Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure





Abstract
:1. Introduction
2. Results
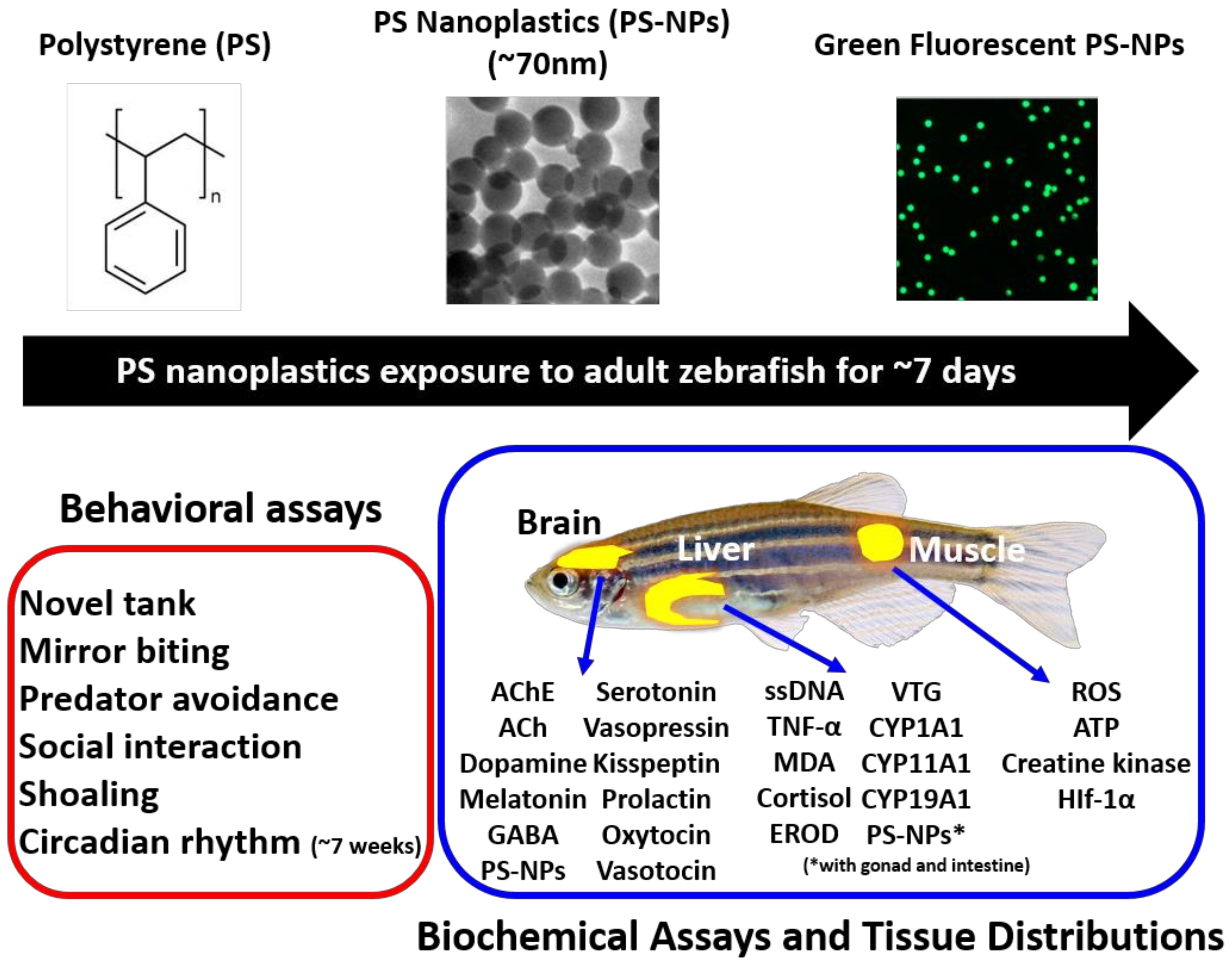
2.1. Overview of Experimental Design
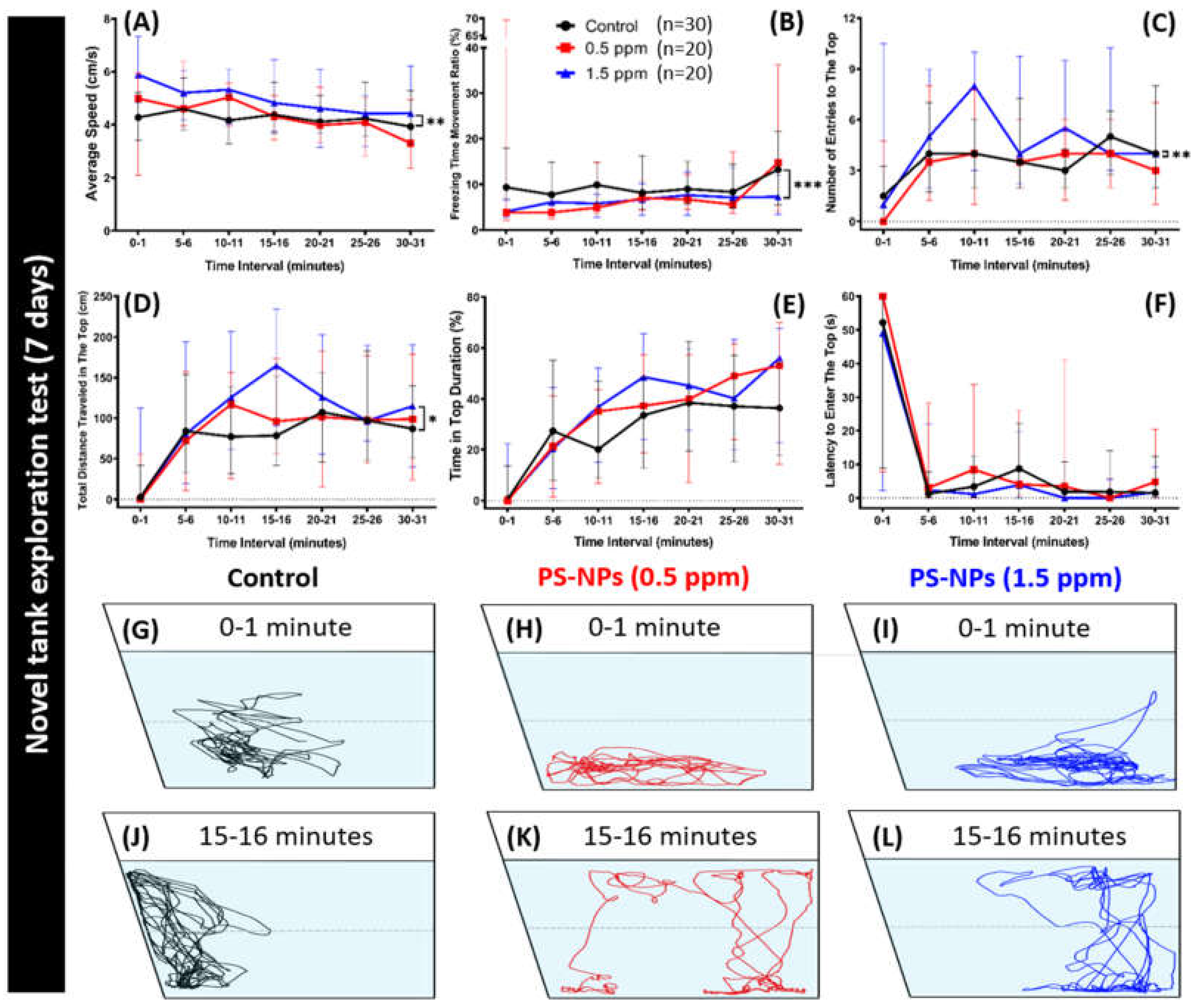
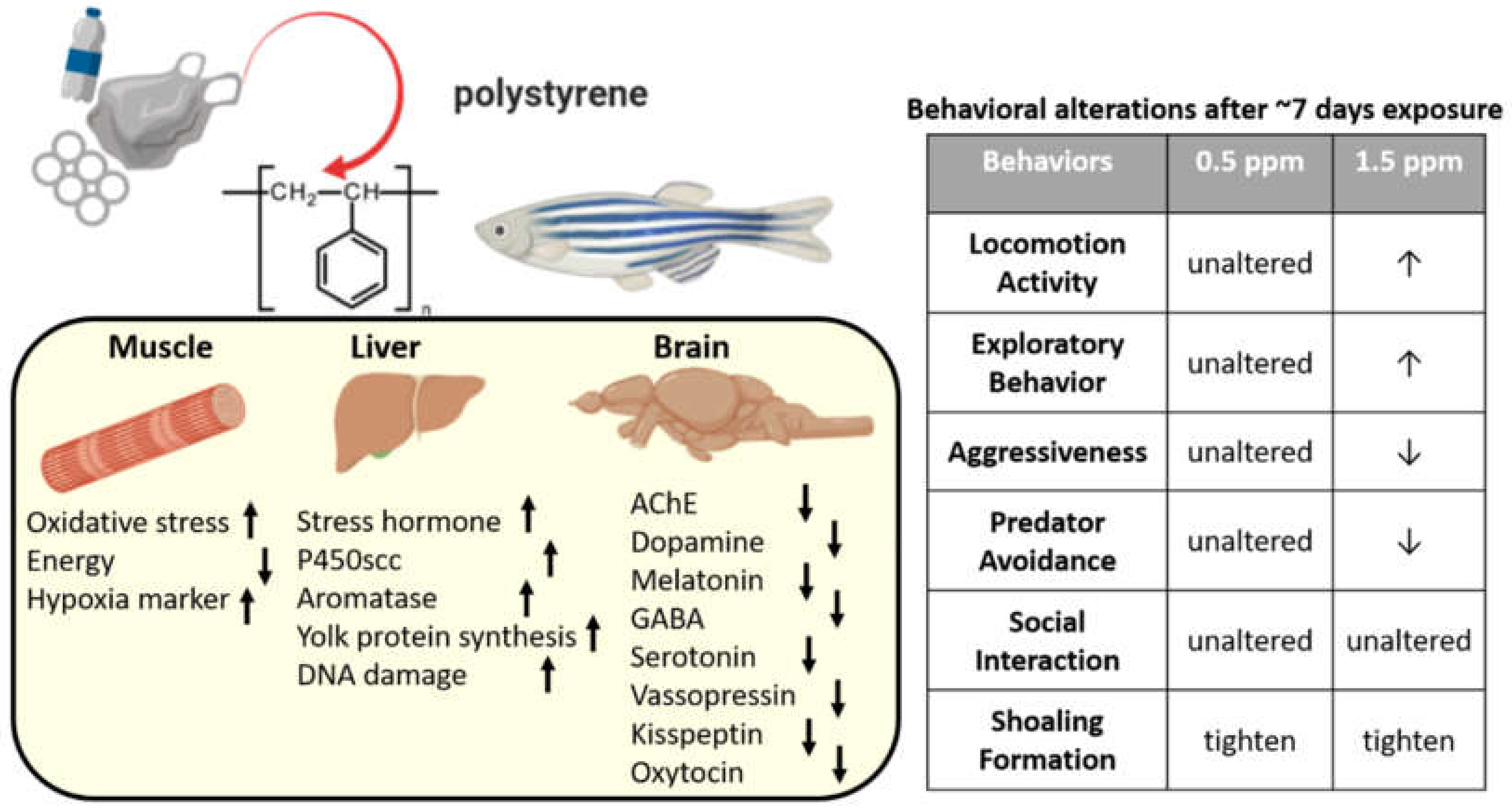
2.2. PS-NPs Exposure Reduced Average Speed and Exploration Behavior in Zebrafish
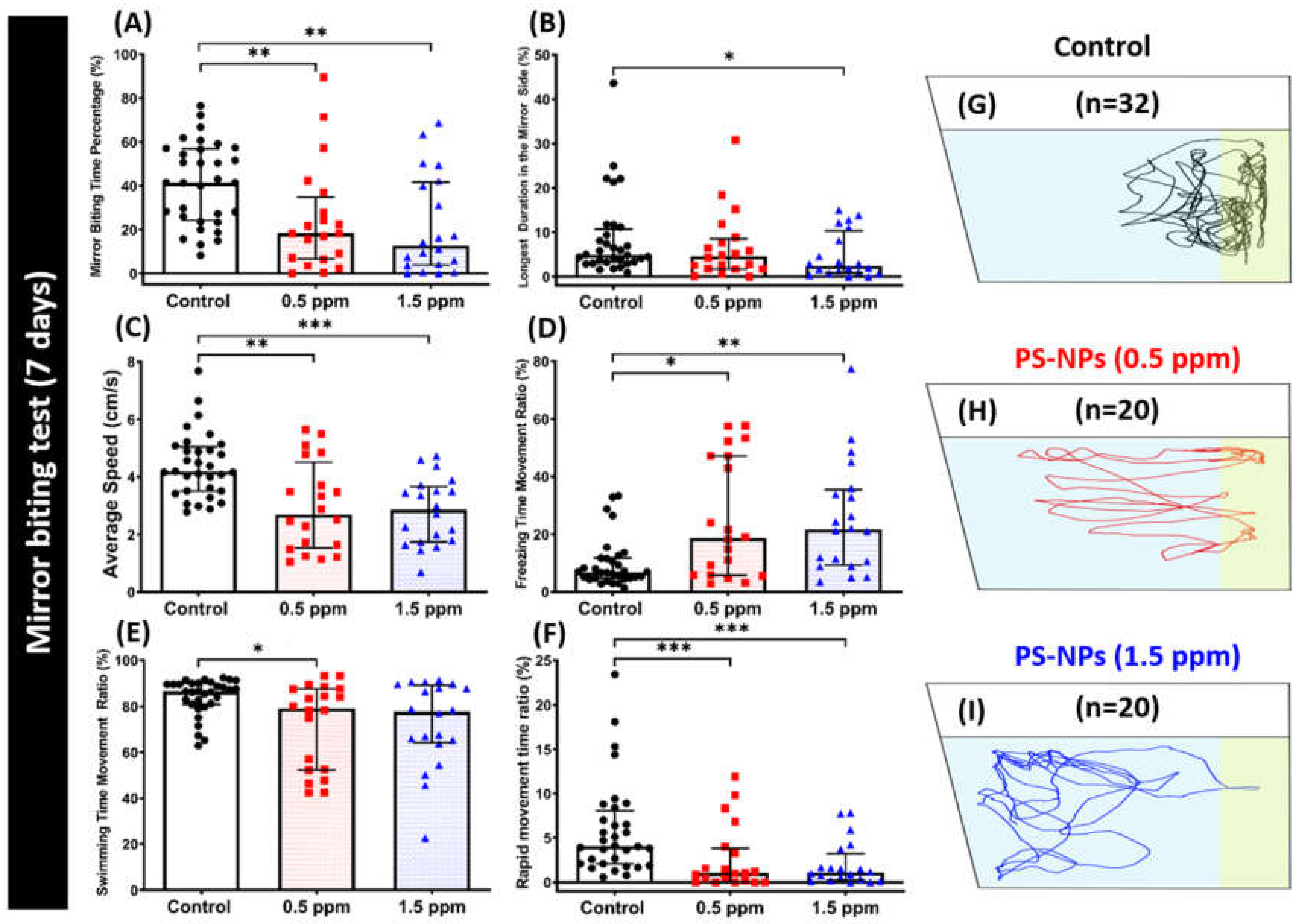
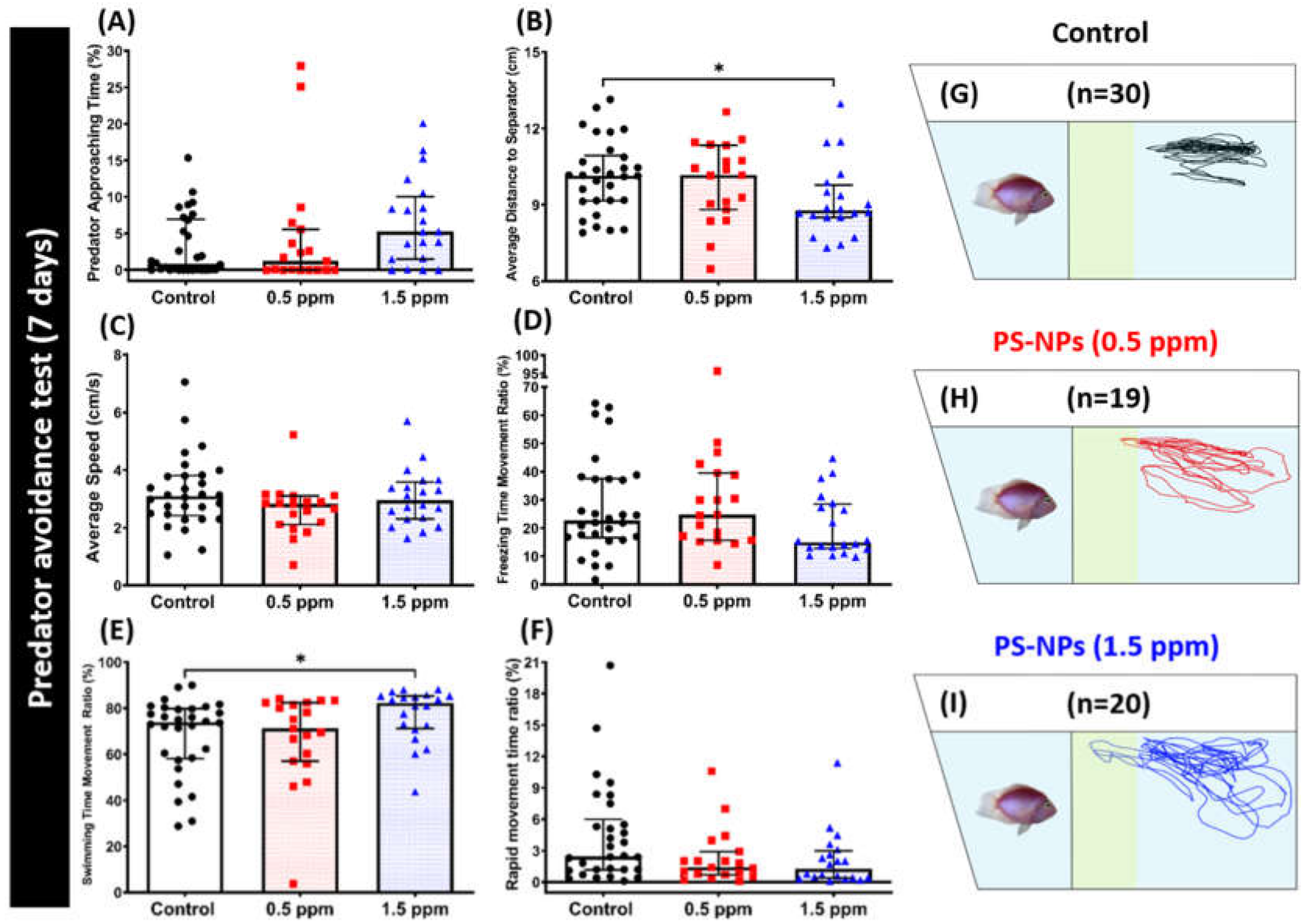
2.3. PS-NPs Exposure Reduced Aggression and Predator Avoidance in Zebrafish
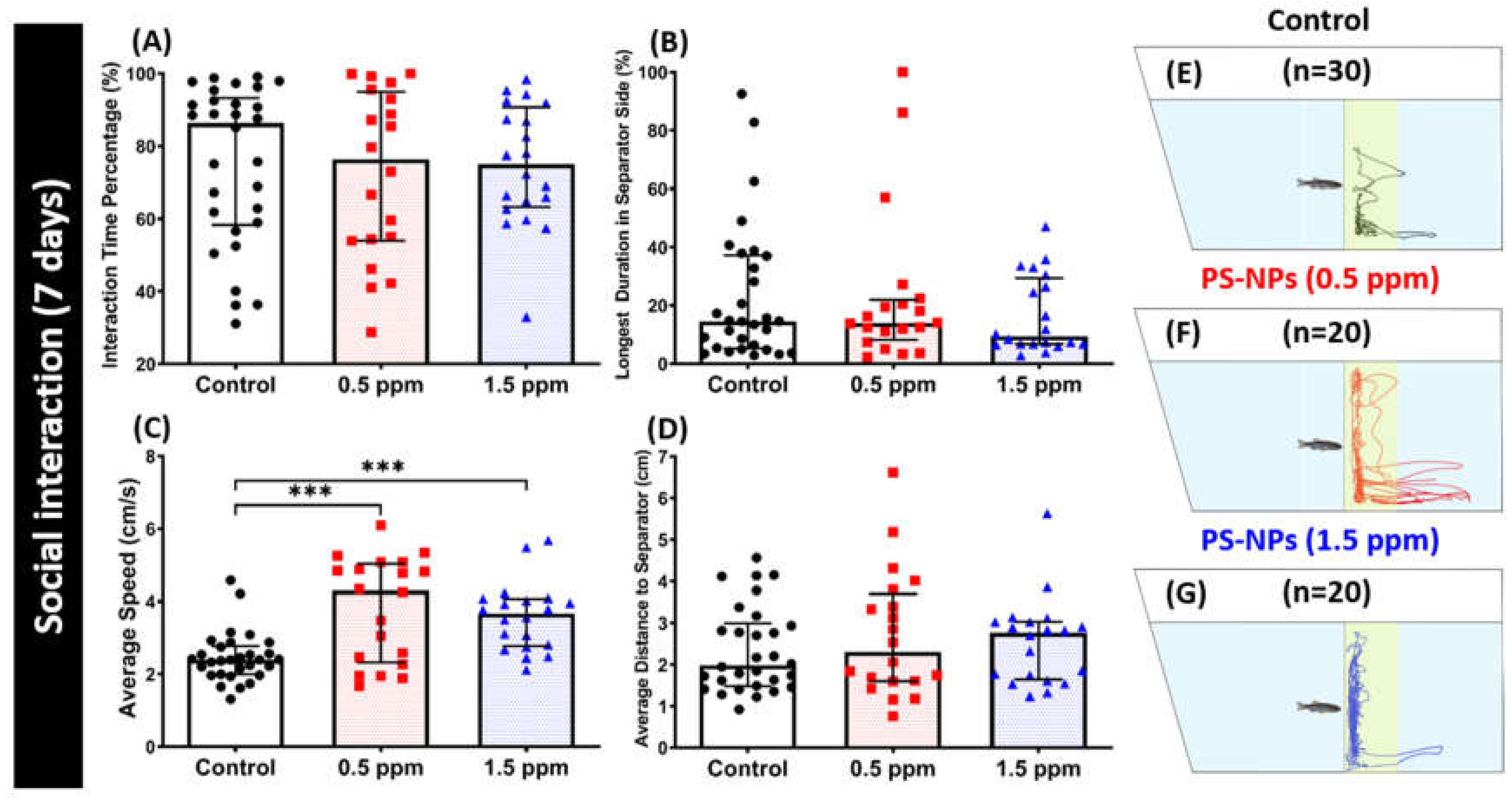
2.4. PS-NPs Exposure Did Not Alter Conspecific Social Behavior in Zebrafish
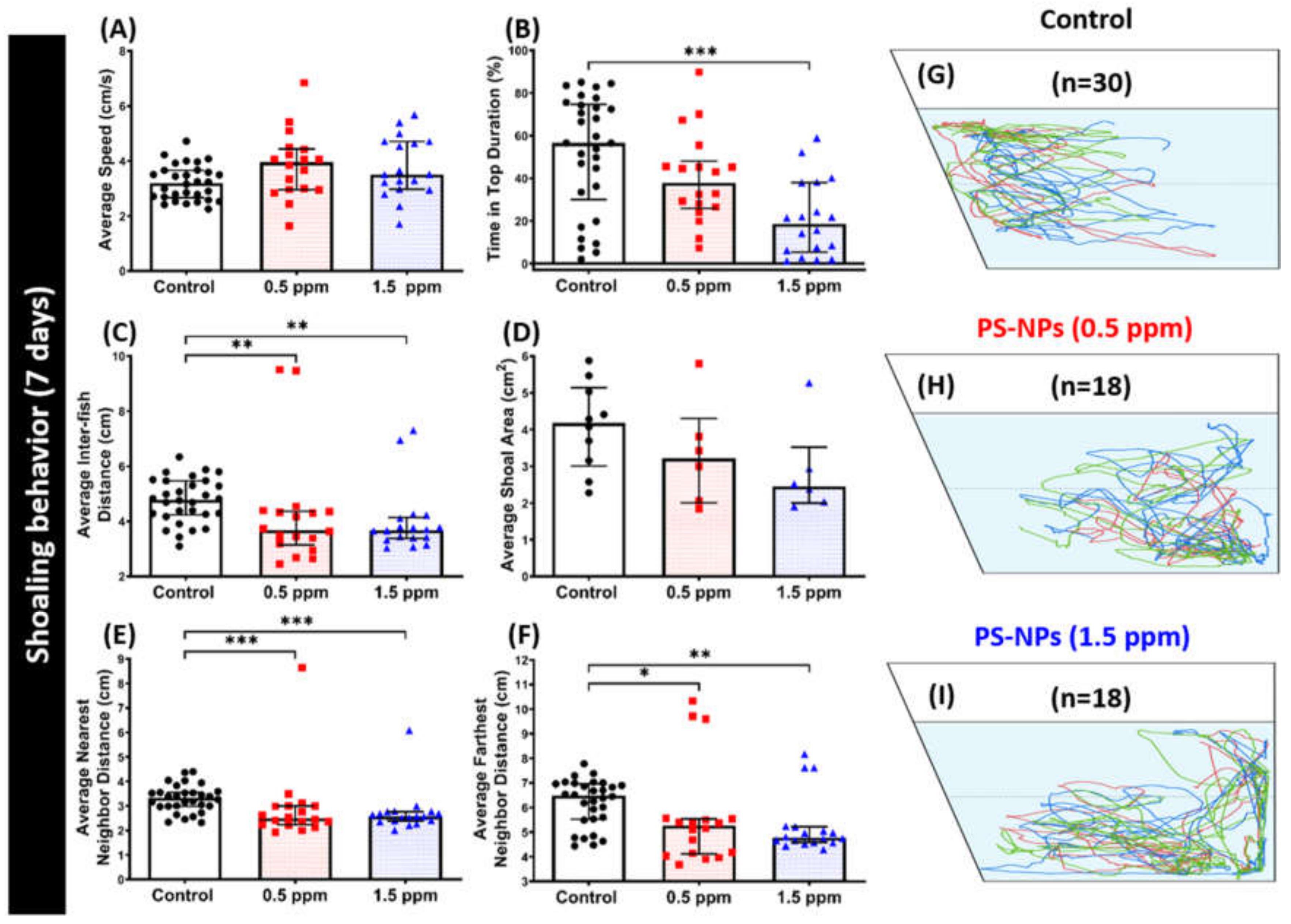
2.5. PS-NPs Exposure Tighten the Shoal in Zebrafish
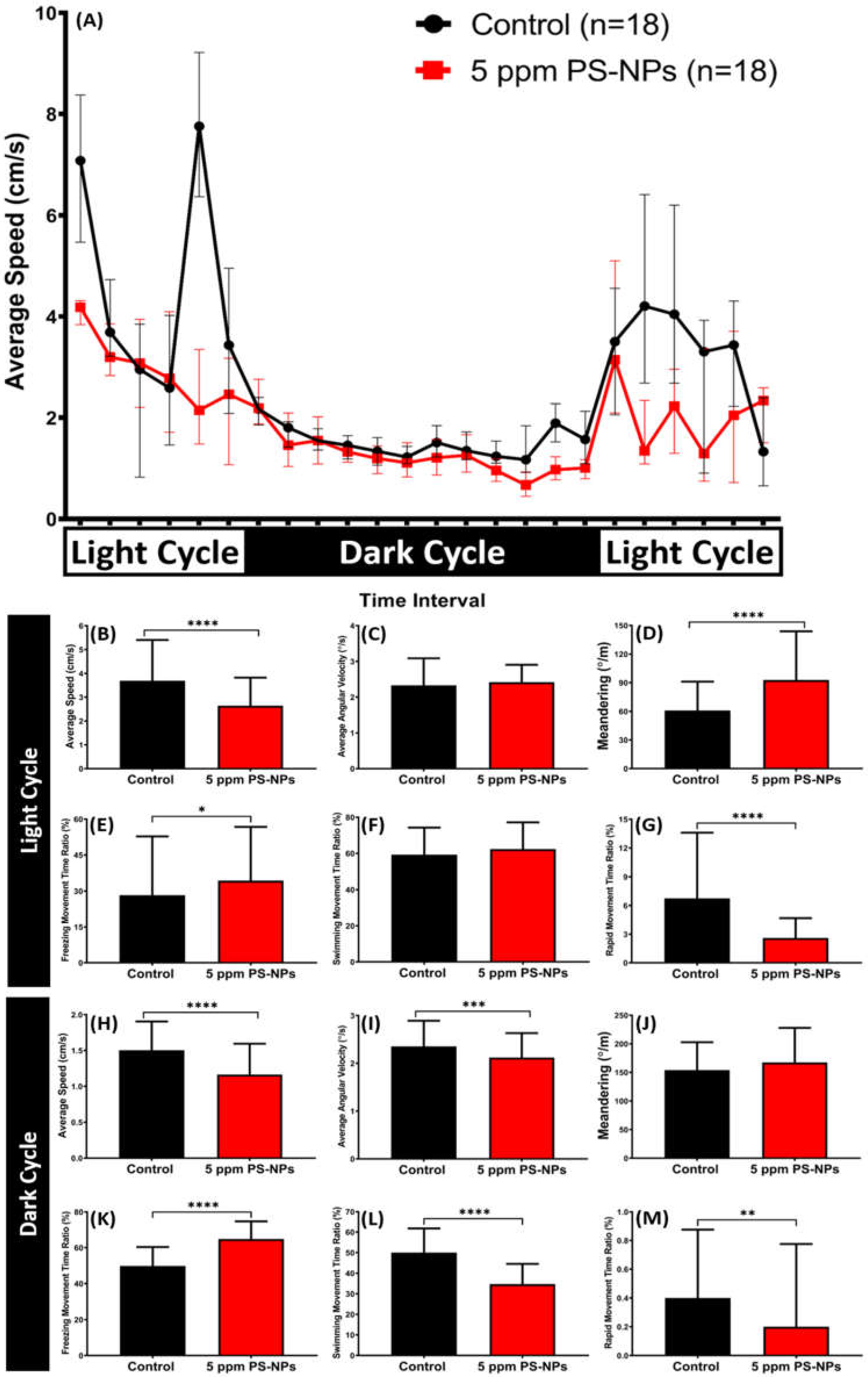
2.6. High Dose of PS-NPs Exposure Dysregulated the Circadian Rhythm
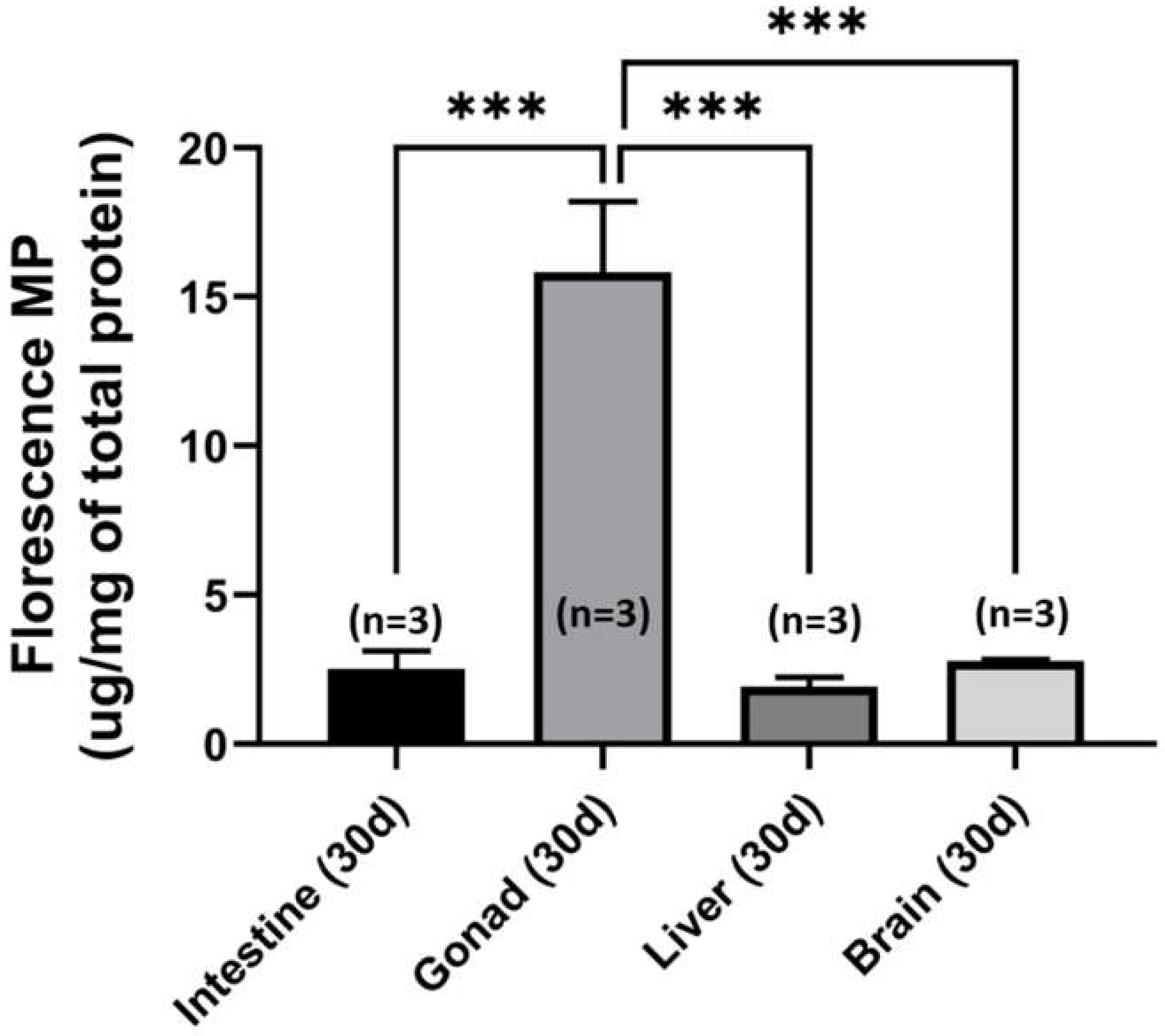
2.7. Tissue Distribution of Fluorescent PS-NPs in Zebrafish
2.8. Measurement of Marker Expression in Muscle and Liver after PS-NPs Exposure
2.9. Measurement of Neurotransmitter Expression in Brain after PS-NPs Exposure
3. Discussion
3.1. Microplastic Pollutions
3.2. Inflammatory Protein Expression and Oxidative Stress
3.3. Energy Metabolism
3.4. Reproductive Toxicity of PS-NPs
3.5. Behavioral and Neurotransmitters Alterations Caused by PS-NPs
4. Conclusions
5. Materials and Methods
5.1. Particle Characterization
5.2. Zebrafish Husbandry
5.3. Exposure of PS-NPs
5.4. Adult Behavior Test Battery
5.5. Circadian Rhythm Locomotion Activity Assay
5.6. Biochemical Analysis of Biomarkers
5.7. Florescence Analysis of Lumisphere Microplastics on Adult Zebrafish
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Gundlach, M.; Yang, S.; Jiang, J.; Velki, M.; Yin, D.; Hollert, H. Quantitative investigation of the mechanisms of microplastics and nanoplastics toward zebrafish larvae locomotor activity. Sci. Total. Environ. 2017, 584, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Lusher, A.; Thompson, R.C.; Morley, A. The deposition and accumulation of microplastics in marine sediments and bottom water from the irish continental shelf. Sci. Rep. 2017, 7, 10772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, A.; Kubowicz, S.; Beegle-Krause, C.; Skancke, J.; Nordam, T.; Landsem, E.; Throne-Holst, M.; Jahren, S. Microplastic in global and norwegian marine environments: Distributions, degradation mechanisms and transport. In Miljødirektoratet M-918; SINTEF: Trondheim, Norway, 2017. [Google Scholar]
- Auta, H.; Emenike, C.; Fauziah, S. Distribution and importance of microplastics in the marine environment: A review of the sources, fate, effects, and potential solutions. Environ. Int. 2017, 102, 165–176. [Google Scholar] [CrossRef]
- Law, K.L.; Thompson, R.C. Microplastics in the seas. Science 2014, 345, 144–145. [Google Scholar] [CrossRef]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 116. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Besseling, E.; Shim, W.J. Nanoplastics in the aquatic environment. Critical review; In Marine Anthropogenic Litter; Springer: Cham, Switzerland, 2015; pp. 325–340. [Google Scholar]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Goodhead, R.; Moger, J.; Galloway, T.S. Microplastic ingestion by zooplankton. Environ. Sci. Technol. 2013, 47, 6646–6655. [Google Scholar] [CrossRef]
- Devriese, L.I.; van der Meulen, M.D.; Maes, T.; Bekaert, K.; Paul-Pont, I.; Frère, L.; Robbens, J.; Vethaak, A.D. Microplastic contamination in brown shrimp (crangon crangon, linnaeus 1758) from coastal waters of the southern north sea and channel area. Mar. Pollut. Bull. 2015, 98, 179–187. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P.A.; Leusch, F.D. Impact of microplastic beads and fibers on waterflea (ceriodaphnia dubia) survival, growth, and reproduction: Implications of single and mixture exposures. Environ. Sci. Technol. 2017, 51, 13397–13406. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, P.; Fonte, E.; Soares, M.E.; Carvalho, F.; Guilhermino, L. Effects of multi-stressors on juveniles of the marine fish pomatoschistus microps: Gold nanoparticles, microplastics and temperature. Aquat. Toxicol. 2016, 170, 89–103. [Google Scholar] [CrossRef]
- Pedà, C.; Caccamo, L.; Fossi, M.C.; Gai, F.; Andaloro, F.; Genovese, L.; Perdichizzi, A.; Romeo, T.; Maricchiolo, G. Intestinal alterations in european sea bass dicentrarchus labrax (linnaeus, 1758) exposed to microplastics: Preliminary results. Environ. Pollut. 2016, 212, 251–256. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hoh, E.; Kurobe, T.; Teh, S.J. Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci. Rep. 2013, 3, 3263. [Google Scholar] [CrossRef]
- Karami, A.; Golieskardi, A.; Ho, Y.B.; Larat, V.; Salamatinia, B. Microplastics in eviscerated flesh and excised organs of dried fish. Sci. Rep. 2017, 7, 5473. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Hong, S.H.; Jang, M.; Han, G.M.; Shim, W.J. Occurrence and distribution of microplastics in the sea surface microlayer in jinhae bay, south korea. Arch. Environ. Contam. Toxicol. 2015, 69, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Hong, S.H.; Jang, M.; Kang, J.-H.; Kwon, O.Y.; Han, G.M.; Shim, W.J. Large accumulation of micro-sized synthetic polymer particles in the sea surface microlayer. Environ. Sci. Technol. 2014, 48, 9014–9021. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.A.; Niven, S.J.; Galloway, T.S.; Rowland, S.J.; Thompson, R.C. Microplastic moves pollutants and additives to worms, reducing functions linked to health and biodiversity. Curr. Biol. 2013, 23, 2388–2392. [Google Scholar] [CrossRef] [Green Version]
- Von Moos, N.; Burkhardt-Holm, P.; Köhler, A. Uptake and effects of microplastics on cells and tissue of the blue mussel mytilus edulis l. After an experimental exposure. Environ. Sci. Technol. 2012, 46, 11327–11335. [Google Scholar] [CrossRef]
- Watts, A.J.; Lewis, C.; Goodhead, R.M.; Beckett, S.J.; Moger, J.; Tyler, C.R.; Galloway, T.S. Uptake and retention of microplastics by the shore crab carcinus maenas. Environ. Sci. Technol. 2014, 48, 8823–8830. [Google Scholar] [CrossRef]
- E Silva, P.P.G.; Nobre, C.R.; Resaffe, P.; Pereira, C.D.S.; Gusmão, F. Leachate from microplastics impairs larval development in brown mussels. Water Res. 2016, 106, 364–370. [Google Scholar] [CrossRef]
- Kaposi, K.L.; Mos, B.; Kelaher, B.P.; Dworjanyn, S.A. Ingestion of microplastic has limited impact on a marine larva. Environ. Sci. Technol. 2014, 48, 1638–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadri, S.S.; Thompson, R.C. On the quantity and composition of floating plastic debris entering and leaving the tamar estuary, southwest england. Mar. Pollut. Bull. 2014, 81, 55–60. [Google Scholar] [CrossRef]
- Blaser, R.E.; Rosemberg, D.B. Measures of anxiety in zebrafish (danio rerio): Dissociation of black/white preference and novel tank test. PLoS ONE 2012, 7, e36931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, M.; Raymond, J.; Hester, J.; Kyzar, E.; Gaikwad, S.; Bruce, I.; Fryar, C.; Chanin, S.; Enriquez, J.; Bagawandoss, S. Assessing social behavior phenotypes in adult zebrafish: Shoaling, social preference, and mirror biting tests. In Zebrafish Protocols for Neurobehavioral Research; Springer: Berlin, Germany, 2012; pp. 231–246. [Google Scholar]
- Moretz, J.A.; Martins, E.P.; Robison, B.D. Behavioral syndromes and the evolution of correlated behavior in zebrafish. Behav. Ecol. 2007, 18, 556–562. [Google Scholar] [CrossRef] [Green Version]
- Speedie, N.; Gerlai, R. Alarm substance induced behavioral responses in zebrafish (danio rerio). Behav. Brain Res. 2008, 188, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledesma, J.M.; McRobert, S.P. Innate and learned shoaling preferences based on body coloration in juvenile mollies, poecilia latipinna. Ethology 2008, 114, 1044–1048. [Google Scholar] [CrossRef]
- Morrell, L.J.; James, R. Mechanisms for aggregation in animals: Rule success depends on ecological variables. Behav. Ecol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Bégay, V.R.; Falcón, J.; Cahill, G.M.; Klein, D.C.; Coon, S.L. Transcripts encoding two melatonin synthesis enzymes in the teleost pineal organ: Circadian regulation in pike and zebrafish, but not in trout. Endocrinology 1998, 139, 905–912. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R. Molecular analysis of mammalian circadian rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Desforges, J.-P.W.; Galbraith, M.; Dangerfield, N.; Ross, P.S. Widespread distribution of microplastics in subsurface seawater in the ne pacific ocean. Mar. Pollut. Bull. 2014, 79, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Nor, N.H.M.; Hermsen, E.; Kooi, M.; Mintenig, S.M.; De France, J. Microplastics in freshwaters and drinking water: Critical review and assessment of data quality. Water Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qu, X.; Su, L.; Zhang, W.; Yang, D.; Kolandhasamy, P.; Li, D.; Shi, H. Microplastics in mussels along the coastal waters of china. Environ. Pollut. 2016, 214, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, X.; Li, Y.; Wang, Y.; Yang, L. A large-scale association study for nanoparticle c60 uncovers mechanisms of nanotoxicity disrupting the native conformations of DNA/rna. Nucleic Acids Res. 2012, 40, 7622–7632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.-M.; Jeong, C.-B.; Lee, Y.H.; Cui, Y.-H.; Kim, D.-H.; Lee, M.-C.; Kim, H.-S.; Han, J.; Hwang, D.-S.; Lee, S.-J. Cross-reactivities of mammalian mapks antibodies in rotifer and copepod: Application in mechanistic studies in aquatic ecotoxicology. Mar. Pollut. Bull. 2017, 124, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Honma, R.; Sumita, M.; Hanawa, T. Cytotoxicity evaluation of ceramic particles of different sizes and shapes. J. Biomed. Mater. Res. Part A 2004, 68, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Watts, D.J. Particle surface characteristics may play an important role in phytotoxicity of alumina nanoparticles. Toxicol. Lett. 2005, 158, 122–132. [Google Scholar] [CrossRef]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Galloway, T.S. The impact of polystyrene microplastics on feeding, function and fecundity in the marine copepod calanus helgolandicus. Environ. Sci. Technol. 2015, 49, 1130–1137. [Google Scholar] [CrossRef]
- Cedervall, T.; Hansson, L.-A.; Lard, M.; Frohm, B.; Linse, S. Food chain transport of nanoparticles affects behaviour and fat metabolism in fish. PLoS ONE 2012, 7, e32254. [Google Scholar] [CrossRef] [Green Version]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Le Goïc, N.; Quillien, V.; Mingant, C.; Epelboin, Y. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, P.R.R.; Reis-Henriques, M.A.; Coimbra, J. Polycyclic aromatic hydrocarbons inhibit in vitro ovarian steroidogenesis in the flounder (platichthys flesus l.). Aquat. Toxicol. 2000, 48, 549–559. [Google Scholar] [CrossRef]
- Britt, K.L.; Saunders, P.K.; McPherson, S.J.; Misso, M.L.; Simpson, E.R.; Findlay, J.K. Estrogen actions on follicle formation and early follicle development. Biol. Reprod. 2004, 71, 1712–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.K.; Singh, J.M.; Leslie, T.C.; Meachum, S.; Flaws, J.A.; Yao, H.H. Di-(2-ethylhexyl) phthalate and mono-(2-ethylhexyl) phthalate inhibit growth and reduce estradiol levels of antral follicles in vitro. Toxicol. Appl. Pharmacol. 2010, 242, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, M.D. Composition, accumulation and utilization of yolk lipids in teleost fish. Rev. Fish Biol. Fish. 1996, 6, 259–286. [Google Scholar] [CrossRef]
- Lazier, C. Vitellogenin gene expression in teleost fish. Biochem. Mol. Biol. Fishes 1993, 391–405. [Google Scholar]
- Arukwe, A.; Goksøyr, A. Eggshell and egg yolk proteins in fish: Hepatic proteins for the next generation: Oogenetic, population, and evolutionary implications of endocrine disruption. Comp. Hepatol. 2003, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Denslow, N.D.; Chow, M.C.; Kroll, K.J.; Green, L. Vitellogenin as a biomarker of exposure for estrogen or estrogen mimics. Ecotoxicology 1999, 8, 385–398. [Google Scholar] [CrossRef]
- Zhong, L.; Yuan, L.; Rao, Y.; Li, Z.; Zhang, X.; Liao, T.; Xu, Y.; Dai, H. Distribution of vitellogenin in zebrafish (danio rerio) tissues for biomarker analysis. Aquat. Toxicol. 2014, 149, 1–7. [Google Scholar] [CrossRef]
- Ota, M.; Saito, T.; Yoshizaki, G.; Otsuki, A. Vitellogenin-like gene expression in liver of male zebrafish (danio rerio) by 1 μm 3-methylcholanthrene treatment: The possibility of a rapid and sensitive in vivo bioassay. Water Res. 2000, 34, 2400–2403. [Google Scholar] [CrossRef]
- Marin, M.G.; Matozzo, V. Vitellogenin induction as a biomarker of exposure to estrogenic compounds in aquatic environments. Mar. Pollut. Bull. 2004, 48, 835–839. [Google Scholar] [CrossRef]
- Sumpter, J.P.; Jobling, S. Vitellogenesis as a biomarker for estrogenic contamination of the aquatic environment. Environ. Health Perspect. 1995, 103, 173–178. [Google Scholar] [PubMed]
- Peéry, A.R.; Devillers, J.; Brochot, C.; Mombelli, E.; Palluel, O.; Piccini, B.; Brion, F.; Beaudouin, R.m. A physiologically based toxicokinetic model for the zebrafish danio rerio. Environ. Sci. Technol. 2013, 48, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ying, G.-G.; Jones, K.C.; Martin, F.L. Real-world carbon nanoparticle exposures induce brain and gonadal alterations in zebrafish (danio rerio) as determined by biospectroscopy techniques. Analyst 2015, 140, 2687–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.-B.; Lu, C.-J.; Junaid, M.; Jia, P.-P.; Yang, L.; Zhang, J.-H.; Pei, D.-S. Potential adverse outcome pathway (aop) of silver nanoparticles mediated reproductive toxicity in zebrafish. Chemosphere 2018, 207, 320–328. [Google Scholar] [CrossRef]
- Orbea, A.; González-Soto, N.; Lacave, J.M.; Barrio, I.; Cajaraville, M.P. Developmental and reproductive toxicity of pvp/pei-coated silver nanoparticles to zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 199, 59–68. [Google Scholar] [CrossRef]
- Oberdörster, G.; Sharp, Z.; Atudorei, V.; Elder, A.; Gelein, R.; Kreyling, W.; Cox, C. Translocation of inhaled ultrafine particles to the brain. Inhal. Toxicol. 2004, 16, 437–445. [Google Scholar] [CrossRef]
- Hou, C.-C.; Zhu, J.-Q. Nanoparticles and female reproductive system: How do nanoparticles affect oogenesis and embryonic development. Oncotarget 2017, 8, 109799. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, A.J.; Billig, H.; Tsafriri, A. Ovarian follicle atresia: A hormonally controlled apoptotic process. Endocr. Rev. 1994, 15, 707–724. [Google Scholar]
- Rollerova, E.; Jurcovicova, J.; Mlynarcikova, A.; Sadlonova, I.; Bilanicova, D.; Wsolova, L.; Kiss, A.; Kovriznych, J.; Kronek, J.; Ciampor, F. Delayed adverse effects of neonatal exposure to polymeric nanoparticle poly (ethylene glycol)-block-polylactide methyl ether on hypothalamic–pituitary–ovarian axis development and function in wistar rats. Reprod. Toxicol. 2015, 57, 165–175. [Google Scholar] [CrossRef]
- Dayal, N.; Thakur, M.; Patil, P.; Singh, D.; Vanage, G.; Joshi, D. Histological and genotoxic evaluation of gold nanoparticles in ovarian cells of zebrafish (danio rerio). J. Nanoparticle Res. 2016, 18, 291. [Google Scholar] [CrossRef]
- Yan, S.-Q.; Xing, R.; Zhou, Y.-F.; Li, K.-L.; Su, Y.-Y.; Qiu, J.-F.; Zhang, Y.-H.; Zhang, K.-Q.; He, Y.; Lu, X.-P. Reproductive toxicity and gender differences induced by cadmium telluride quantum dots in an invertebrate model organism. Sci. Rep. 2016, 6, 34182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Song, B.; Wu, J.; Zhang, Y.; Chen, A.; Shao, L. Potential adverse effects of nanoparticles on the reproductive system. Int. J. Nanomed. 2018, 13, 8487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.L.; Brown, R.M. Selective investment theory: Recasting the functional significance of close relationships. Psychol. Inq. 2006, 17, 1–29. [Google Scholar] [CrossRef]
- Neumann, I.D. Brain oxytocin: A key regulator of emotional and social behaviours in both females and males. J. Neuroendocrinol. 2008, 20, 858–865. [Google Scholar] [CrossRef] [PubMed]
- De Dreu, C.K.; Shalvi, S.; Greer, L.L.; Van Kleef, G.A.; Handgraaf, M.J. Oxytocin motivates non-cooperation in intergroup conflict to protect vulnerable in-group members. PLoS ONE 2012, 7, e46751. [Google Scholar] [CrossRef] [Green Version]
- Parhar, I.S.; Ogawa, S.; Ubuka, T. Reproductive neuroendocrine pathways of social behavior. Front. Endocrinol. 2016, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Mineur, Y.S.; Obayemi, A.; Wigestrand, M.B.; Fote, G.M.; Calarco, C.A.; Li, A.M.; Picciotto, M.R. Cholinergic signaling in the hippocampus regulates social stress resilience and anxiety-and depression-like behavior. Proc. Natl. Acad. Sci. USA 2013, 110, 3573–3578. [Google Scholar] [CrossRef] [Green Version]
- Chacón, M.A.; Reyes, A.E.; Inestrosa, N.C. Acetylcholinesterase induces neuronal cell loss, astrocyte hypertrophy and behavioral deficits in mammalian hippocampus. J. Neurochem. 2003, 87, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Ribeiro, A.; Hylland, K.; Guilhermino, L. Single and combined effects of microplastics and pyrene on juveniles (0+ group) of the common goby pomatoschistus microps (teleostei, gobiidae). Ecol. Indic. 2013, 34, 641–647. [Google Scholar] [CrossRef]
- Ribeiro, F.; Garcia, A.R.; Pereira, B.P.; Fonseca, M.; Mestre, N.C.; Fonseca, T.G.; Ilharco, L.M.; Bebianno, M.J. Microplastics effects in scrobicularia plana. Mar. Pollut. Bull. 2017, 122, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Wei, R.; Niu, R.; Wang, C.; Wang, J. Pubertal exposure to bisphenol a increases anxiety-like behavior and decreases acetylcholinesterase activity of hippocampus in adult male mice. Food Chem. Toxicol. 2013, 60, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.; Santana, M.; Do Nascimento, J.; Picanço-Diniz, D.; Maués, L.; Santos, S.; Ferreira, V.; Alfonso, M.; Duran, R.; Faro, L. Behavioral and biochemical effects of neonicotinoid thiamethoxam on the cholinergic system in rats. Ecotoxicol. Environ. Saf. 2010, 73, 101–107. [Google Scholar] [CrossRef] [PubMed]
- DuRant, S.E.; Hopkins, W.A.; Talent, L.G. Impaired terrestrial and arboreal locomotor performance in the western fence lizard (sceloporus occidentalis) after exposure to an ache-inhibiting pesticide. Environ. Pollut. 2007, 149, 18–24. [Google Scholar] [CrossRef]
- Manzanares, P.A.R.; Isoardi, N.A.; Carrer, H.F.; Molina, V.A. Previous stress facilitates fear memory, attenuates gabaergic inhibition, and increases synaptic plasticity in the rat basolateral amygdala. J. Neurosci. 2005, 25, 8725–8734. [Google Scholar] [CrossRef]
- Lira, A.; Zhou, M.; Castanon, N.; Ansorge, M.S.; Gordon, J.A.; Francis, J.H.; Bradley-Moore, M.; Lira, J.; Underwood, M.D.; Arango, V. Altered depression-related behaviors and functional changes in the dorsal raphe nucleus of serotonin transporter-deficient mice. Biol. Psychiatry 2003, 54, 960–971. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, H.J.; Kim, J.G.; Ryu, V.; Kim, B.-T.; Kang, D.-W.; Jahng, J.W. Depressive behaviors and decreased expression of serotonin reuptake transporter in rats that experienced neonatal maternal separation. Neurosci. Res. 2007, 58, 32–39. [Google Scholar] [CrossRef]
- Wysocki, L.E.; Dittami, J.P.; Ladich, F. Ship noise and cortisol secretion in european freshwater fishes. Biol. Conserv. 2006, 128, 501–508. [Google Scholar] [CrossRef]
- Øverli, Ø.; Kotzian, S.; Winberg, S. Effects of cortisol on aggression and locomotor activity in rainbow trout. Horm. Behav. 2002, 42, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Ariyomo, T.O.; Carter, M.; Watt, P.J. Heritability of boldness and aggressiveness in the zebrafish. Behav. Genet. 2013, 43, 161–167. [Google Scholar] [CrossRef]
- Giammanco, M.; Tabacchi, G.; Giammanco, S.; Di Majo, D.; La Guardia, M. Testosterone and aggressiveness. Med. Sci. Monit. 2005, 11, RA136–RA145. [Google Scholar] [PubMed]
- Winslow, J.T.; Insel, T.R. Social status in pairs of male squirrel monkeys determines the behavioral response to central oxytocin administration. J. Neurosci. 1991, 11, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.G.; Hofmann, H.A. Neuropeptide regulation of social behavior in a monogamous cichlid fish. Physiol. Behav. 2011, 102, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Coverdill, A.; McCarthy, M.; Bridges, R.; Nephew, B. Effects of chronic central arginine vasopressin (avp) on maternal behavior in chronically stressed rat dams. Brain Sci. 2012, 2, 589–604. [Google Scholar] [CrossRef] [Green Version]
- Ogrizek, M.; Grgurevič, N.; Snoj, T.; Majdič, G. Injections to pregnant mice produce prenatal stress that affects aggressive behavior in their adult male offspring. Horm. Behav. 2018, 106, 35–43. [Google Scholar] [CrossRef]
- Kanda, S.; Akazome, Y.; Mitani, Y.; Okubo, K.; Oka, Y. Neuroanatomical evidence that kisspeptin directly regulates isotocin and vasotocin neurons. PLoS ONE 2013, 8, e62776. [Google Scholar] [CrossRef] [Green Version]
- Campbell, P.; Ophir, A.G.; Phelps, S.M. Central vasopressin and oxytocin receptor distributions in two species of singing mice. J. Comp. Neurol. 2009, 516, 321–333. [Google Scholar] [CrossRef]
- Miczek, K.A.; Fish, E.W.; Joseph, F.; de Almeida, R.M. Social and neural determinants of aggressive behavior: Pharmacotherapeutic targets at serotonin, dopamine and γ-aminobutyric acid systems. Psychopharmacology 2002, 163, 434–458. [Google Scholar] [CrossRef]
- De Almeida, R.M.; Ferrari, P.F.; Parmigiani, S.; Miczek, K.A. Escalated aggressive behavior: Dopamine, serotonin and gaba. Eur. J. Pharmacol. 2005, 526, 51–64. [Google Scholar] [CrossRef]
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.; Seguin, D.; Gerlai, R. An automated predator avoidance task in zebrafish. Behav. Brain Res. 2011, 216, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audira, G.; Sampurna, B.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A versatile setup for measuring multiple behavior endpoints in zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, S.; Nathan, F.M.; Parhar, I.S. Habenular kisspeptin modulates fear in the zebrafish. Proc. Natl. Acad. Sci. USA 2014, 111, 3841–3846. [Google Scholar] [CrossRef] [Green Version]
- Brambilla, P.; Perez, J.; Barale, F.; Schettini, G.; Soares, J. Gabaergic dysfunction in mood disorders. Mol. Psychiatry 2003, 8, 721. [Google Scholar] [CrossRef] [Green Version]
- Tsubokawa, T.; Saito, K.; Kawano, H.; Kawamura, K.; Shinozuka, K.; Watanabe, S. Pharmacological effects on mirror approaching behavior and neurochemical aspects of the telencephalon in the fish, medaka (oryzias latipes). Soc. Neurosci. 2009, 4, 276–286. [Google Scholar] [CrossRef]
- Reynolds, S.M.; Berridge, K.C. Fear and feeding in the nucleus accumbens shell: Rostrocaudal segregation of gaba-elicited defensive behavior versus eating behavior. J. Neurosci. 2001, 21, 3261–3270. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.; Laland, K. Social learning and life skills training for hatchery reared fish. J. Fish Biol. 2001, 59, 471–493. [Google Scholar] [CrossRef]
- Green, J.; Collins, C.; Kyzar, E.J.; Pham, M.; Roth, A.; Gaikwad, S.; Cachat, J.; Stewart, A.M.; Landsman, S.; Grieco, F.; et al. Automated high-throughput neurophenotyping of zebrafish social behavior. J. Neurosci. Methods 2012, 210, 266–271. [Google Scholar] [CrossRef]
- Young, L.J.; Flanagan-Cato, L.M. Editorial comment: Oxytocin, vasopressin and social behavior. Horm. Behav. 2012, 61, 227. [Google Scholar] [CrossRef] [Green Version]
- Winslow, J.T.; Insel, T.R. The social deficits of the oxytocin knockout mouse. Neuropeptides 2002, 36, 221–229. [Google Scholar] [CrossRef]
- Shams, S.; Amlani, S.; Buske, C.; Chatterjee, D.; Gerlai, R. Developmental social isolation affects adult behavior, social interaction, and dopamine metabolite levels in zebrafish. Dev. Psychobiol. 2018, 60, 43–56. [Google Scholar] [CrossRef]
- Saif, M.; Chatterjee, D.; Buske, C.; Gerlai, R. Sight of conspecific images induces changes in neurochemistry in zebrafish. Behav. Brain Res. 2013, 243, 294–299. [Google Scholar] [CrossRef]
- Scerbina, T.; Chatterjee, D.; Gerlai, R. Dopamine receptor antagonism disrupts social preference in zebrafish: A strain comparison study. Amino Acids 2012, 43, 2059–2072. [Google Scholar] [CrossRef]
- Nishimura, Y.; Yoshida, M.; Watanabe, S. The effect on other individual presentations of the goldfish by fg7142 injection. Nihon shinkei seishin yakurigaku zasshi. Jpn. J. Psychopharmacol. 2002, 22, 55–59. [Google Scholar]
- Lahiri, K.; Vallone, D.; Gondi, S.B.; Santoriello, C.; Dickmeis, T.; Foulkes, N.S. Temperature regulates transcription in the zebrafish circadian clock. PLoS Biol. 2005, 3, e351. [Google Scholar] [CrossRef] [Green Version]
- Auld, F.; Maschauer, E.L.; Morrison, I.; Skene, D.J.; Riha, R.L. Evidence for the efficacy of melatonin in the treatment of primary adult sleep disorders. Sleep Med. Rev. 2017, 34, 10–22. [Google Scholar] [CrossRef]
- Zhdanova, I.V.; Wang, S.Y.; Leclair, O.U.; Danilova, N.P. Melatonin promotes sleep-like state in zebrafish. Brain Res. 2001, 903, 263–268. [Google Scholar] [CrossRef]
- Altun, A.; Ugur-Altun, B. Melatonin: Therapeutic and clinical utilization. Int. J. Clin. Pract. 2007, 61, 835–845. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Li, T.; Li, X.; Feng, D.; Kuang, X.; Xu, J.; Zhao, X.; Sun, M.; Chen, D. Sio 2 nanoparticles cause depression and anxiety-like behavior in adult zebrafish. RSC Adv. 2017, 7, 2953–2963. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-D.; Burton, K.J.; Zhang, C.; Hu, S.-B.; Zhou, Q.-Y. Vasopressin receptor v1a regulates circadian rhythms of locomotor activity and expression of clock-controlled genes in the suprachiasmatic nuclei. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R824–R830. [Google Scholar] [CrossRef] [Green Version]
- Wisor, J.; Wurts, S.; Hall, F.; Lesch, K.; Murphy, D.; Uhl, G.; Edgar, D. Altered rapid eye movement sleep timing in serotonin transporter knockout mice. Neuroreport 2003, 14, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Escudero, A.; Vicente-Page, J.; Hinz, R.C.; Arganda, S.; De Polavieja, G.G. Idtracker: Tracking individuals in a group by automatic identification of unmarked animals. Nat. Methods 2014, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A simple setup to perform 3d locomotion tracking in zebrafish by using a single camera. Inventions 2018, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Han, L.; Hsiao, C.-D. Establishing simple image-based methods and a cost-effective instrument for toxicity assessment on circadian rhythm dysregulation in fish. Biol. Open 2019, 8, bio041871. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarker | WT | PS-NPs (0.5 ppm) | PS-NPs (1.5 ppm) | Unit | Significance | ANOVA F Value | p Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Muscle | |||||||||
| ROS | 4.53 ± 0.3 | 5.27 ± 0.9 | NS | 7.31 ± 0.32 | * | U/ug of total protein | YES | F (2, 6) =5.862 | p = 0.0388 |
| ATP | 296.4 ± 5.80 | 290.3 ± 4.36 | NS | 242.4 ± 3.78 | *** | ng/ug of total protein | YES | F (2,6) = 39.27 | p = 0.0004 |
| Creatine kinase | 6.70 ± 0.11 | 7.50 ± 0.78 | NS | 6.88 ± 0.39 | NS | pg/ug of total protein | NO | F (2,6) = 0.468 | p = 0.646 |
| Hif-1α | 18.77 ± 0.55 | 21.07 ± 1.6 | NS | 22.26 ± 0.30 | NS | pg/ug of total protein | NO | F (2,6) = 2.974 | p = 0.126 |
| Liver | |||||||||
| ssDNA | 2.63 ± 0.29 | 2.28 ± 0.61 | NS | 5.13 ± 0.58 | * | U/ug of total protein | YES | F (2,6) = 9.0 | p = 0.01 |
| TNF-α | 21.71 ± 1.67 | 17.59 ± 3.37 | NS | 45.26 ± 8.20 | * | pg/ug of total protein | YES | F (2,6) = 8.20 | p = 0.01 |
| MDA | 0.32 ± 0.08 | 0.20 ± 0.06 | NS | 0.27 ± 0.06 | NS | pg/ug of total protein | NO | F (2,6) = 0.84 | p = 0.47 |
| Cortisol | 64.76 ± 5.50 | 57.35 ± 5.50 | NS | 88.5 ± 6.93 | * | pg/ug of total protein | YES | F (2,6) = 9.60 | p = 0.01 |
| EROD | 362.5 ± 45.89 | 315.2 ± 29.00 | NS | 319.7 ± 8.40 | NS | U/ug of total protein | NO | F (2,6) = 0.67 | p = 0.5 |
| VTG | 32.14 ± 2.5 | 54.19 ± 3.8 | ** | 68.34 ± 2.2 | *** | ng/ug of total protein | YES | F (2,6) = 37.82 | p = 0.0004 |
| CYP1A1 | 2.4 ± 0.62 | 1.01 ± 0.24 | NS | 0.77 ± 0.03 | * | ng/ug of total protein | YES | F (2,6) = 5.42 | p = 0.045 |
| CYP11A1 | 0.30 ± 0.03 | 0.27 ± 0.02 | NS | 0.50 ± 0.05 | * | ng/ug of total protein | YES | F (2,6) = 9.64 | p = 0.013 |
| CYP19A1 | 0.42 ± 0.07 | 0.28 ± 0.03 | NS | 0.77 ± 0.01 | ** | ng/ug of total protein | YES | F (2,6) = 25.38 | p = 0.001 |
| Brain | |||||||||
| AChE | 19.48 ± 0.90 | 16.73 ± 0.73 | * | 13.5 ± 0.02 | ** | U/ug of total protein | YES | F (2,6) = 20.86 | p = 0.002 |
| Acetylcholine | 32.24 ± 1.80 | 30.02 ± 1.90 | NS | 29.54 ± 1.20 | NS | U/ug of total protein | NO | F (2,6) = 14.16 | p = 0.005 |
| Dopamine | 66.80 ± 2.80 | 60.57 ± 2.25 | NS | 50.42 ± 2.80 | ** | pg/ug of total protein | YES | F (2,6) = 9.10 | p = 0.01 |
| Melatonin | 7.91 ± 0.12 | 6.64 ± 0.06 | ** | 5.64 ± 0.23 | *** | pg/ug of total protein | YES | F (2,6) = 52.11 | p = 0.0002 |
| GABA | 0.26 ± 0.01 | 0.23 ± 0.00 | NS | 0.19 ± 0.00 | ** | U/ug of total protein | YES | F (2,6) = 19.48 | p = 0.002 |
| 5-HT | 0.85 ± 0.05 | 0.72 ± 0.01 | NS | 0.58 ± 0.03 | ** | ng/ug of total protein | YES | F (2,6) = 14.16 | p = 0.005 |
| Vassopressin | 4.18 ± 0.23 | 3.28 ± 0.21 | * | 2.47 ± 0.13 | ** | ng/ug of total protein | YES | F (2,6) = 18.58 | p = 0.002 |
| Kisspeptin | 11.72 ± 0.91 | 9.53 ± 0.18 | NS | 7.49 ± 0.31 | ** | ng/ug of total protein | YES | F (2,6) = 13.91 | p = 0.005 |
| PRL | 60.45 ± 6.9 | 39.46 ± 6.9 | NS | 49.01 ± 1.9 | NS | ng/ug of total protein | NO | F (2,6) = 3.26 | p = 0.10 |
| Oxytocin | 24.86 ± 1.7 | 19.85 ± 0.17 | NS | 15.9 ± 1.43 | ** | pg/ug of total protein | YES | F (2,6) = 12.04 | p = 0.007 |
| Vasotocin | 381 ± 15.29 | 364 ± 13.37 | NS | 419 ± 30.58 | NS | pg/ug of total protein | NO | F (2,6) = 1.78 | p = 0.24 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarasamma, S.; Audira, G.; Siregar, P.; Malhotra, N.; Lai, Y.-H.; Liang, S.-T.; Chen, J.-R.; Chen, K.H.-C.; Hsiao, C.-D. Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. Int. J. Mol. Sci. 2020, 21, 1410. https://doi.org/10.3390/ijms21041410
Sarasamma S, Audira G, Siregar P, Malhotra N, Lai Y-H, Liang S-T, Chen J-R, Chen KH-C, Hsiao C-D. Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. International Journal of Molecular Sciences. 2020; 21(4):1410. https://doi.org/10.3390/ijms21041410
Chicago/Turabian StyleSarasamma, Sreeja, Gilbert Audira, Petrus Siregar, Nemi Malhotra, Yu-Heng Lai, Sung-Tzu Liang, Jung-Ren Chen, Kelvin H.-C. Chen, and Chung-Der Hsiao. 2020. "Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure" International Journal of Molecular Sciences 21, no. 4: 1410. https://doi.org/10.3390/ijms21041410
APA StyleSarasamma, S., Audira, G., Siregar, P., Malhotra, N., Lai, Y. -H., Liang, S. -T., Chen, J. -R., Chen, K. H. -C., & Hsiao, C. -D. (2020). Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. International Journal of Molecular Sciences, 21(4), 1410. https://doi.org/10.3390/ijms21041410