbach, M.J.; Rashid, C.S.; András, I.E.; Chen, L.; Park, M.; Esser, K.A. Voluntary exercise protects against methamphetamine-induced oxidative stress in brain microvasculature and disruption of the blood-brain barrier. Mol. Neurodegener. 2013, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cichello, S. Herbal treatment for hepatotoxicity associated with high fat diet, methamphetamine use and anxiety: A case study. Aust. J. Herbal Med. 2013, 25, 202–204. [Google Scholar]
- Pourmand, A.; Mazer-Amirshahi, M.; Chistov, S.; Li, A.; Park, M. Designer drugs: Review and implications for emergency management. Hum. Exp. Toxicol. 2018, 37, 94–101. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Y.; Li, Q.Y.; Zhong, Y.X.; Chen, L.P.; Du, Y.J.; He, J.; Liao, L.S.; Xiong, K.; Yi, C.X.; et al. The Main Molecular Mechanisms Underlying Methamphetamine-Induced Neurotoxicity and Implications for Pharmacological Treatment. Front. Mol. Neurosci. 2018, 11, 186. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. Mitochondrial Mechanisms of Neuronal Cell Death: Potential Therapeutics. Ann. Rev. Pharmacol. Toxicol. 2017, 57, 437–454. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Liang, M.-Z.; Chen, L. Current progress of mitochondrial transplantation that promotes neuronal regeneration. Trans. Neurodegener. 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Zündorf, G.; Reiser, G. Calcium Dysregulation and Homeostasis of Neural Calcium in the Molecular Mechanisms of Neurodegenerative Diseases Provide Multiple Targets for Neuroprotection. Antiox. Redox Signal. 2011, 14, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, D.J.; Capela, J.P.; Silva, R.; Vilas-Boas, V.; Ferreira, L.M.; Branco, P.S.; Fernandes, E.; de Lourdes Bastos, M.; Carvalho, F. The mixture of “ecstasy” and its metabolites is toxic to human SH-SY5Y differentiated cells at in vivo relevant concentrations. Arch. Toxicol. 2014, 88, 455. [Google Scholar] [CrossRef] [PubMed]
- Lantz, S.M.; Rosas-Hernandez, H.; Cuevas, E.; Robinson, B.; Rice, K.C.; Fantegrossi, W.E.; Imam, S.Z.; Paule, M.G.; Ali, S.F. Monoaminergic toxicity induced by cathinone phthalimide: An in vitro study. Neurosci. Lett. 2017, 655, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Hernandez, H.; Cuevas, E.; Lantz, S.M.; Rice, K.C.; Gannon, B.M.; Fantegrossi, W.E.; Gonzalez, C.; Paule, M.G.; Ali, S.F. Methamphetamine, 3,4-methylenedioxymethamphetamine (MDMA) and 3,4-methylenedioxypyrovalerone (MDPV) induce differential cytotoxic effects in bovine brain microvessel endothelial cells. Neurosci. Lett. 2016, 629, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leong, H.S.; Philp, M.; Simone, M.; Witting, P.K.; Fu, S. Synthetic Cathinones Induce Cell Death in Dopaminergic SH-SY5Y Cells via Stimulating Mitochondrial Dysfunction. Int. J. Mol. Sci. 2020, 21, 1370. https://doi.org/10.3390/ijms21041370
Leong HS, Philp M, Simone M, Witting PK, Fu S. Synthetic Cathinones Induce Cell Death in Dopaminergic SH-SY5Y Cells via Stimulating Mitochondrial Dysfunction. International Journal of Molecular Sciences. 2020; 21(4):1370. https://doi.org/10.3390/ijms21041370
Chicago/Turabian StyleLeong, Huey Sze, Morgan Philp, Martin Simone, Paul Kenneth Witting, and Shanlin Fu. 2020. "Synthetic Cathinones Induce Cell Death in Dopaminergic SH-SY5Y Cells via Stimulating Mitochondrial Dysfunction" International Journal of Molecular Sciences 21, no. 4: 1370. https://doi.org/10.3390/ijms21041370
APA StyleLeong, H. S., Philp, M., Simone, M., Witting, P. K., & Fu, S. (2020). Synthetic Cathinones Induce Cell Death in Dopaminergic SH-SY5Y Cells via Stimulating Mitochondrial Dysfunction. International Journal of Molecular Sciences, 21(4), 1370. https://doi.org/10.3390/ijms21041370