TLR3 Expression Induces Apoptosis in Human Non-Small-Cell Lung Cancer

 ,
,  , ,
, , 
 and
and 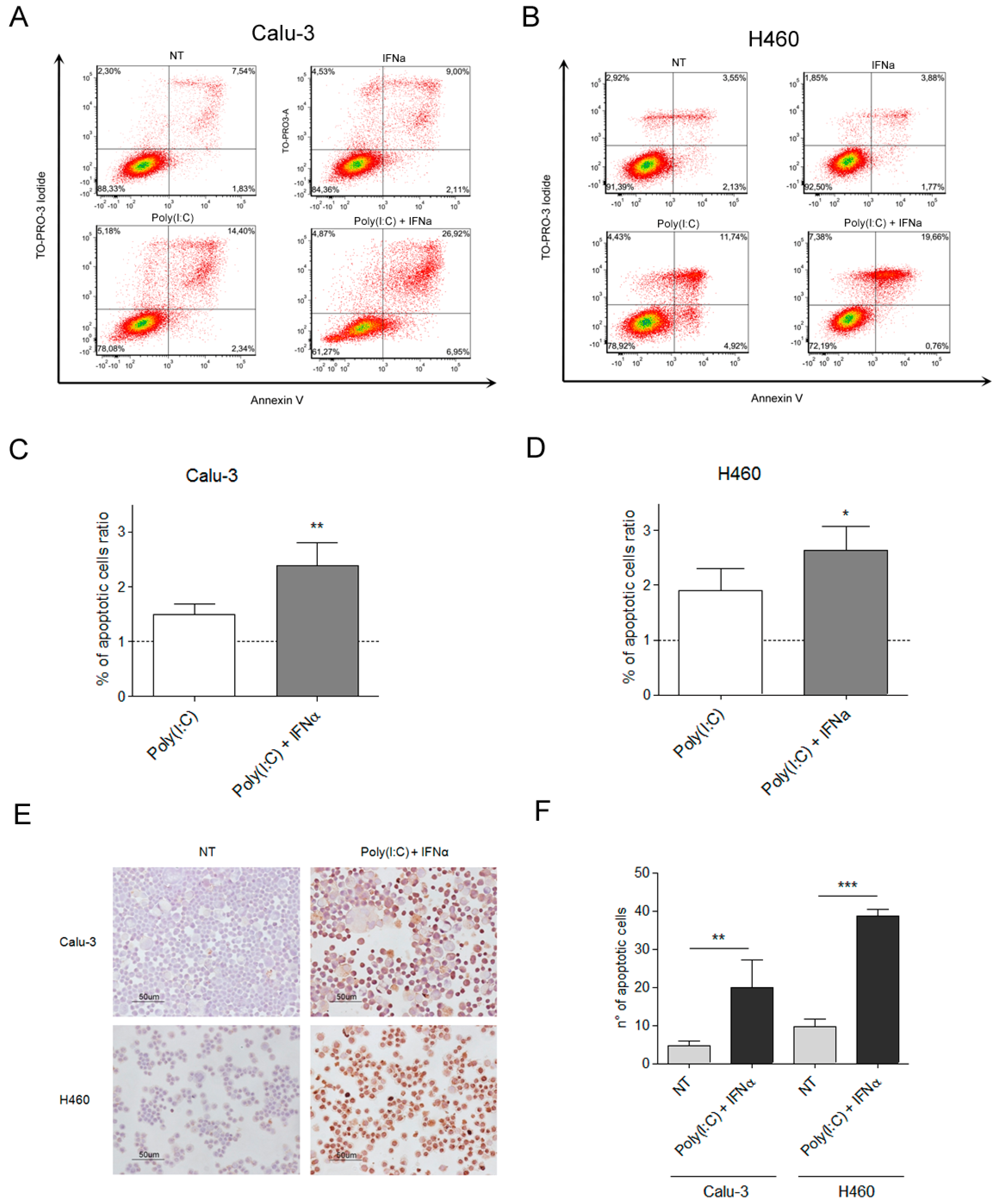{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. TLR3 Induces Apoptosis in Lung Tumor Cells
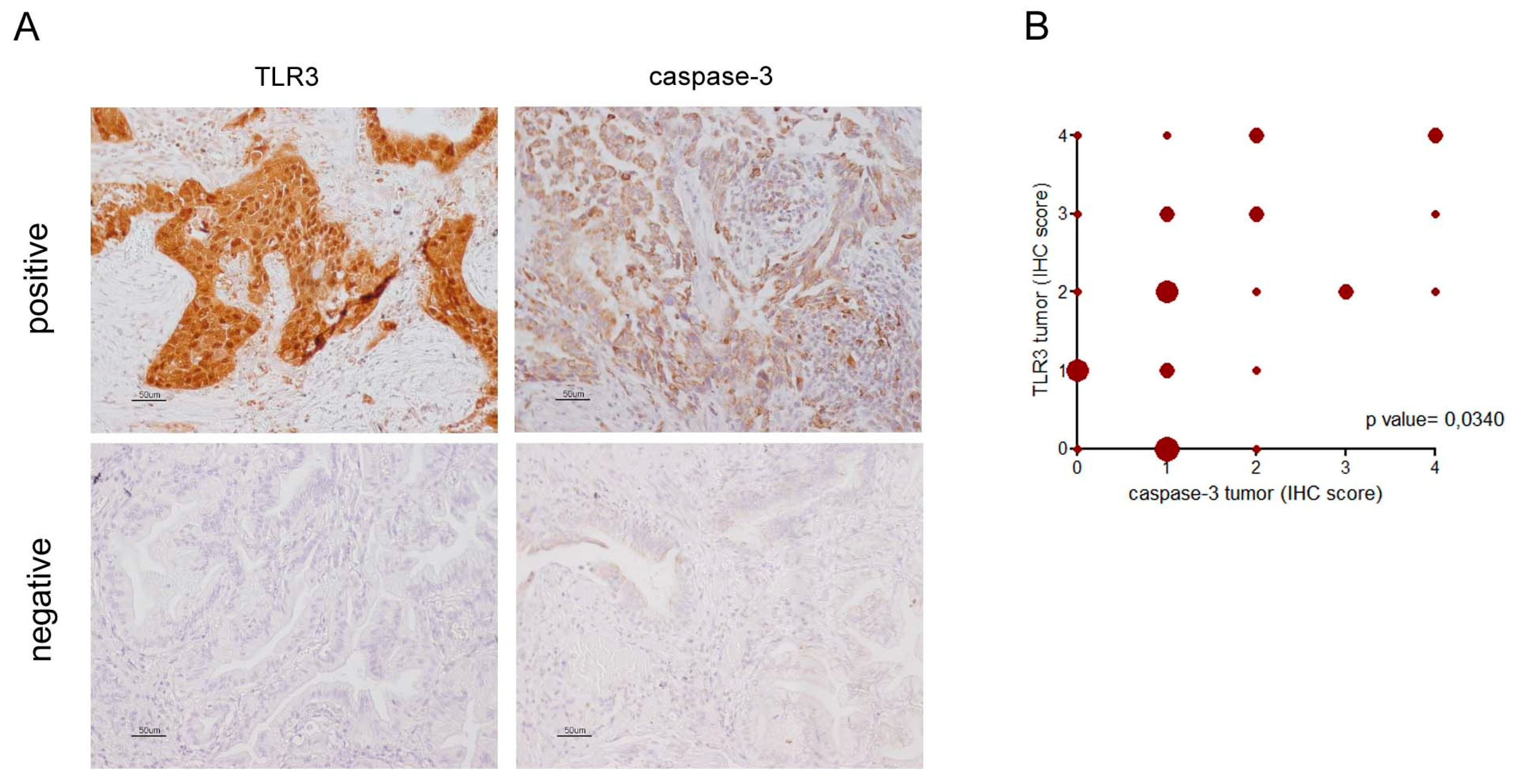
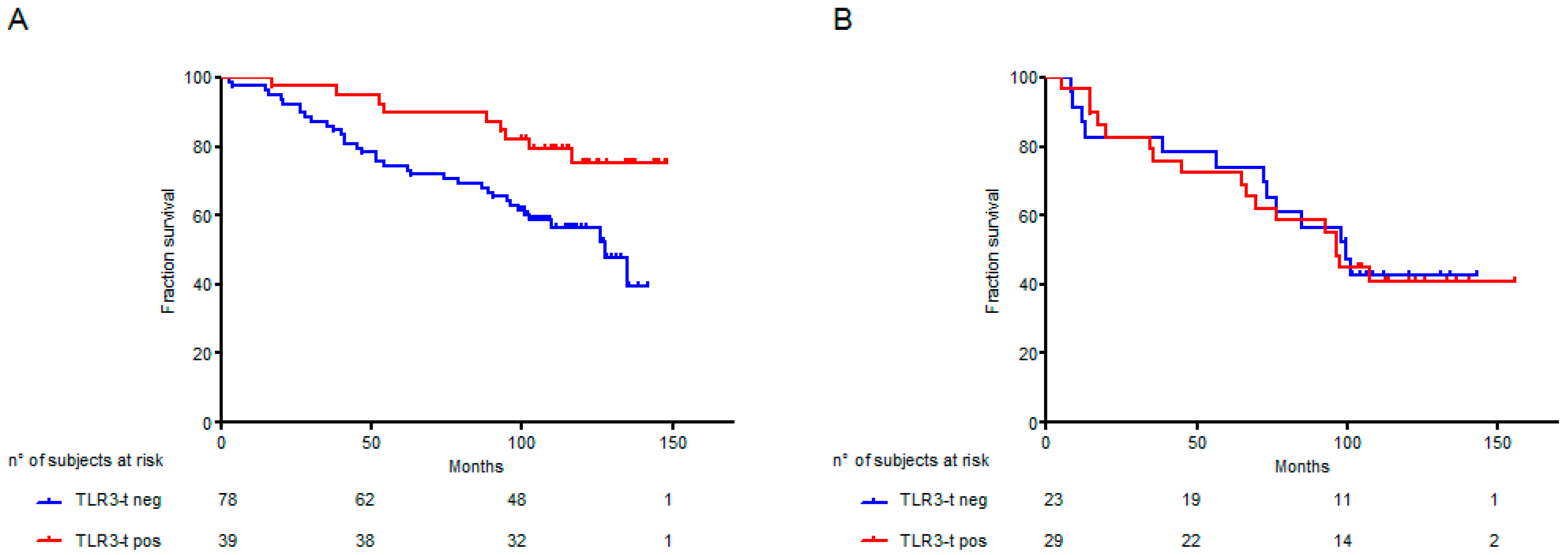
2.2. TLR3 Expression on Cancer Cells Associated with Higher Apoptosis in NSCLC Adenocarcinoma
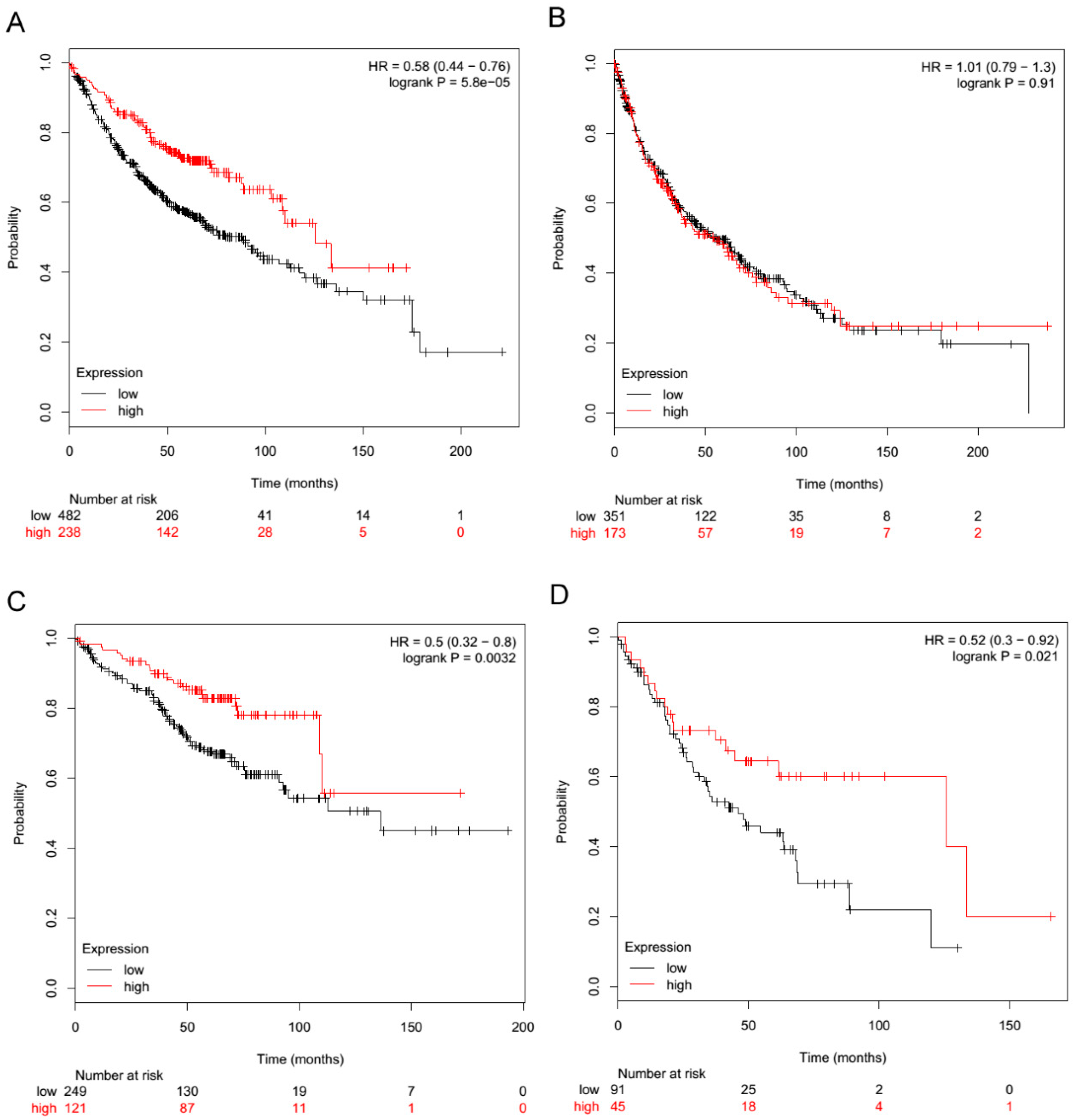
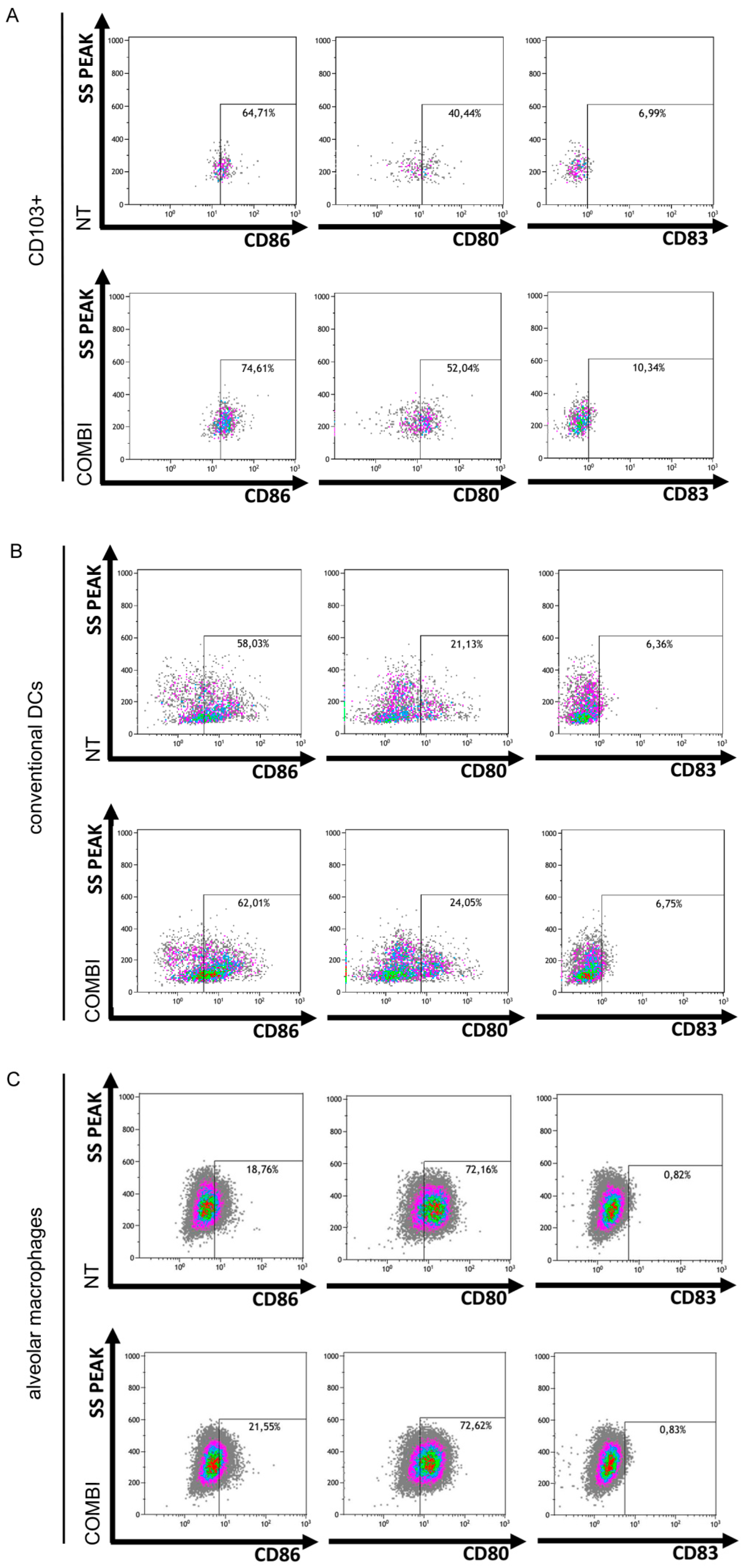
2.3. TLR3 Boosts the Innate Immune Response
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Treatments
4.2. Quantitative PCR Analysis
4.3. Cytofluorimetric Analysis of Lung Cancer Cell Apoptosis
4.4. TUNEL Assay
4.5. Cytofluorometric Analysis of Caspase-3 Activation
4.6. Patients
4.7. Immunohistochemical Analysis of FFPE NSCLC Specimens
4.8. TLR3 Expression in NSCLC by Gene Expression Microarray
4.9. Immunofluorescence Analysis of Mouse Lung Immune Infiltrates
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tatematsu, M.; Seya, T.; Matsumoto, M. Beyond dsRNA: Toll-like receptor 3 signalling in RNA-induced immune responses. Biochem. J. 2014, 458, 195–201. [Google Scholar] [CrossRef]
- Topfer, K.; Kempe, S.; Muller, N.; Schmitz, M.; Bachmann, M.; Cartellieri, M.; Schackert, G.; Temme, A. Tumor evasion from T cell surveillance. J. Biomed. Biotechnol. 2011, 2011, 918471. [Google Scholar] [CrossRef] [Green Version]
- Iribarren, K.; Bloy, N.; Buque, A.; Cremer, I.; Eggermont, A.; Fridman, W.H.; Fucikova, J.; Galon, J.; Spisek, R.; Zitvogel, L.; et al. Trial Watch: Immunostimulation with Toll-like receptor agonists in cancer therapy. OncoImmunology 2015, 5, e1088631. [Google Scholar] [CrossRef] [Green Version]
- Glavan, T.M.; Pavelic, J. The exploitation of Toll-like receptor 3 signaling in cancer therapy. Curr. Pharm. Des. 2014, 20, 6555–6564. [Google Scholar] [CrossRef]
- Kang, J.; Demaria, S.; Formenti, S. Current clinical trials testing the combination of immunotherapy with radiotherapy. J. Immunother. Cancer 2016, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Di, S.; Zhou, M.; Pan, Z.; Sun, R.; Chen, M.; Jiang, H.; Shi, B.; Luo, H.; Li, Z. Combined Adjuvant of Poly I:C Improves Antitumor Effects of CAR-T Cells. Front. Oncol. 2019, 9, 241. [Google Scholar] [CrossRef]
- Takeda, Y.; Kataoka, K.; Yamagishi, J.; Ogawa, S.; Seya, T.; Matsumoto, M. A TLR3-Specific Adjuvant Relieves Innate Resistance to PD-L1 Blockade without Cytokine Toxicity in Tumor Vaccine Immunotherapy. Cell Rep. 2017, 19, 1874–1887. [Google Scholar] [CrossRef] [Green Version]
- Seya, T.; Akazawa, T.; Uehori, J.; Matsumoto, M.; Azuma, I.; Toyoshima, K. Role of toll-like receptors and their adaptors in adjuvant immunotherapy for cancer. Anticancer. Res. 2003, 23, 4369–4376. [Google Scholar] [PubMed]
- Estornes, Y.; Micheau, O.; Renno, T.; Lebecque, S. Dual role of TLR3 in inflammation and cancer cell apoptosis. In Oncogene and Cancer—From Bench to Clinic; InTech: Rijeka, Croatia, 2013; pp. 247–270. [Google Scholar]
- Estornes, Y.; Toscano, F.; Virard, F.; Jacquemin, G.; Pierrot, A.; Vanbervliet, B.; Bonnin, M.; Lalaoui, N.; Mercier-Gouy, P.; Pacheco, Y.; et al. dsRNA induces apoptosis through an atypical death complex associating TLR3 to caspase-8. Cell Death Differ. 2012, 19, 1482–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Pretto, S.; Tagliabue, E.; Balsari, A.; Sfondrini, L. Exploiting poly(I:C) to induce cancer cell apoptosis. Cancer Biol. Ther. 2017, 18, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Motoyama, S.; Wakita, A.; Kawakita, Y.; Liu, J.; Nagaki, Y.; Nanjo, H.; Terata, K.; Imai, K.; Saito, H.; et al. TLR3 expression status predicts prognosis in patients with advanced thoracic esophageal squamous cell carcinoma after esophagectomy. Am. J. Surg. 2018, 216, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Milione, M.; Casalini, P.; Centonze, G.; Le Noci, V.M.; Storti, C.; Alexiadis, S.; Truini, M.; Sozzi, G.; Pastorino, U.; et al. Toll-like receptor 3 as a new marker to detect high risk early stage Non-Small-Cell Lung Cancer patients. Sci. Rep. 2019, 9, 14288–50756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Buque, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunogenic cell death in cancer and infectious disease. Nat. Rev. Immunol. 2017, 17, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Salaun, B.; Coste, I.; Rissoan, M.C.; Lebecque, S.J.; Renno, T. TLR3 can directly trigger apoptosis in human cancer cells. J. Immunol. 2006, 176, 4894–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salaun, B.; Lebecque, S.; Matikainen, S.; Rimoldi, D.; Romero, P. Toll-like receptor 3 expressed by melanoma cells as a target for therapy? Clin. Cancer Res. 2007, 13, 4565–4574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.; Zhang, Y.; Lv, Q.; Liu, B.; Jin, M.; Zhang, W.; He, Q.; Deng, M.; Liu, X.; Li, G.; et al. Toll-like receptor 3 (TLR3) induces apoptosis via death receptors and mitochondria by up-regulating the transactivating p63 isoform alpha (TAP63alpha). J. Biol. Chem. 2011, 286, 15918–15928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef]
- Gyorffy, B.; Surowiak, P.; Budczies, J.; Lanczky, A. Online survival analysis software to assess the prognostic value of biomarkers using transcriptomic data in non-small-cell lung cancer. PLoS ONE 2013, 8, e82241. [Google Scholar] [CrossRef] [Green Version]
- Desch, A.N.; Randolph, G.J.; Murphy, K.; Gautier, E.L.; Kedl, R.M.; Lahoud, M.H.; Caminschi, I.; Shortman, K.; Henson, P.M.; Jakubzick, C.V. CD103+ pulmonary dendritic cells preferentially acquire and present apoptotic cell-associated antigen. J. Exp. Med. 2011, 208, 1789–1797. [Google Scholar] [CrossRef] [Green Version]
- Alkurdi, L.; Virard, F.; Vanbervliet, B.; Weber, K.; Toscano, F.; Bonnin, M.; Le, S.N.; Lantuejoul, S.; Micheau, O.; Renno, T.; et al. Release of c-FLIP brake selectively sensitizes human cancer cells to TLR3-mediated apoptosis. Cell Death Dis. 2018, 9, 874. [Google Scholar] [CrossRef]
- Blachere, N.E.; Darnell, R.B.; Albert, M.L. Apoptotic cells deliver processed antigen to dendritic cells for cross-presentation. PLoS Biol. 2005, 3, e185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, G.; Rega, A.; Morello, S.; Luciano, A.; Arra, C.; Pinto, A.; Sorrentino, R. Polyinosinic-polycytidylic acid limits tumor outgrowth in a mouse model of metastatic lung cancer. J. Immunol. 2012, 188, 5357–5364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebihara, T.; Azuma, M.; Oshiumi, H.; Kasamatsu, J.; Iwabuchi, K.; Matsumoto, K.; Saito, H.; Taniguchi, T.; Matsumoto, M.; Seya, T. Identification of a polyI:C-inducible membrane protein that participates in dendritic cell-mediated natural killer cell activation. J. Exp. Med. 2010, 207, 2675–2687. [Google Scholar] [CrossRef] [PubMed]
- Shime, H.; Kojima, A.; Maruyama, A.; Saito, Y.; Oshiumi, H.; Matsumoto, M.; Seya, T. Myeloid-derived suppressor cells confer tumor-suppressive functions on natural killer cells via polyinosinic:polycytidylic acid treatment in mouse tumor models. J. Innate Immun. 2014, 6, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Shime, H.; Matsumoto, M.; Oshiumi, H.; Tanaka, S.; Nakane, A.; Iwakura, Y.; Tahara, H.; Inoue, N.; Seya, T. Toll-like receptor 3 signaling converts tumor-supporting myeloid cells to tumoricidal effectors. Proc. Natl. Acad. Sci. USA 2012, 109, 2066–2071. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Zhao, Z.; Jiang, S.; Lin, Y.; Yang, H.; Xie, Z.; Lin, Y.; Long, H. Immunological markers predict the prognosis of patients with squamous non-small cell lung cancer. Immunol. Res. 2015, 62, 316–324. [Google Scholar] [CrossRef]
- De Cesare, M.; Sfondrini, L.; Campiglio, M.; Sommariva, M.; Bianchi, F.; Perego, P.; van Rooijen, N.; Supino, R.; Rumio, C.; Zunino, F.; et al. Ascites regression and survival increase in mice bearing advanced-stage human ovarian carcinomas and repeatedly treated intraperitoneally with CpG-ODN. J. Immunother. 2010, 33, 8–15. [Google Scholar] [CrossRef]
- Le Noci, V.; Tortoreto, M.; Gulino, A.; Storti, C.; Bianchi, F.; Zaffaroni, N.; Tripodo, C.; Tagliabue, E.; Balsari, A.; Sfondrini, L. Poly(I:C) and CpG-ODN combined aerosolization to treat lung metastases and counter the immunosuppressive microenvironment. OncoImmunology 2015, 4, e1040214. [Google Scholar] [CrossRef] [Green Version]
- Sommariva, M.; Le Noci, V.; Storti, C.; Bianchi, F.; Tagliabue, E.; Balsari, A.; Sfondrini, L. Activation of NK cell cytotoxicity by aerosolized CpG-ODN/poly(I:C) against lung melanoma metastases is mediated by alveolar macrophages. Cell. Immunol. 2017, 313, 52–58. [Google Scholar] [CrossRef]
- Le Noci, V.; Guglielmetti, S.; Arioli, S.; Camisaschi, C.; Bianchi, F.; Sommariva, M.; Storti, C.; Triulzi, T.; Castelli, C.; Balsari, A.; et al. Modulation of pulmonary microbiota by antibiotic or probiotic aerosol therapy: A strategy to promote immunosurveillance against lung metastases. Cell Rep. 2018, 24, 3528–3538. [Google Scholar] [CrossRef] [Green Version]
- Salaun, B.; Zitvogel, L.; Asselin-Paturel, C.; Morel, Y.; Chemin, K.; Dubois, C.; Massacrier, C.; Conforti, R.; Chenard, M.P.; Sabourin, J.C.; et al. TLR3 as a biomarker for the therapeutic efficacy of double-stranded RNA in breast cancer. Cancer Res. 2011, 71, 1607–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turdo, F.; Bianchi, F.; Gasparini, P.; Sandri, M.; Sasso, M.; De Cecco, L.; Forte, L.; Casalini, P.; Aiello, P.; Sfondrini, L.; et al. CDCP1 is a novel marker of the most aggressiveness human triple-negative breast cancer. Oncotarget 2016, 7, 69649–69665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Sasso, M.; Turdo, F.; Beretta, G.L.; Casalini, P.; Ghirelli, C.; Sfondrini, L.; Menard, S.; Tagliabue, E.; Campiglio, M. Fhit nuclear import following EGF stimulation sustains proliferation of Breast Cancer cells. J. Cell. Physiol. 2015, 230, 2661–2670. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Sommariva, M.; De Cecco, L.; Triulzi, T.; Romero-Cordoba, S.; Tagliabue, E.; Sfondrini, L.; Balsari, A. Expression and prognostic significance of the autoimmune regulator gene in breast cancer cells. Cell Cycle 2016, 15, 3220–3229. [Google Scholar] [CrossRef]
- Cox, D.R.; Oakes, D. Analysis of Survival Data; Chapman and Hall: London, UK, 1984. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, F.; Alexiadis, S.; Camisaschi, C.; Truini, M.; Centonze, G.; Milione, M.; Balsari, A.; Tagliabue, E.; Sfondrini, L. TLR3 Expression Induces Apoptosis in Human Non-Small-Cell Lung Cancer. Int. J. Mol. Sci. 2020, 21, 1440. https://doi.org/10.3390/ijms21041440
Bianchi F, Alexiadis S, Camisaschi C, Truini M, Centonze G, Milione M, Balsari A, Tagliabue E, Sfondrini L. TLR3 Expression Induces Apoptosis in Human Non-Small-Cell Lung Cancer. International Journal of Molecular Sciences. 2020; 21(4):1440. https://doi.org/10.3390/ijms21041440
Chicago/Turabian StyleBianchi, Francesca, Spyridon Alexiadis, Chiara Camisaschi, Mauro Truini, Giovanni Centonze, Massimo Milione, Andrea Balsari, Elda Tagliabue, and Lucia Sfondrini. 2020. "TLR3 Expression Induces Apoptosis in Human Non-Small-Cell Lung Cancer" International Journal of Molecular Sciences 21, no. 4: 1440. https://doi.org/10.3390/ijms21041440
APA StyleBianchi, F., Alexiadis, S., Camisaschi, C., Truini, M., Centonze, G., Milione, M., Balsari, A., Tagliabue, E., & Sfondrini, L. (2020). TLR3 Expression Induces Apoptosis in Human Non-Small-Cell Lung Cancer. International Journal of Molecular Sciences, 21(4), 1440. https://doi.org/10.3390/ijms21041440